
Induction of Immune Response in Arabidopsis thaliana Treated with Phytopathogen Filtrates †


Abstract
:1. Introduction
2. Materials and Methods
In Planta Essays
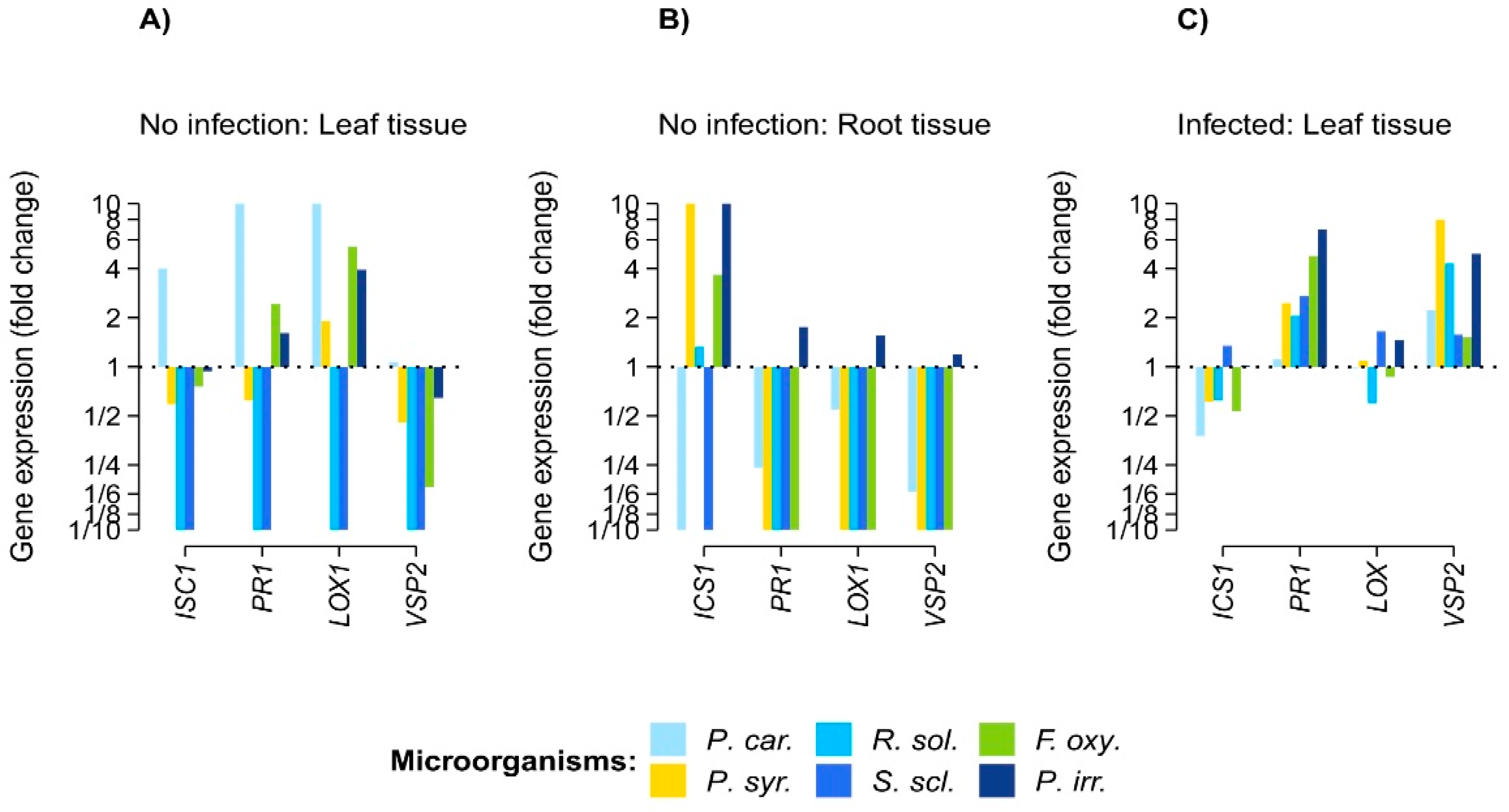
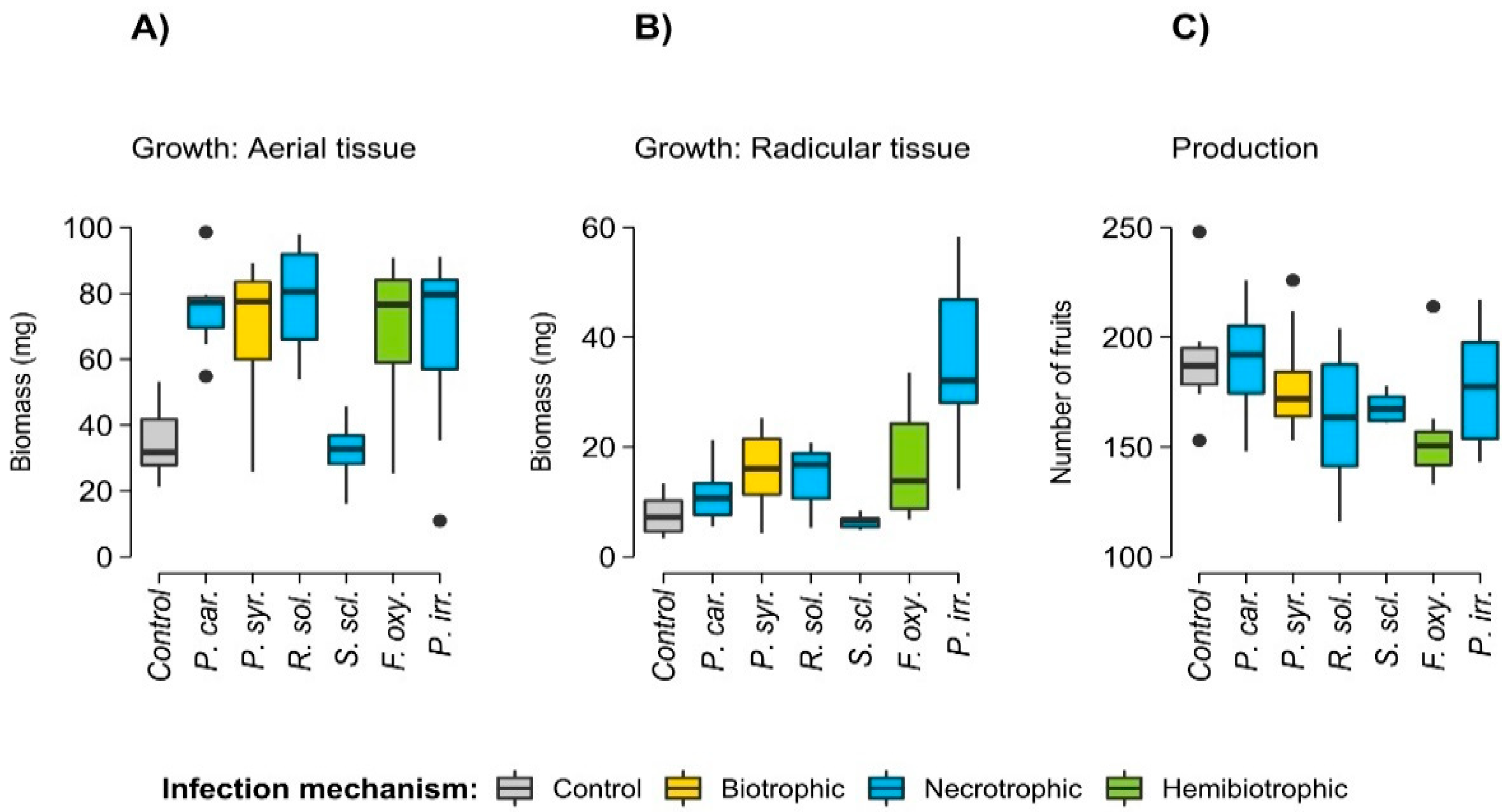
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Gene | Application | Sequences (5′-3′) |
|---|---|---|
| ICS1 | SA synthesis | GATCTAGCTAACGAGAACGG |
| ICS1 | SA synthesis | CATTAAACTCAACCTGAGGGAC |
| PR1 | SA response | CAAAGTGAGGTGTAACAATGGTGGA |
| PR1 | SA response | ATGGCTTCTCGTTCACATAATTCCC |
| LOX1 | JA synthesis | TCAACGATTTCAATGCTTCGTTTCT |
| LOX1 | JA synthesis | TCAGAGCTTACAAGACGAAGAGTG |
| VSP2 | JA response | GTTAGGGACCGGAGCATCAA |
| VSP2 | JA response | TCAATCCCGAGCTCTATGATGTT |
| Actin | Endogenous gene | CTCCCGCTATGTATGTCGCC |
| Actin | Endogenous gene | TTGGCACAGTGTGAGACACAC |
| Response Variable | Fixed Factor | Estimate ± S.E. | F | p Value |
|---|---|---|---|---|
| Root dry-weight (log10 transformed mg) (n = 49) | Phytopathogen filtrate: | 5.79 | <0.01 | |
| F. oxysporum | 0.41 ± 0.11 | |||
| P. carotovorum | 0.15 ± 0.11 | |||
| P. irregulare | 0.46 ± 0.11 | |||
| P. syringae | 0.23 ± 0.11 | |||
| R. solani | 0.32 ± 0.11 | |||
| S. sclerotiorum | −0.01 ± 0.11 | |||
| Aerial dry-weight | Phytopathogen filtrate: | 8.91 | <0.01 | |
| (n = 49) | F. oxysporum | 0.03 ± 0.01 | ||
| P. carotovorum | 0.03 ± 0.01 | |||
| P. irregulare | 0.02 ± 0.01 | |||
| P. syringae | 0.02 ± 0.01 | |||
| R. solani | 0.03 ± 0.01 | |||
| S. sclerotiorum | 0.01 ± 0.01 | |||
| Number of siliques | Phytopathogen filtrate: | 2.15 | <0.06 | |
| (n = 56) | F. oxysporum | −34.2 ± 12.7 | ||
| P. carotovorum | 0.5 ± 12.7 | |||
| P. irregulare | −13.2 ± 12.7 | |||
| P. syringae | −10.6 ± 12.7 | |||
| R. solani | −26.6 ± 12.7 | |||
| S. sclerotiorum | −22.1 ± 12.7 |
References
- Binyamin, R.; Nadeem, S.M.; Akhtar, S.; Khan, M.Y.; Anjum, R. Beneficial and pathogenic plant-microbe interactions: A review. Soil Environ. 2019, 38, 127–150. [Google Scholar] [CrossRef]
- Kamoun, S.; Wu, C.H.; Derevnina, L. Receptor networks underpin plant immunity. Science 2018, 360, 1300–1301. [Google Scholar]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Huang, Y.; Li, Y.; Dong, J.; Liu, X.; Li, C. Biocontrol of Rhizoctonia solani via Induction of the Defense Mechanism and Antimicrobial Compounds Produced by Bacillus subtilis SL-44 on Pepper (Capsicum annuum L.). Front. Microbiol. 2019, 10, 02676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, M.; Sohal, B.S. Role of Elicitors in Inducing Resistance in Plants against Pathogen Infection: A Review. ISRN Biochem. 2013, 2013, 762412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuc, J. Induced immunity to plant disease. BioScience 1982, 32, 854–860. [Google Scholar]
- Conrath, U.; Pieterse, C.; Mauch-Mani, B. Priming in plant–pathogen interactions. Trends Plant Sci. 2002, 7, 210–216. [Google Scholar] [CrossRef] [Green Version]
- da Silva, A.; Freitas, K.; Bastidas, J.; dos Santos, M.; Lilianne, M. Induction of defense mechanisms from filtrates of saprophytic fungi against early blight disease in tomato. Afr. J. Microbiol. Res. 2016, 10, 1849–1859. [Google Scholar]
- Dubery, I.; Sanabria, N.; Huang, J. Nonself perception in plant innate immunity. Adv. Exp. Med. Biol. 2012, 738, 79–107. [Google Scholar]
- Bae, S.J.; Mohanta, T.K.; Chung, J.Y.; Ryu, M.; Park, G.; Shim, S.; Hong, S.B.; Seo, H.; Bae, D.W.; Bae, I.; et al. Trichoderma metabolites as biological control agents against Phytophthora pathogens. Biol. Control 2016, 92, 128–138. [Google Scholar] [CrossRef]
- Stracquadanio, C.; Quiles, J.M.; Meca, G.; Cacciola, S.O. Antifungal activity of bioactive metabolites produced by trichoderma asperellum and trichoderma atroviride in liquid medium. J. Fungi 2020, 6, 263. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; da Silva, A.; Bastidas, J.; Pascholati, S.; Freitas, K. Induction of defense mechanisms in tomato plants by saprobic fungi filtrates against early blight disease1. Rev. Caatinga 2020, 33, 671–678. [Google Scholar]
- Malik, N.A.A.; Kumar, I.S.; Nadarajah, K. Elicitor and receptor molecules: Orchestrators of plant defense and immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.; Basu, A.; Kundu, S. Biotrophy-necrotrophy switch in pathogen evoke differential response in resistant and susceptible sesame involving multiple signaling pathways at different phases. Sci. Rep. 2017, 7, 17251. [Google Scholar] [CrossRef] [Green Version]
- Panthapulakkal, S.; Lung, S.C.; Liao, P.; Lo, C.; Chye, M.L. The overexpression of OsACBP5 protects transgenic rice against necrotrophic, hemibiotrophic and biotrophic pathogens. Sci. Rep. 2020, 10, 14918. [Google Scholar] [CrossRef]
- Poveda, J. Use of plant-defense hormones against pathogen-diseases of postharvest fresh produce. Physiol. Mol. Plant Pathol. 2020, 111, 101521. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Poveda, J. Investigación Básica y Aplicada En La Interacción Trichoderma-Brassicaceae. Ph.D. Thesis, Universidad de Salamanca, Salamanca-España, Spain, 2018. [Google Scholar]
- Livak, K.; Schmittgen, T. Analysis of Relative Gene Expression Data Using RealTime Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kassambara, A. Ggpubr R Package: Ggplot2-Based. Publication Ready Plots. 2019. Available online: http://www.sthda.com/english/articles/24-ggpubr-publication-ready-plots/ (accessed on 31 October 2020).
- McKinnon, S. Package Lemon: Freshing Up Your “Ggplot2” Plots. CRAN. 2020. Available online: https://cran.r-project.org/web/packages/lemon/lemon.pdf (accessed on 31 October 2020).
- Hossain, M.M.; Sultana, F.; Kubota, M.; Koyama, H.; Hyakumachi, M. The plant growth-promoting fungus Penicillium simplicissimum GP17-2 induces resistance in Arabidopsis thaliana by activation of multiple defense signals. Plant Cell Physiol. 2007, 48, 1724–1736. [Google Scholar] [CrossRef] [Green Version]
- de Wit, P. How plants recognize pathogens and defend themselves. Cell. Mol. Life Sci. 2007, 64, 2726–2732. [Google Scholar] [CrossRef]
- Bailey, B. Purification of a protein from culture filtrates of Fusarium oxysporum that induces ethylene and necrosis in leaves of Erythroxylum coca. Phytopathology 1995, 85, 1250–1255. [Google Scholar] [CrossRef]
- Tian, H.; Zhang, Y. The emergence of a mobile signal for systemic acquired resistance. Plant Cell 2019, 31, 1414–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Zhang, Y. Short- and long-distance signaling in plant defense. Plant J. 2021, 105, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Bouizgarne, B.; El-Maarouf-Bouteau, H.; Frankart, C.; Reboutier, D.; Madiona, K.; Pennarun, A.M.; Monestiez, M.; Trouverie, J.; Amiar, Z.; Briand, J.; et al. Early physiological responses of Arabidopsis thaliana cells to fusaric acid: Toxic and signalling effects. New Phytol. 2006, 169, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Betsuyaku, S.; Katou, S.; Takebayashi, Y.; Sakakibara, H.; Nomura, N.; Fukuda, H. Salicylic Acid and Jasmonic Acid Pathways are Activated in Spatially Different Domains around the Infection Site during Effector-Triggered Immunity in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Na, R.; Gijzen, M. Escaping Host Immunity: New Tricks for Plant Pathogens. PLoS Pathog. 2016, 12, e1005631. [Google Scholar] [CrossRef]
- Xin, X.-F.; Kvitko, B.; He, S. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef]
- Patel, Z.M.; Mahapatra, R.; Jampala, S.S.M. Role of fungal elicitors in plant defense mechanism. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Elsevier: Amsterdam, The Netherlands, 2020; pp. 143–158. [Google Scholar]
- Davies, D.R.; Bindschedler, L.V.; Strickland, T.S.; Bolwell, G.P. Production of reactive oxygen species in Arabidopsis thaliana cell suspension cultures in response to an elicitor from Fusarium oxysporum: Implications for basal resistance. J. Exp. Bot. 2006, 57, 1817–1827. [Google Scholar] [CrossRef] [Green Version]
- Osier, T.L.; Lindroth, R.L. Genotype and environment determine allocation to and costs of resistance in quaking aspen. Oecologia 2006, 148, 293–303. [Google Scholar] [CrossRef]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; AL-Harrasi, A. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef]
- de Souza, R.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef] [PubMed]
- Baroja-Fernández, E.; Almagro, G.; Sánchez-López, Á.M.; Bahaji, A.; Gámez-Arcas, S.; de Diego, N.; Dolezal, K.; Muñoz, F.J.; Climent Sanz, E.; Pozueta-Romero, J. Enhanced Yield of Pepper Plants Promoted by Soil Application of Volatiles From Cell-Free Fungal Culture Filtrates Is Associated With Activation of the Beneficial Soil Microbiota. Front. Plant Sci. 2021, 12, 752653. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, P.; Bahaji, A.; Gámez-Arcas, S.; Muñoz, F.J.; Sánchez-López, Á.M.; Almagro, G.; Baroja-Fernández, E.; Ameztoy, K.; de Diego, N.; Ugena, L.; et al. Volatiles from the fungal phytopathogen Penicillium aurantiogriseum modulate root metabolism and architecture through proteome resetting. Plant Cell Environ. 2020, 43, 2551–2570. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Calderón, E.; Aviles-Garcia, M.E.; Castulo-Rubio, D.Y.; Macías-Rodríguez, L.; Ramírez, V.M.; Santoyo, G.; López-Bucio, J.; Valencia-Cantero, E. Volatile compounds from beneficial or pathogenic bacteria differentially regulate root exudation, transcription of iron transporters, and defense signaling pathways in Sorghum bicolor. Plant Mol. Biol. 2018, 96, 291–304. [Google Scholar] [CrossRef]
- Fincheira, P.; Quiroz, A. Microbial volatiles as plant growth inducers. Microbiol. Res. 2018, 208, 63–75. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ávila, A.C.; Poveda, J. Induction of Immune Response in Arabidopsis thaliana Treated with Phytopathogen Filtrates. Biol. Life Sci. Forum 2022, 11, 85. https://doi.org/10.3390/IECPS2021-11974
Ávila AC, Poveda J. Induction of Immune Response in Arabidopsis thaliana Treated with Phytopathogen Filtrates. Biology and Life Sciences Forum. 2022; 11(1):85. https://doi.org/10.3390/IECPS2021-11974
Chicago/Turabian StyleÁvila, Ana Cristina, and Jorge Poveda. 2022. "Induction of Immune Response in Arabidopsis thaliana Treated with Phytopathogen Filtrates" Biology and Life Sciences Forum 11, no. 1: 85. https://doi.org/10.3390/IECPS2021-11974