
Effects of Some New Antioxidants on Apoptosis and ROS Production in AFB1 Treated Chickens †



 ,
,  and
and
Abstract
:1. Introduction
2. Experiments
2.1. Ethics Statement
2.2. Animals and Diet
2.3. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.3.1. RNA Extraction and Complementary DNA (cDNA) Synthesis
2.3.2. Selection of Gene of Interest and RT-qPCR
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
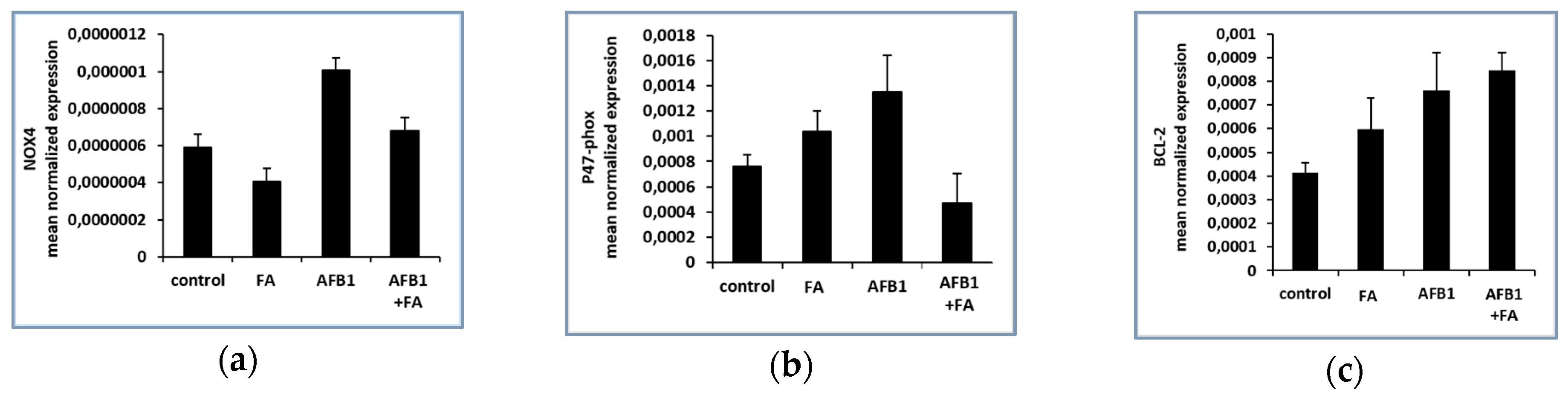
3.1. Gene Expression Results
NOX4, p47-phox and BCL-2 Genes Expression Results
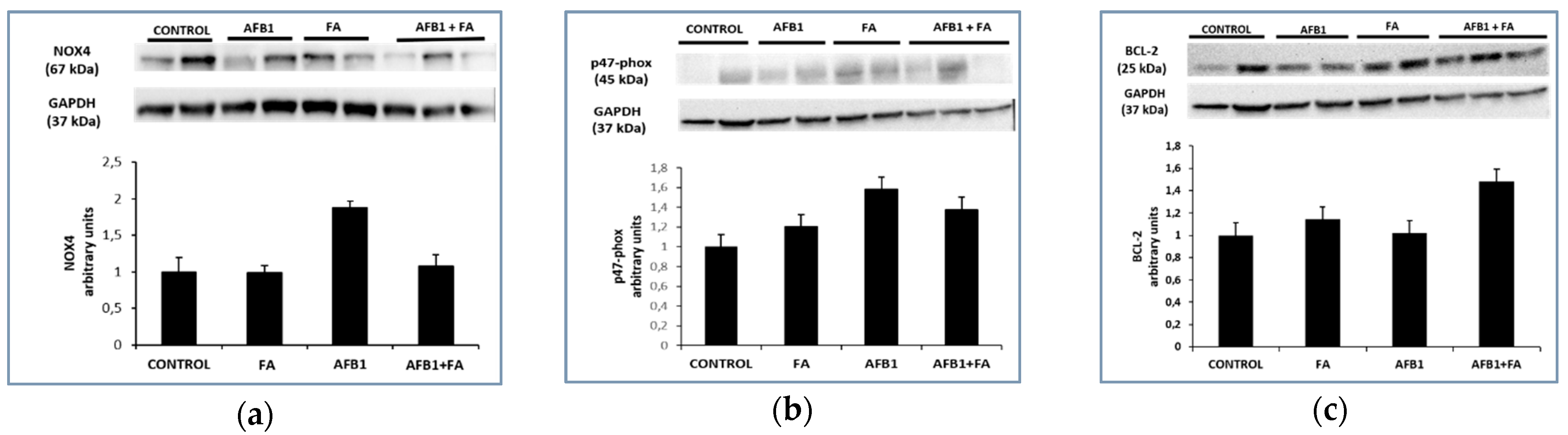
3.2. Protein Expression Results
NOX4, p47-phox and BCL-2 Proteins Expression Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | linear dichroism |
| AFB1 | Aflatoxin B1 |
| ROS | Reactive oxygen species |
| NOX4 | NADPH oxidase 4 |
| FA | Feed Additive |
References
- Omotayo, O.P.; Omotayo, A.O.; Mwanza, M.; Babalola, O.O. Prevalence of Mycotoxins and Their Consequences on Human Health. Toxicol. Res. 2019, 35, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raduly, Z.; Szabo, L.; Madar, A.; Pocsi, I.; Csernoch, L. Toxicological and Medical Aspects of Aspergillus-Derived Mycotoxins Entering the Feed and Food Chain. Front. Microbiol. 2019, 10, 2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marasas, W.F.O.; Gelderblom, W.; Shephard, G.; Vismer, H. Mycotoxins: A global problem. Mycotoxins Detect. Methods Manag. Public Health Agric. Trade 2008, 29–39. [Google Scholar] [CrossRef]
- A review of human carcinogens: Chemical agents and related occupations. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans 2012; IARC Working Group (Ed.) IARC: Lyon, France, 2012; Volume 100F, pp. 225–244. [Google Scholar]
- Rotimi, O.A.; Rotimi, S.O.; Duru, C.U.; Ebebeinwe, O.J.; Abiodun, A.O.; Oyeniyi, B.O.; Faduyile, F.A. Acute aflatoxin B1—Induced hepatotoxicity alters gene expression and disrupts lipid and lipoprotein metabolism in rats. Toxicol. Rep. 2017, 4, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, S.; Kaya, E.; Karaca, A.; Karatas, O. Aflatoxin B1 induced renal and cardiac damage in rats: Protective effect of lycopene. Res. Vet. Sci. 2018, 119, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Meissonnier, G.; Pinton, P.; Laffitte, J.; Cossalter, A.-M.; Gong, Y.Y.; Wild, C.; Bertin, G.; Galtier, P.; Oswald, I. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmacol. 2008, 231, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Guindon, K.A.; Bedard, L.L.; Massey, T.E. Elevation of 8-Hydroxydeoxyguanosine in DNA from Isolated Mouse Lung Cells Following In Vivo Treatment with Aflatoxin B1. Toxicol. Sci. 2007, 98, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. Commission Regulation (EC) No 165/2010 of 26 February 2010 amending Regulation (EC) No 1881/2006 setting maximun levels of certain contaminants in foodstuffs as regards alatoxins. Off. J. Eur. Union L 2021, 50, 8–12. [Google Scholar]
- Bintvihok, A.; Thiengnin, S.; Doi, K.; Kumagai, S. Residues of aflatoxins in the liver, muscle and eggs of domestic fowls. J. Vet. Med. Sci. 2002, 64, 1037–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawal, S.; Kim, J.E.; Coulombe, R. Aflatoxin B1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Glahn, R.P.; Beers, K.W.; Bottje, W.G.; Wideman, R.F., Jr.; Huff, W.E. Altered renal function in broilers during aflatoxicosis. Poult. Sci. 1990, 69, 1796–1799. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Gonçalves, B.L.; de Neeff, D.V.; Ponzilacqua, B.; Coppa, C.F.S.C.; Hintzsche, H.; Sajid, M.; Cruz, A.G.; Corassin, C.H.; Oliveira, C.A.F. Aflatoxin in foodstuffs: Occurrence and recent advances in decontamination. Food Res. Int. 2018, 113, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Alpsoy, L.; Yildirim, A.; Agar, G. The antioxidant effects of vitamin A, C, and E on aflatoxin B1-induced oxidative stress in human lymphocytes. Toxicol. Ind. Health 2009, 25, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.S.; Wilcox, C.S. NADPH Oxidases in the Kidney. Antioxid. Redox Signal. 2006, 8, 1597–1607. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.C.; Srivastava, A.; Lall, R. Nutraceuticals in Veterinary Medicine; Springer International Publishing: Cham, Switzerland, 2019; Available online: https://link.springer.com/10.1007/978-3-030-04624-8 (accessed on 30 November 2020).



{kind=link}
{kind=link}
| Gene Name | Primer F Primer R | Amplicon Size | E | R2 | Acc. Number |
|---|---|---|---|---|---|
| NOX4 | TCGGGTGGCTTGTTGAAGTA-GTCTGTGGGAAATGAGCTTGG | 224 | 90 | 0.99 | NM_053524 |
| p47-phox | TACGCTGCTGTTGAAGAGGA-GATGTCCCCTTTCCTGACCA | 105 | 100 | 0.99 | AY029167.1 |
| BCL-2 | GCCTTCTTTGAGTTCGGTGG-CTGAGCAGCGTCTTCAGAGA | 221 | 100 | 0.99 | L14680.1 |
| 18S | AGAAACGGCTACCACATCCA-CCCTCCAATGGATCCTCGTT | 158 | 93 | 0.99 | NR_046237.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Longobardi, C.; Andretta, E.; Romano, V.; Lauritano, C.; Avantaggiato, G.; Schiavone, A.; Jarriyawattanachaikul, W.; Florio, S.; Ciarcia, R.; Damiano, S. Effects of Some New Antioxidants on Apoptosis and ROS Production in AFB1 Treated Chickens. Med. Sci. Forum 2021, 2, 12. https://doi.org/10.3390/CAHD2020-08640
Longobardi C, Andretta E, Romano V, Lauritano C, Avantaggiato G, Schiavone A, Jarriyawattanachaikul W, Florio S, Ciarcia R, Damiano S. Effects of Some New Antioxidants on Apoptosis and ROS Production in AFB1 Treated Chickens. Medical Sciences Forum. 2021; 2(1):12. https://doi.org/10.3390/CAHD2020-08640
Chicago/Turabian StyleLongobardi, Consiglia, Emanuela Andretta, Vincenzo Romano, Chiara Lauritano, Giuseppina Avantaggiato, Achille Schiavone, Watanya Jarriyawattanachaikul, Salvatore Florio, Roberto Ciarcia, and Sara Damiano. 2021. "Effects of Some New Antioxidants on Apoptosis and ROS Production in AFB1 Treated Chickens" Medical Sciences Forum 2, no. 1: 12. https://doi.org/10.3390/CAHD2020-08640