
Comparative Taxonomic Study of Balanites aegyptiaca (L.) Delile (Zygophyllaceae) †


 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Statistical Analysis
2.2. Anatomical Study and Scanning Electron Microscope (SEM)
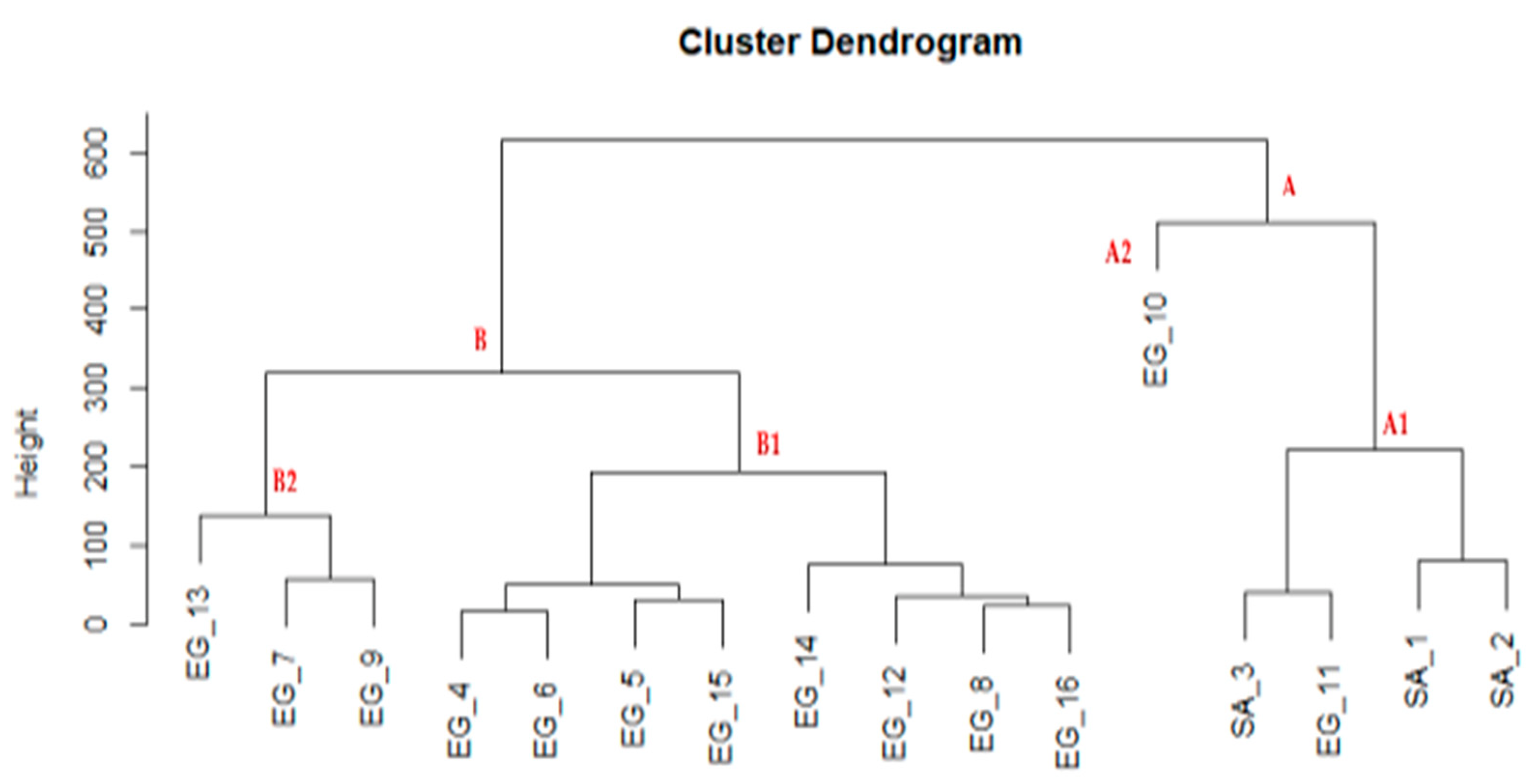
3. Results
3.1. Vegetative and Fruit Macromorphology
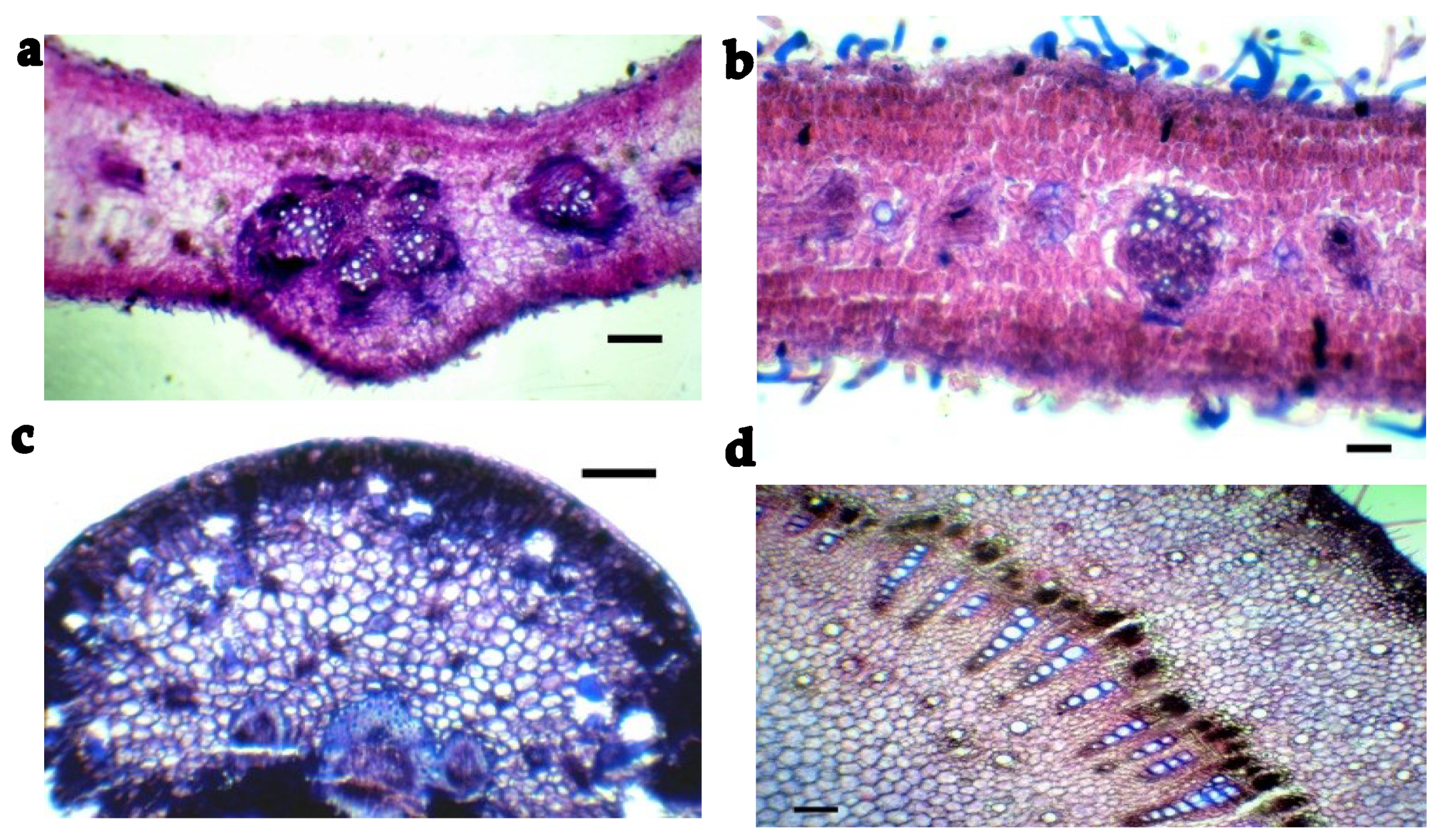
3.2. Anatomical Characterization
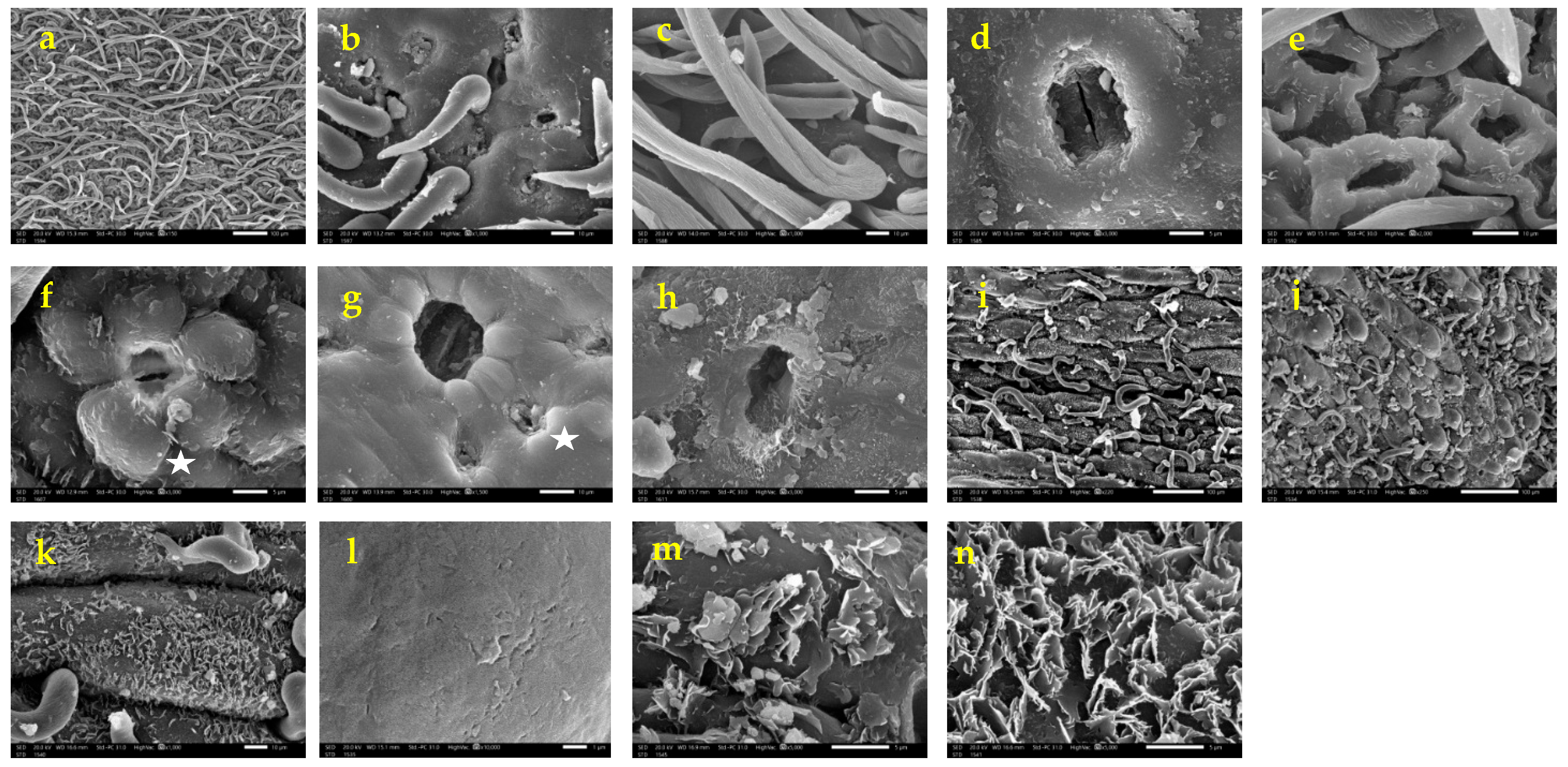
3.3. Vegetative and Fruit Micromorphology (SEM)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| ID | Collection Sites | Date | Latitude | Longitude |
|---|---|---|---|---|
| SA1 | South Al Jumum, Makkah, Saudi Arabia | 3 April 2021 | 21°34′27.0″ N | 39°41′16.4″ E |
| SA 2 | North Al Jumum, Makkah, Saudi Arabia | 5 April 2021 | 21°35′56.1″ N | 39°42′11.3″ E |
| SA 3 | North Al Jumum, Makkah, Saudi Arabia | 20 May 2021 | 21°38′51.5″ N | 39°41′06.8″ E |
| EG4 | Wadi el-Gemal National Park, Marsa Alam, Red Sea, Egypt | 6 March 2021 | 24°38′30.88″ N | 35°2′25.55″ E |
| EG 5 | Wadi el-Gemal National Park, Marsa Alam, Red Sea, Egypt | 6 March 2021 | 24°37′50.58″ N | 35°1′43.186″ E |
| EG 6 | Wadi el-Gemal National Park, Marsa Alam, Red Sea, Egypt | 6 March 2021 | 24°37′37.03″ N | 35°1′16.50″ E |
| EG 7 | Wadi el-Gemal National Park, Marsa Alam, Red Sea, Egypt | 6 March 2021 | 24°37′33.68″ N | 35°1′11.06″ E |
| EG 8 | Wadi el-Gemal National Park, Marsa Alam, Red Sea, Egypt | 6 March 2021 | 24°41′12.8″ N | 35°4′59.64″ E |
| EG 9 | Wadi el-Gemal National Park, Marsa Alam, Red Sea, Egypt | 6 March 2021 | 25°4′48.55″ N | 34°53′12.22″ E |
| EG 10 | Marsa Alam, Red Sea, Egypt | 8 March 2021 | 25°04′43.3″ N | 34°53′14″ E |
| EG 11 | Edfu- Marsa Alam Road, Edfu, Red Sea, Egypt | 8 March 2021 | 25°1′39.18″ N | 33°3′13.79″ E |
| EG 12 | Edfu- Marsa Alam Road, Edfu, Red Sea, Egypt | 15 April 2021 | 25°1′39.40″ N | 33°3′14.08″ E |
| EG 13 | Edfu, Aswan, Egypt | 15 April 2021 | 25°1′39.92″ N | 33°3′12.54″ E |
| EG 14 | High Dam region, Aswan, Egypt | 30 April 2021 | 23°58′36.69″ N | 32°53′50.28″ E |
| EG 15 | High Dam region, Aswan, Egypt | 30 April 2021 | 23°58′41.26″ N | 32°53′53.52″ E |
| EG 16 | Aswan University, Aswan, Egypt | 30 April 2021 | 23°59′52.21″ N | 32°51′37.07″ E |
| Variables | Mean ± StDev | SE Mean | Minimum | Q1 | Median | Q3 | Maximum | IQR |
|---|---|---|---|---|---|---|---|---|
| Leaflet length (cm) | 2.693 ± 1.22 | 0.227 | 0.89 | 1.927 | 2.415 | 3.083 | 6.4 | 1.155 |
| Leaflet width (cm) | 1.273 ± 0.736 | 0.137 | 0.331 | 0.73 | 1 | 1.585 | 3.55 | 0.855 |
| Leaflet length/width ratio | 2.321 ± 0.625 | 0.116 | 1.268 | 1.839 | 2.368 | 2.774 | 3.636 | 0.935 |
| Leaflet area (cm2) | 2.864 ± 3.167 | 0.588 | 0.21 | 1.093 | 2.04 | 3.045 | 14.845 | 1.952 |
| Petiole length (cm) | 0.3936 ± 0.2466 | 0.0458 | 0.1 | 0.185 | 0.3 | 0.549 | 0.99 | 0.364 |
| Petiolule length (cm) | 0.377 ± 0.571 | 0.202 | 0.06 | 0.064 | 0.092 | 0.71 | 1.6 | 0.646 |
| Apex angle | 114.77 ± 21.38 | 4.04 | 58.5 | 107.13 | 117 | 127 | 171 | 19.88 |
| Base angle | 59.21 ± 26.36 | 4.89 | 26.5 | 39.25 | 54.5 | 69 | 124.5 | 29.75 |
| Stipule length (cm) | 0.2072 ± 0.0546 | 0.0223 | 0.156 | 0.156 | 0.2025 | 0.2545 | 0.28 | 0.0985 |
| Interpetiolar stipule (cm) | 0.5594 ± 0.1715 | 0.0767 | 0.275 | 0.4015 | 0.624 | 0.685 | 0.685 | 0.2835 |
| Spine length (cm) | 1.711 ± 1.291 | 0.358 | 0.42 | 0.662 | 1.37 | 2.457 | 4.7 | 1.794 |
| Spinule length (cm) | 0.4988 ± 0.2335 | 0.0953 | 0.1662 | 0.3328 | 0.461 | 0.7195 | 0.836 | 0.3867 |
| Fruit length (cm) | 3.0852 ± 0.3596 | 0.0848 | 2.438 | 2.819 | 3.048 | 3.4275 | 3.664 | 0.6085 |
| Fruit width (cm) | 1.751 ± 0.506 | 0.119 | 0.997 | 1.334 | 1.711 | 1.924 | 2.712 | 0.59 |
| Fruit length/width ratio | 1.5792 ± 0.2597 | 0.0612 | 1.2142 | 1.3712 | 1.5348 | 1.8121 | 2.1509 | 0.4409 |
| Fruit area (cm2) | 4.843 ± 1.88 | 0.443 | 2.59 | 3.521 | 4.229 | 6.137 | 9.49 | 2.616 |
References
- Abdelaziz, S.M.; Medraoui, L.; Alami, M.; Pakhrou, O.; Makkaoui, M.; Boukhary, A.O.M.S.; Filali-Maltouf, A. Inter simple sequence repeat markers to assess genetic diversity of the desert date (Balanites aegyptiaca Del.) for Sahelian ecosystem restoration. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sands, M.J.S. The desert date and its relatives: A revision of the genus Balanites. Kew Bull. 2001, 56, 1–128. [Google Scholar] [CrossRef]
- Amer, W.; Soliman, M.; Sheded, M. Biosystematic studies for Balanites aegyptiaca (Balanitaceae) populations in Egypt. Flora Mediterr. 2002, 12, 353–367. [Google Scholar]
- Alpinio, P. Prosperi Alpini De plantis Aegypti liber. Venetiis Apud Fr. De Fr. Senensem 1592, 4, 80. [Google Scholar]
- Hansen, C.; Maule, A.F. Pehr ’Osbeck’s collections and Linnaeus’s Species Plantarum (1753). Bot. J. Linn. Soc. 1973, 67, 189–212. [Google Scholar] [CrossRef]
- De Roziere, M. Description Minéralogique de la Vallée de Qoceyr in Description de l’Egypte, Histoire Naturelle; L’Imprimerie Impériale: Paris, France, 1813; Volume 2, pp. 83–98. [Google Scholar]
- Hegnauer, R. Chemotaxonomy of plants: An overview of the distribution and systematic significance of plant materials. In Bd 6, Dicotyledoneae: Rafflesiaceae-Zygophyllaceae; Birkhäuser: Basel, Switzerland, 1973. [Google Scholar]
- Cronquist, A.; Takhtadzhian, A.L. An Integrated System of Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Sands, M. Flora of Ethiopia. Balanitaceae; Edwards, S., Ed.; Uppsala University: Uppsala, Sweden, 1989; Volume 3, pp. 433–436. [Google Scholar]
- Boesewinkel, F.D. Ovule and seed characters of Balanites aegyptiaca and the classification of the Linales-Geraniales-Polygalales assembly. Acta Bot. Neerl. 1994, 43, 15–25. [Google Scholar] [CrossRef]
- Singh, B.P.; Kaur, I.; Gauchan, D.P. Floral anatomy and systematic position of the genus Balanites. Acta Bot. Hungarica. 2002, 1–2, 137–143. [Google Scholar] [CrossRef]
- Sheahan, M.C.; Chase, M.W. Phylogenetic relationships within Zygophyllaceae based on DNA sequences of three plastid regions, with special emphasis on Zygophylloideae. Syst. Bot. 2000, 25, 371. [Google Scholar] [CrossRef]
- Godoy-Bürki, A.C.; Acosta, J.M.; Aagesen, L. Phylogenetic relationships within the New World subfamily Larreoideae (Zygophyllaceae) confirm polyphyly of the disjunct genus Bulnesia. Syst. Biodivers. 2018, 16, 453–468. [Google Scholar] [CrossRef]
- Maksoud, S.A.; El Hadidi, M.N. The flavonoids of Balanites aegyptiaca (Balanitaceae) from Egypt. Plant Syst. Evol. 1988, 160, 153–158. [Google Scholar] [CrossRef]
- Boulos, L. Flora of Egypt Checklist, Revised Annotated Edition; Al-Hadara Publishing: Cairo, Egypt, 2009; pp. 198–201. [Google Scholar]
- Migahid, A. Flora of Saudi Arabia; King Saud University Press: Riyadh, Saudi Arabia, 1974. [Google Scholar]
- Smith, T. Plant Systematics by Michael G. Simpson. Syst. Bot. 2006, 31, 631–632. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 15 November 2021).
- McCracken, E.; Johansen, D.A. Plant Microtechnique. Trans. Am. Microsc. Soc. 1940, 59, 405. [Google Scholar] [CrossRef]
- Barthlott, W. Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects. Nord. J. Bot. 1981, 1, 345–355. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Chernetskyy, M. Structure of trichomes from the surface of leaves of some species of Kalanchoë Adans. Acta Biol. Cracov. Ser. Bot. 2005, 47, 15–22. [Google Scholar]
- Koch, K.; Barthlott, W. Plant epicuticular waxes: Chemistry, form, self-assembly and function. Nat. Prod. Commun. 2006, 1, 1067–1072. [Google Scholar] [CrossRef]
- Khokhar, A.L.; Rajput, M.T.; Tahir, S.S. Toxonomic study of the trichomes in the some members of the genus Convolvulus (Convolvulaceae). Pak. J. Bot. 2012, 44, 1219–1224. [Google Scholar]
- El-Banhawy, A.H.; Elkordy, A.; Farag, R.; Elbar, O.A.; Faried, A.; Ellamouni, F. Taxonomic significance of the leaf geometric and micrometric attributes in the discrimination of some cultivars of Mangifera indica L. (Anacardiaceae). Egypt. J. Bot. 2020, 61, 255–269. [Google Scholar] [CrossRef]
- El-Banhawy, A.; Al-Juhani, W. DNA barcoding and phylogeny of Phlomis aurea (Lamiaceae) endemic to Sinai peninsula, Egypt. Pak. J. Bot. 2019, 51, 1263–1271. [Google Scholar] [CrossRef]
- Faried, A.; El-Banhawy, A.; Elqahtani, M. Taxonomic, DNA barcoding and phylogenetic reassessment of the Egyptian Ephedra L. (Ephedraceae). Catrina Int. J. Environ. Sci. 2018, 17, 1–13. [Google Scholar] [CrossRef]
- El-Banhawy, A.; Kamel, W.M.; El-Din, E.M.G. Salvia lanigera var. grandiflora Benth: A new record in the Flora of Egypt. J. Ecol. Health Environ. 2016, 4, 87–89. [Google Scholar] [CrossRef]
- El-Banhawy, A.; Nour, I.; Acedo, C.; ElKordy, A.; Faried, A.; Al-Juhani, W.; Gawhari, A.; Olwey, A.; Ellmouni, F. Taxonomic revisiting and phylogenetic placement of two endangered plant species: Silene leucophylla Boiss. and Silene schimperiana Boiss. (Caryophyllaceae). Plants 2021, 10, 740. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamada, F.A.; El-Banhawy, A.; Ellmouni, F.Y.; Al-Juhani, W.; Makharita, R.R.; Nour, I.H. Comparative Taxonomic Study of Balanites aegyptiaca (L.) Delile (Zygophyllaceae). Biol. Life Sci. Forum 2022, 11, 72. https://doi.org/10.3390/IECPS2021-12060
Hamada FA, El-Banhawy A, Ellmouni FY, Al-Juhani W, Makharita RR, Nour IH. Comparative Taxonomic Study of Balanites aegyptiaca (L.) Delile (Zygophyllaceae). Biology and Life Sciences Forum. 2022; 11(1):72. https://doi.org/10.3390/IECPS2021-12060
Chicago/Turabian StyleHamada, Fatma A., Ahmed El-Banhawy, Faten Y. Ellmouni, Widad Al-Juhani, Rabab R. Makharita, and Iman H. Nour. 2022. "Comparative Taxonomic Study of Balanites aegyptiaca (L.) Delile (Zygophyllaceae)" Biology and Life Sciences Forum 11, no. 1: 72. https://doi.org/10.3390/IECPS2021-12060