1. Introduction
Overweight and obesity have achieved epidemic proportions worldwide, mostly due to lifestyle choices, such as low levels of physical activity and the adoption of a high-fat diet (HFD) [
1,
2]. These numbers raise concerns about the consequences of sexual health in men of reproductive age suffering from excess adiposity and common comorbidities, such as type 2 diabetes (T2D) [
3,
4]. These concerns were aggravated by recent evidence of the intergenerational effects of acquired traits, i.e., by the evidence that acquired traits can compromise the health of offspring, especially in male reproductive health [
5,
6].
We have previously described the effects of an HFD, even if temporary, in sperm parameters and testicular composition of mice [
7,
8]. Hereby we investigate the effects of an HFD on sperm parameters and testicular physiology of the offspring (sons), with a focus on testicular antioxidative status.
2. Experiments
2.1. Animal Model
This study was performed in 2 generations of Mus musculus C57BL6/J mice. The first generation (Generation F0) was originated from normoponderal progenitors (both male and female) fed standard chow (#F4031, BioServ, Flemington, NJ, USA—carbohydrate: 61.6%, protein: 20.5%, and fat: 7.2% (16.3% Kcals)) and water ad libitum. After weaning (21–23 days), F0 mice (n = 36) were randomly divided in three groups: control (CTRL) (n = 12), HFD (n = 12). and HFDt (n = 12). The mice from the CTRL group were fed with a standard chow. Mice from the HFD group received a fat-enriched diet (#F3282, BioServ, Flemington, NJ, USA—carbohydrate: 35.7%, protein: 20.5%, and fat: 36.0% (59.0% Kcals)). The mice from the HFDt group were fed a fat-enriched diet for 60 days (#F3282, BioServ, New Jersey, USA) and then switched to standard chow (#F4031, BioServ, NJ, USA). F0 mice were mated at 120 days of age with normoponderal, chow-fed, same-age randomly selected females to generate the F1 generation. Mating lasted for 8 days and consisted of placing a male and a female in the same cage for 6 h each day without water or food supply. After weaning, F1 mice were assigned to the same experimental group as their fathers: CTRL—sons of CTRL (n = 12); HFD—sons of HFD (n = 12); HFDt—sons of HFDt (n = 12). Litters were generated until the target number of mice per group (n = 12) was achieved. In this generation (F1), all mice were fed with standard chow. Food and water were supplied without restrictions. The mating of F1 mice was performed under the same conditions as those used for their progenitors (Generation F0). Mice from both generations were killed by cervical dislocation 200 days after weaning, and tissues were collected for further analysis. Total body weight, water, and food intake were monitored weekly from weaning to sacrifice. The animal model is compliant with the ARRIVE guidelines and was licensed by the Portuguese Veterinarian and Food Department (0421/000/000/2016).
2.2. Glucose Homeostasis Assessment
One week before sacrifice, glucose homeostasis was evaluated using the intraperitoneal glucose tolerance test (ipGTT) and intraperitoneal insulin resistance test (ipITT) according to the previously described protocol [
8]. Fasting glucose was measured before sacrifice, as previously described [
7,
8]. Serum was separated from blood obtained by cardiac puncture at sacrifice, and insulin was measured via ELISA [
7]. HOMA2 indexes were calculated based on the serum insulin and glucose at sacrifice, using the HOMA2 Calculator [
9].
2.3. Enzymatic Activity of Antioxidant Enzymes and Mitochondrial Complexes
The enzymatic activity of antioxidant enzymes and mitochondrial complexes was evaluated from testicular extracts. A phase separation protocol was used to obtain a mitochondria-rich fraction and a cytosolic, mitochondria-free fraction according to a protocol previously described [
7]. Lipid peroxidation was measured in the cytosolic fraction of the testicular extract by the TBARS assay [
7]. Enzymatic activities of glutathione peroxidase (GPx), glutathione S-reductase (GSR), and mitochondrial complex I were measured using fluoroscopic methods in 96-well plates, as previously described [
7]. Enzymatic activities of superoxide dismutase (SOD), citrate synthase, mitochondrial complex II and mitochondrial complex IV were measured by colorimetric methods in 96-well plates, as previously described [
7]. All colorimetric and fluorometric readings were obtained using a Biotek Synergy H1 plate reader (Winooski, VT, USA). Catalase (CAT) activity was polarographically determined following oxygen production resulting from H
2O
2 decomposition using a Clark-type oxygen electrode (Hansatech, Norfolk, UK), as previously described [
7].
2.4. Assessment of Sperm Parameters
Sperm was collected from the right epididymis of each mice after sacrifice. Sperm count and motility were immediately assessed as previously described [
7,
8]. Sperm viability and morphology were evaluated using specific staining techniques and optical microscopy as previously described [
7,
8].
2.5. Untargeted Metabolomics and Lipidomics
A combined extraction of polar and nonpolar metabolites from testicular tissue (50 mg) was performed, as previously described [
7,
8]. Two fractions resulted from the method—an aqueous, polar fraction, and an organic, nonpolar fraction.
1H-NMR was performed to analyze and quantify the metabolites in the polar fraction according to our methods [
7,
8]. GC-MS was used to analyze and quantify the nonpolar metabolites soluble in the organic fraction [
7].
2.6. Statistics
Univariate parametric statistics were the preferred statistical methods. The assumptions of normality and homoscedasticity requested for parametric statistics were tested for each variable using the Kolmogorov–Smirnoff test with Lillefor’s correction and Levene’s test, respectively. Univariate ANOVA was corrected for pairwise corrections by Tukey’s honest significant difference (HSD). ipGTT and ipITT data were tested using repeated-measures (RM) ANOVA corrected by Šidak’s method for pairwise comparisons. Data were previously tested for sphericity using Bartlett’s test. The distribution of sperm defects was tested using the χ2 test, and column proportions were tested by the Z-test corrected for pairwise comparisons using Bonferroni’s method. The significance cutoff was set when p < 0.05. All methods were performed using IBM SPSS Statistics v26 (Armonk, NY, USA).
3. Results
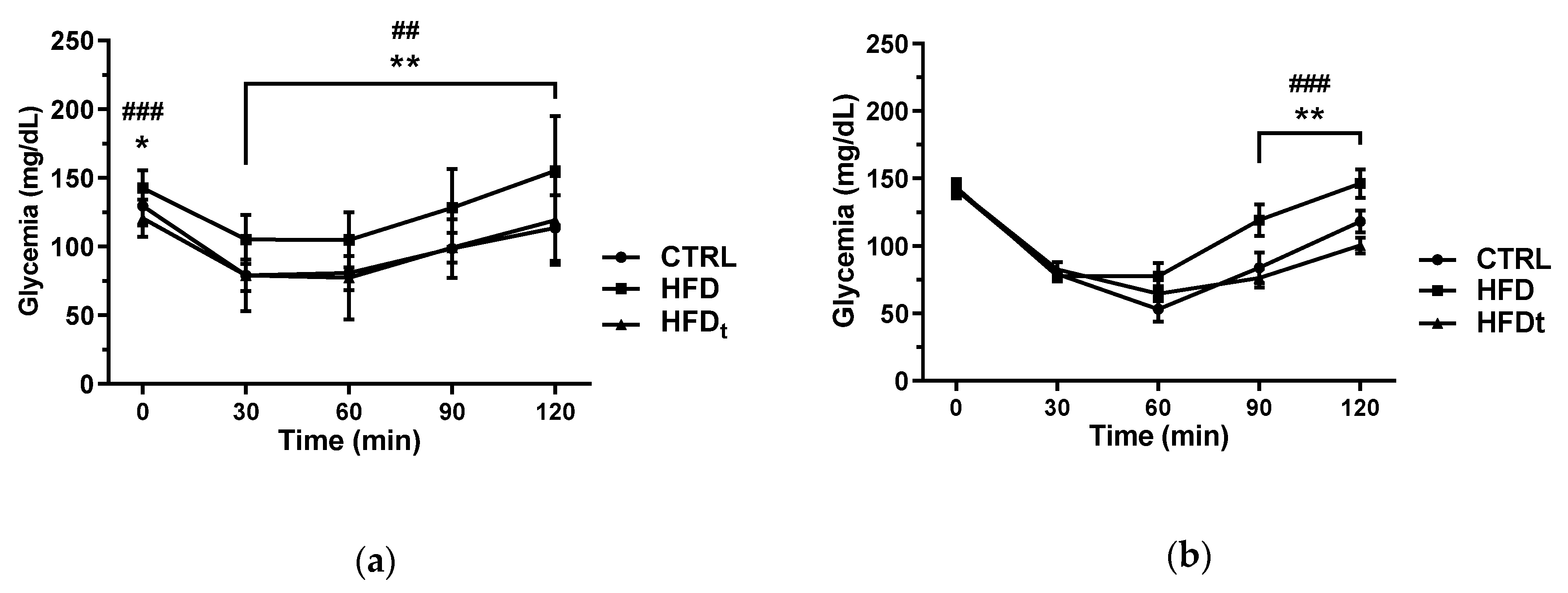
3.1. The Offspring of HFD-Fed Mice Display Abnormal Insulin Tolerance
Glucose homeostasis was assessed by ipGTT, ipITT, and HOMA2 indexes. The sons of the mice fed a lifelong HFD displayed higher serum glucose at 90 and 120 min of the ipITT than that of the sons of CTRL and HFDt (
Figure 1). Regarding HOMA2 indexes, no differences were found between the sons of diet-challenged mice.
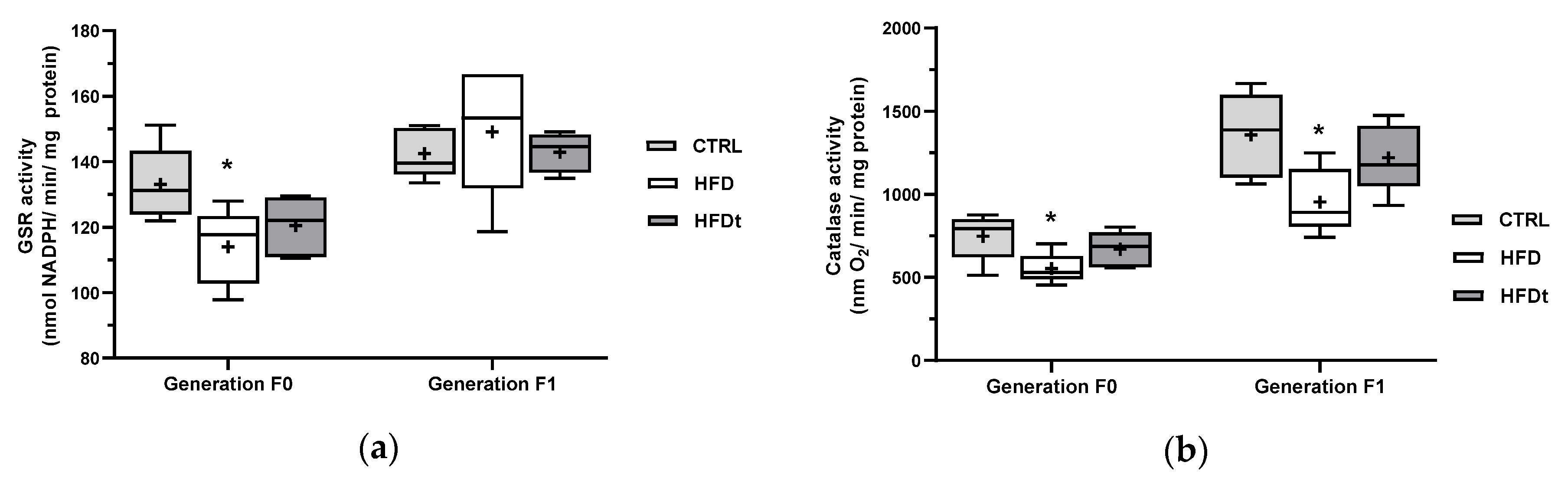
3.2. The Adoption of HFD Inhibits Testicular Antioxidant Defences Even in Offspring
The enzymatic activity of antioxidant enzymes was measured in testes. Mice fed an HFD for a lifetime had decreased activity of GSR and catalase (
Figure 2). Interestingly, this phenotype was partially inherited by their sons, which had decreased testicular catalase activity, when compared to the sons of CTRL.
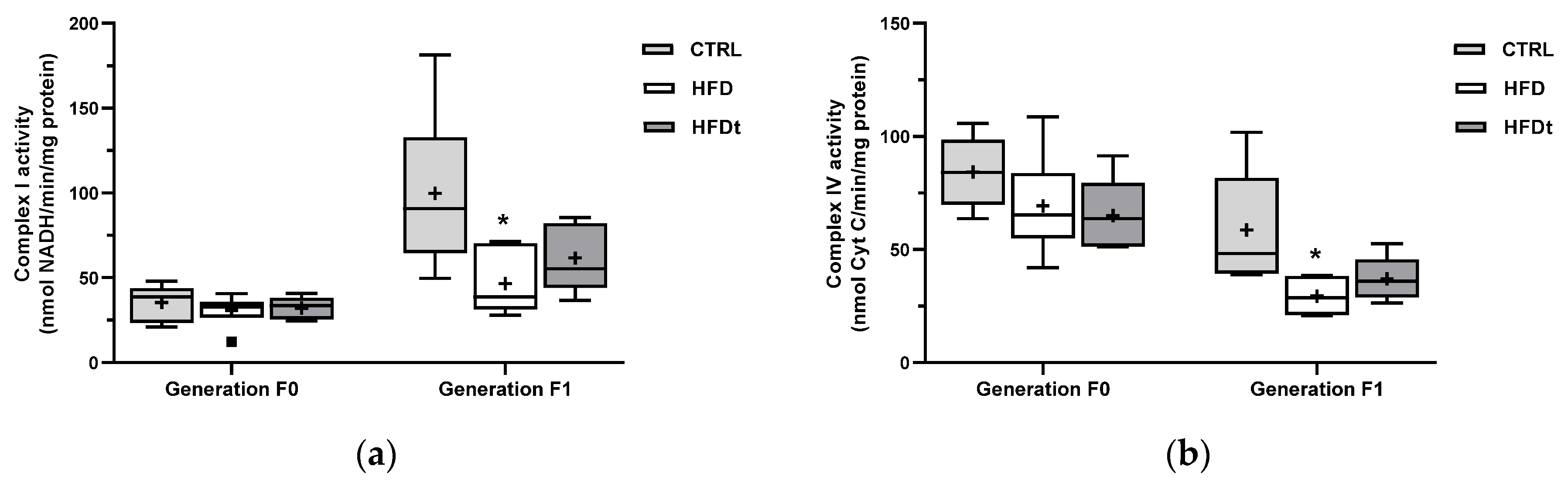
3.3. Testicular Mitochondrial Defects Are Only Detected in Offspring of HFD-Fed Mice
The enzymatic activity of mitochondrial complexes was measured in testes. No changes were found between groups of diet-challenged mice (generation F0) (
Figure 3). The sons of lifelong HFD-fed mice showed reduced activity of mitochondrial complex I and complex IV.
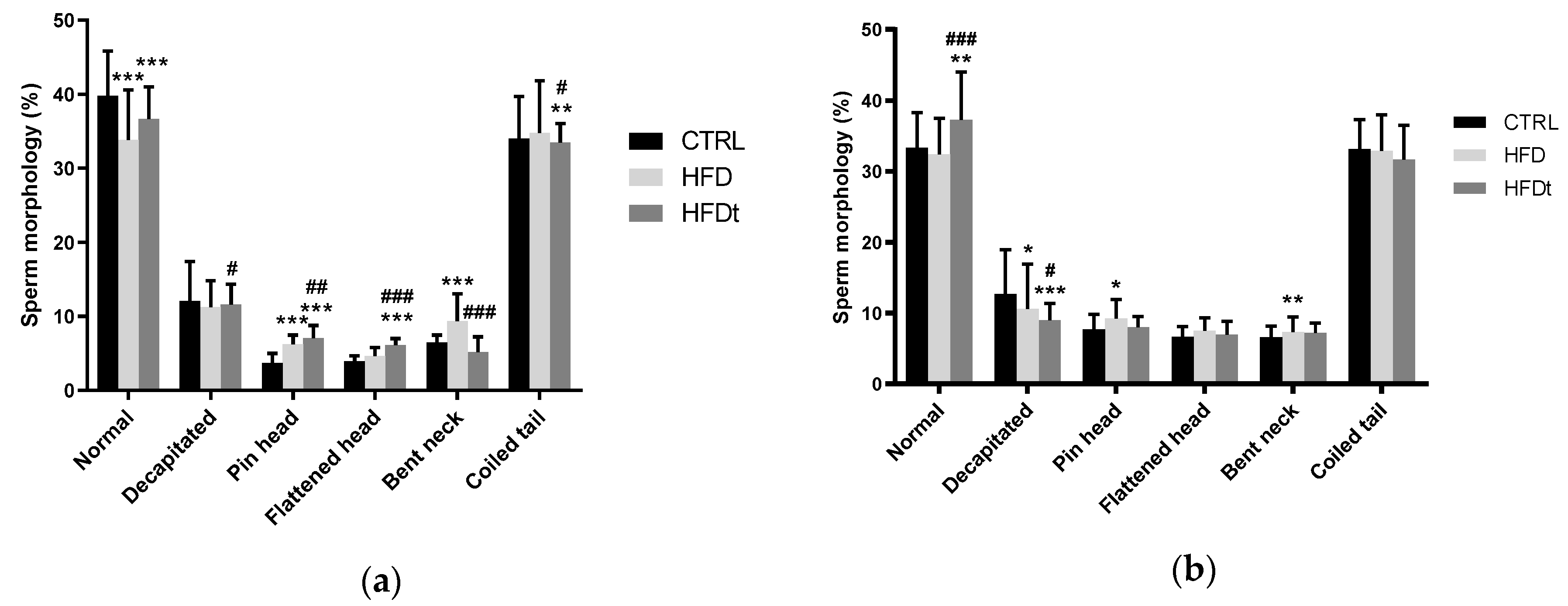
3.4. Paternal HFD Causes Intergenerational Sperm Defects
Epidydimal sperm parameters were evaluated after sacrifice. Sperm count and motility were assessed immediately after collection. Contrary to their progenitors, the sons of HFD and HFDt did not show differences in sperm motility and viability when compared to sons of CTRL. However, regarding sperm morphology, the sons of HFD mice had a greater prevalence of pinhead defects than that of the sons of CTRL and a greater prevalence of bent neck defects than that of the sons of HFDt (
Figure 4).
3.5. Testicular Metabolic and Lipidomic Signatures of HFD Are Not Inherited by Direct Offspring
Testicular polar and nonpolar metabolites were extracted and quantified by either 1H-NMR or GC-MS. However, no significant changes were found in the testicular content of the sons of the diet-challenged mice.
4. Discussion
In this work, we evaluated the impact of an ancestral paternal exposure to an HFD, either lifelong or up to early adulthood, on testicular metabolism and sperm parameters of the direct offspring (sons). To do so, we evaluated the oxidative status, the mitochondrial function, and the metabolite content in the testis of mice whose parents were fed standard chow (CTRL), a lifelong HFD (HFD), or an HFD from weaning to early adulthood and then replaced with standard chow (HFDt).
We also evaluated the whole-body glucose homeostasis to search for cues of metabolic syndrome that may be inherited by the offspring. Indeed, the sons of HFD mice showed signs of insulin intolerance regarding their performance in the ipITT (
Figure 1). We previously described that the inhibition of testicular antioxidant enzyme catalase and GSR and the increased testicular content of ω6-polyunsaturated fatty acids elicit that a lifelong HFD promotes a pro-inflammatory environment in testis [
7]. To balance this HFD-induced inflammation, mice fed with HFD have increased testicular glutathione (GSH) and taurine levels [
8]. Similarly to their progenitors, the sons of HFD mice showed lower enzymatic activity of catalase in testes, suggesting an inheritable trait caused by the pro-inflammatory testicular environment of the progenitor. However, no differences in lipid peroxidation in both diet-challenged mice and their sons were found. Notwithstanding, no changes in testicular metabolome were found in the sons of HFD when compared to the sons of CTRL and HFDt. Therefore, it is unclear whether the testicular oxidative balance is being balanced by other mechanisms. This is even more interesting when considering the decrease in the mitochondrial activity of complex I and IV found in the sons of HFD. In contrast, the sons of HFD mice have the highest prevalence of sperm head defects (compared to sons of CTRL) and sperm neck defects (compared to sons of HFDt). Indeed, oxidative damage in the testis has been linked to a higher prevalence of abnormal sperm [
10]. Hence, the changes in antioxidant defenses and mitochondrial activity observed in the testis of the sons of HFD mice might be associated with the increase in abnormal sperm.
Interestingly, the differences found in sons of diet-challenged mice in all evaluated parameters are restricted to the sons of HFD. These mice were the only group receiving an HFD at the moment of conception. Several reports mention that sperm carries non-genomic factors, such as small non-coding RNA (sncRNA) and epigenetic modifications, that are sensitive to diet [
11,
12]. Therefore, the phenotypes observed in the sons of HFD mice are likely manifestations of intergenerational effects of an HFD; i.e., they are the result of direct exposure of the male gamete to the toxicant (HFD) [
13].
5. Conclusions
The adoption of HFD by fathers causes intergenerational signatures in testis, notably in antioxidant defenses and mitochondrial activity. Those signatures, in turn, are associated with a higher prevalence of sperm head and neck defects. The negative impact of paternal HFD is more evident if it is continued at the moment of conception.
Author Contributions
L.C., P.F.O., M.G.A., and R.L.B. contributed to study design, analysis, and interpretation of data. L.R., I.J., K.S., T.M., R.A.V., and L.C. performed experimental work. L.C. edited the images and tables, performed the statistics, and contributed to the analysis and interpretation of data. J.F.R., and R.A.V. critically reviewed the manuscript and suggested modifications. All of the authors contributed to manuscript writing/editing and approved the final version.
Institutional Review Board Statement
The animal model is compliant with the ARRIVE guidelines and was licensed by the Portuguese Veterinarian and Food Department (0421/000/000/2016).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data included in this report is available upon reasonable request.
Acknowledgments
This work was supported by the Portuguese Foundation for Science and Technology: L. Crisóstomo (SFRH/BD/128584/2017), M.G. Alves (IFCT2015 and PTDC/MEC-AND/28691/2017), P.F. Oliveira (IFCT2015), UMIB (UID/Multi/00215/2019), and QOPNA (UID/QUI/00062/2019) co-funded by FEDER funds (POCI/COMPETE 2020); by the Portuguese Society of Diabetology: L. Crisóstomo and M.G. Alves (“Nuno Castel-Branco” research grant and Group of Fundamental and Translational Research); and by the Croatian Science Foundation: K. Starčević (IP-2016-06-3163). NMR data were collected at the UC-NMR facility, which is supported in part by FEDER—European Regional Development Fund through the COMPETE Programme (Operational Programme for Competitiveness) and by National Funds through FCT—Fundação para a Ciência e a Tecnologia (Portuguese Foundation for Science and Technology) through grants REEQ/481/QUI/2006, RECI/QEQ-QFI/0168/2012, CENTRO-07-CT62-FEDER-002012, and Rede Nacional de Ressonância Magnética Nuclear (RNRMN).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 1H-NMR | proton nuclear magnetic resonance |
| ANOVA | analysis of variance |
| CTRL | standard diet (standard chow) |
| GC-MS | gaseous chromatography–mass spectroscopy |
| GPx | glutathione peroxidase |
| GSH | glutathione |
| GSR | glutathione S-reductase |
| HFD | high-fat diet |
| HFDt | transient high-fat diet |
| HSD | honest significant difference |
| RM | repeated measures |
| SD | standard deviation |
| sncRNA | small non-coding RNA |
| SOD | superoxide dismutase |
References
- World Health Organization. Global Status Report on Noncommunicable Diseases 2014; WHO Press: Geneva, Switzerland, 2014; p. 280. [Google Scholar]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imani, M.; Talebi, A.R.; Fesahat, F.; Rahiminia, T.; Seifati, S.M.; Dehghanpour, F. Sperm parameters, DNA integrity, and protamine expression in patients with type II diabetes mellitus. J. Obstet. Gynaecol. 2021, 41, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Salas-Huetos, A.; Maghsoumi-Norouzabad, L.; James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Becerra-Tomás, N.; Javid, A.Z.; Abed, R.; Torres, P.J.; et al. Male adiposity, sperm parameters and reproductive hormones: An updated systematic review and collaborative meta-analysis. Obes. Rev. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Pavlinkova, G.; Margaryan, H.; Zatecka, E.; Valaskova, E.; Elzeinova, F.; Kubatova, A.; Bohuslavova, R.; Peknicova, J. Transgenerational inheritance of susceptibility to diabetes-induced male subfertility. Sci. Rep. 2017, 7, 4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, J.R.; Jenkins, T.G.; Carrell, D.T.; Hotaling, J.M. Obesity, male infertility, and the sperm epigenome. Fertil. Steril. 2017, 107, 848–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisóstomo, L.; Videira, R.A.; Jarak, I.; Starčević, K.; Mašek, T.; Rato, L.P.; Raposo, J.F.; Batterham, R.L.; Oliveira, P.F.; Alves, M.G. Diet during early life defines testicular lipid content and sperm quality in adulthood. Am. J. Physiol. Metab. 2020, 319, E1061–E1073. [Google Scholar] [CrossRef] [PubMed]
- Crisóstomo, L.; Rato, L.; Jarak, I.; Silva, B.M.; Raposo, J.F.; Batterham, R.L.; Oliveira, P.F.; Alves, M.G. A switch from high-fat to normal diet does not restore sperm quality but prevents metabolic syndrome. Reproduction 2019, 158, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.C.; Matthews, D.R.; Hermans, M.P. Correct Homeostasis Model Assessment (HOMA) Evaluation Uses the Computer Program. Diabetes Care 1998, 21, 2191–2192. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.F.; Tomás, G.D.; Dias, T.R.; Martins, A.D.; Rato, L.; Alves, M.G.; Silva, B.M. White tea consumption restores sperm quality in prediabetic rats preventing testicular oxidative damage. Reprod. Biomed. Online 2015, 31, 544–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nätt, D.; Kugelberg, U.; Casas, E.; Nedstrand, E.; Zalavary, S.; Henriksson, P.; Nijm, C.; Jäderquist, J.; Sandborg, J.; Flinke, E. Human sperm displays rapid responses to diet. PLoS Biol. 2019, 17, e3000559. [Google Scholar] [CrossRef] [Green Version]
- Watkins, A.J.; Dias, I.; Tsuro, H.; Allen, D.; Emes, R.D.; Moreton, J.; Wilson, R.; Ingram, R.J.M.; Sinclair, K.D. Paternal diet programs offspring health through sperm- and seminal plasma-specific pathways in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 10064–10069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, E.; Ben Maamar, M.; Skinner, M.K. Chapter 2—Definition of epigenetic transgenerational inheritance and biological impacts. In Transgenerational Epigenetics, 2nd ed.; Tollefsbol, T.O., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 13, pp. 13–24. [Google Scholar]
| Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}



