
Elucidating the Role of Inositol Monophosphatases Gene Family in Abiotic Stress Management †


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Chickpea IMPL Genes along with Its cDNA and Sequence Study
2.2. Expression Pattern of IMP Gene Family in Different Environmental Stresses and Different Tissues/Organs
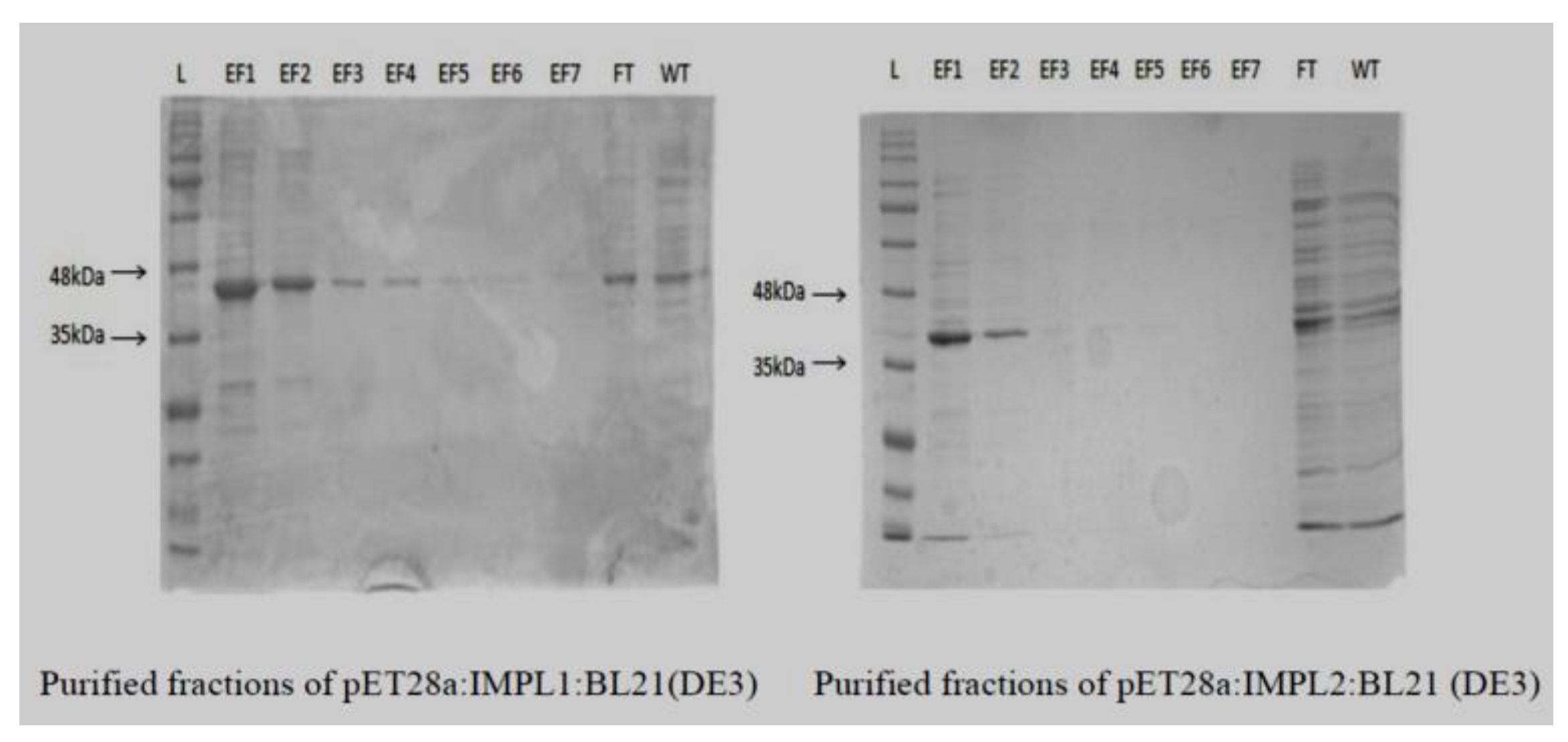
2.3. Bacterial Overexpression, Purification, and Biochemical Characterization of CaIMPL1 and CaIMPL2 Gene(s) Product
3. Results
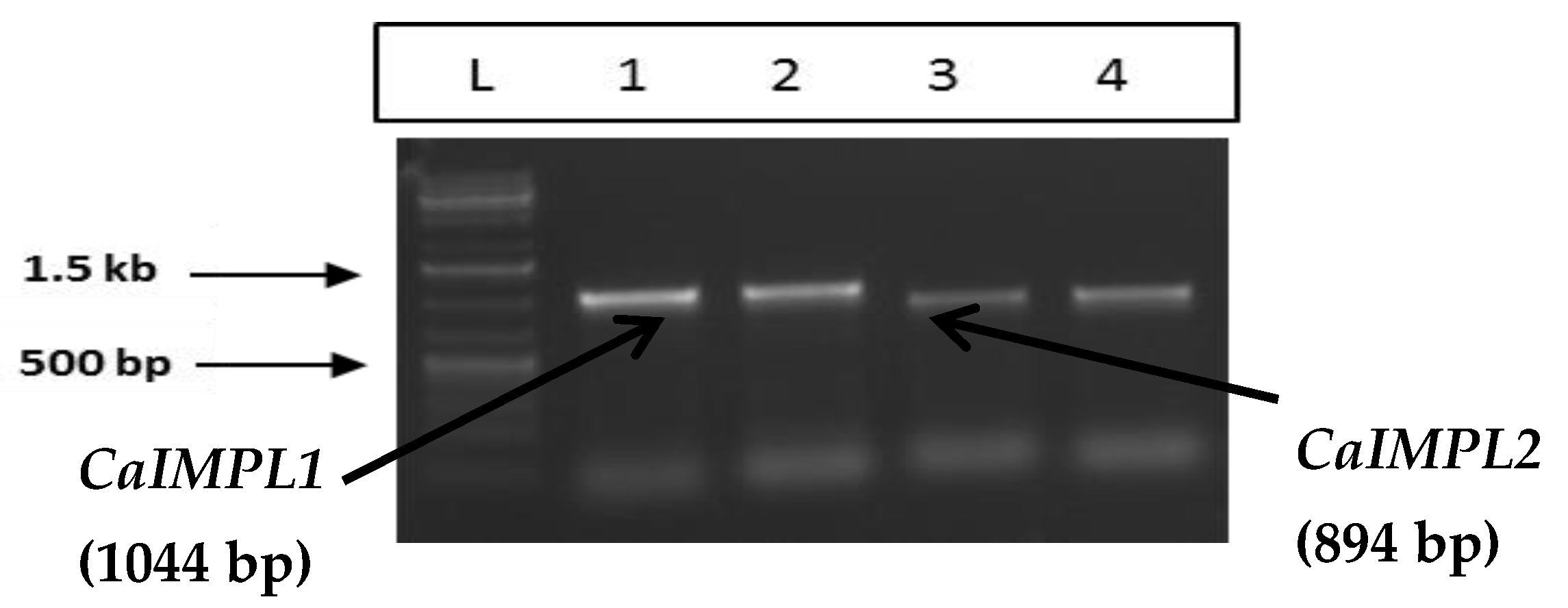
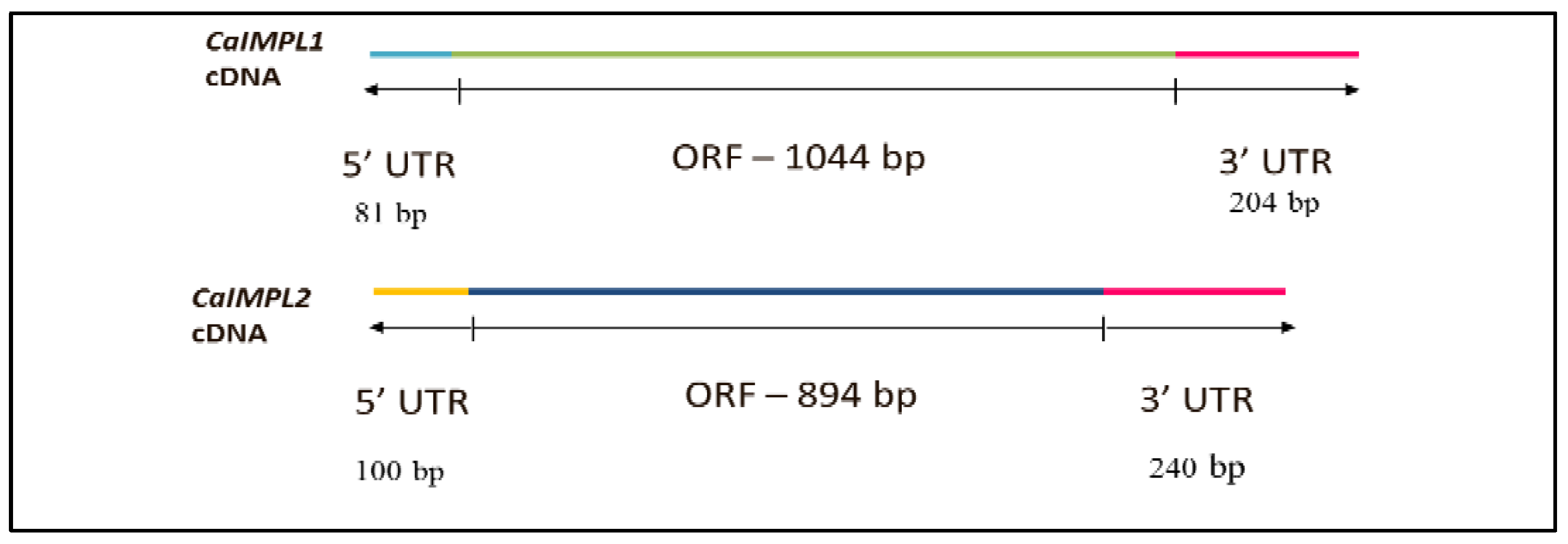
3.1. Molecular Cloning, Sequencing, and Characterization of CaIMPL1 and CaIMPL2 Genes and cDNA(s)
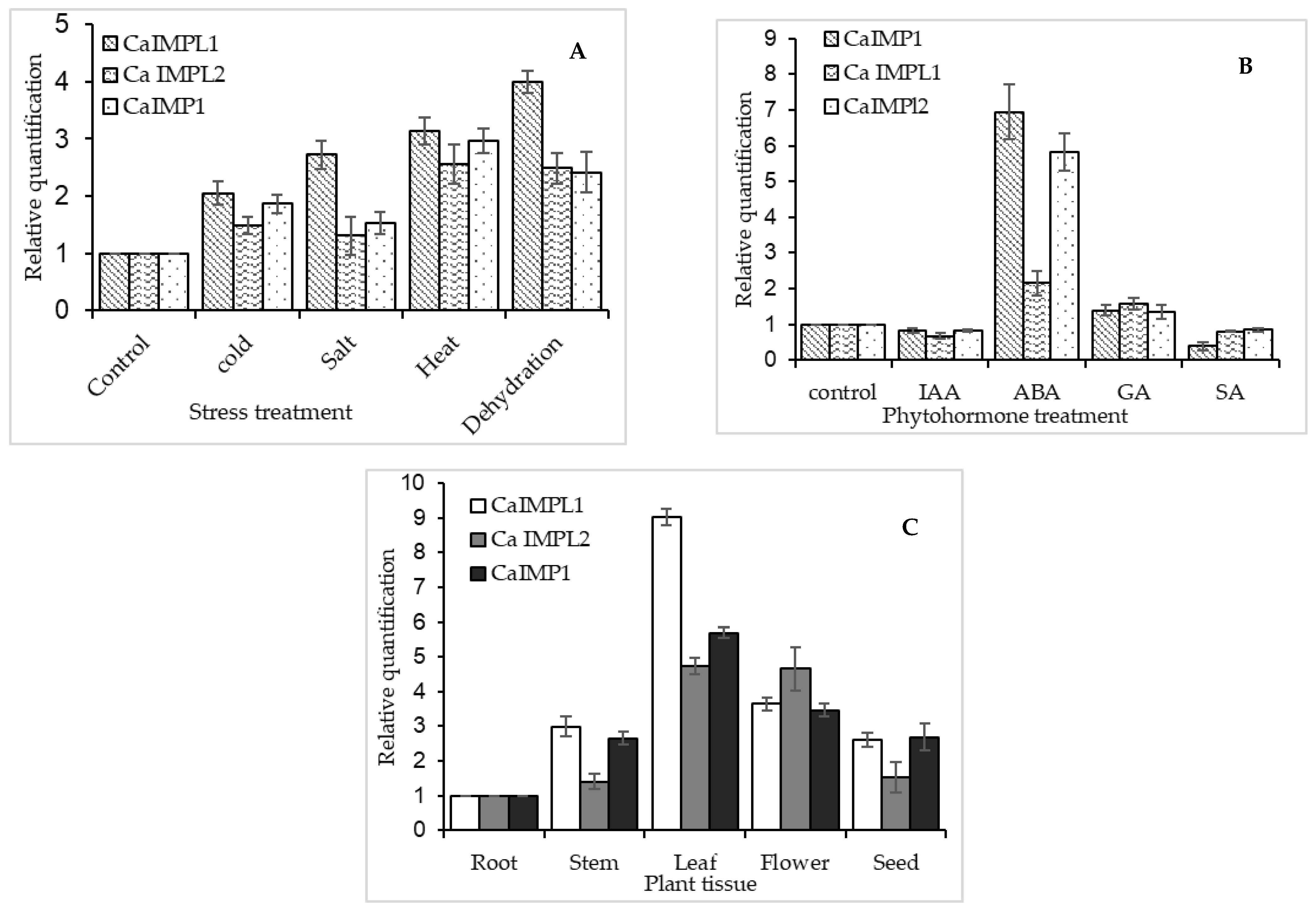
3.2. IMP Gene Family Transcript Differentially Expressed in Different Environmental Stresses, in Presence of Different Hormones, and in Different Tissues/Organs
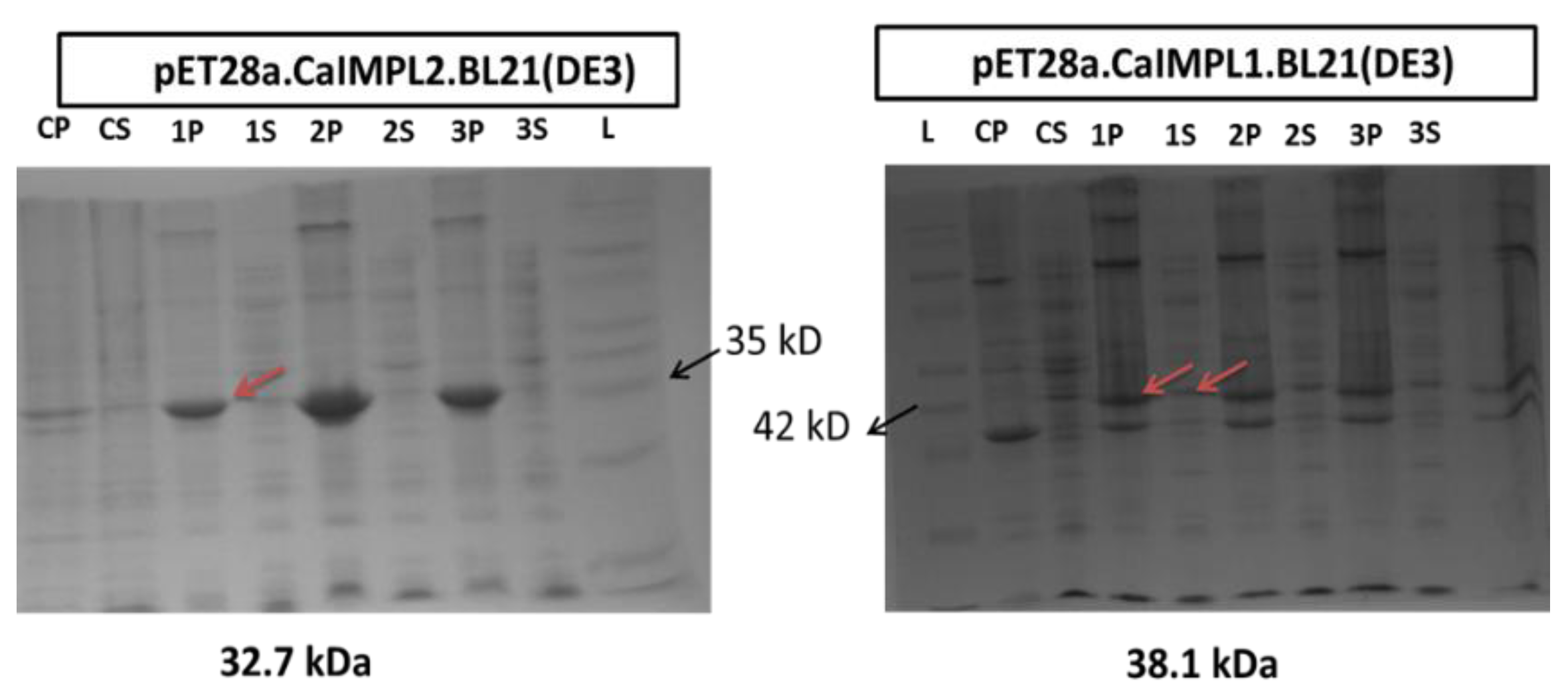
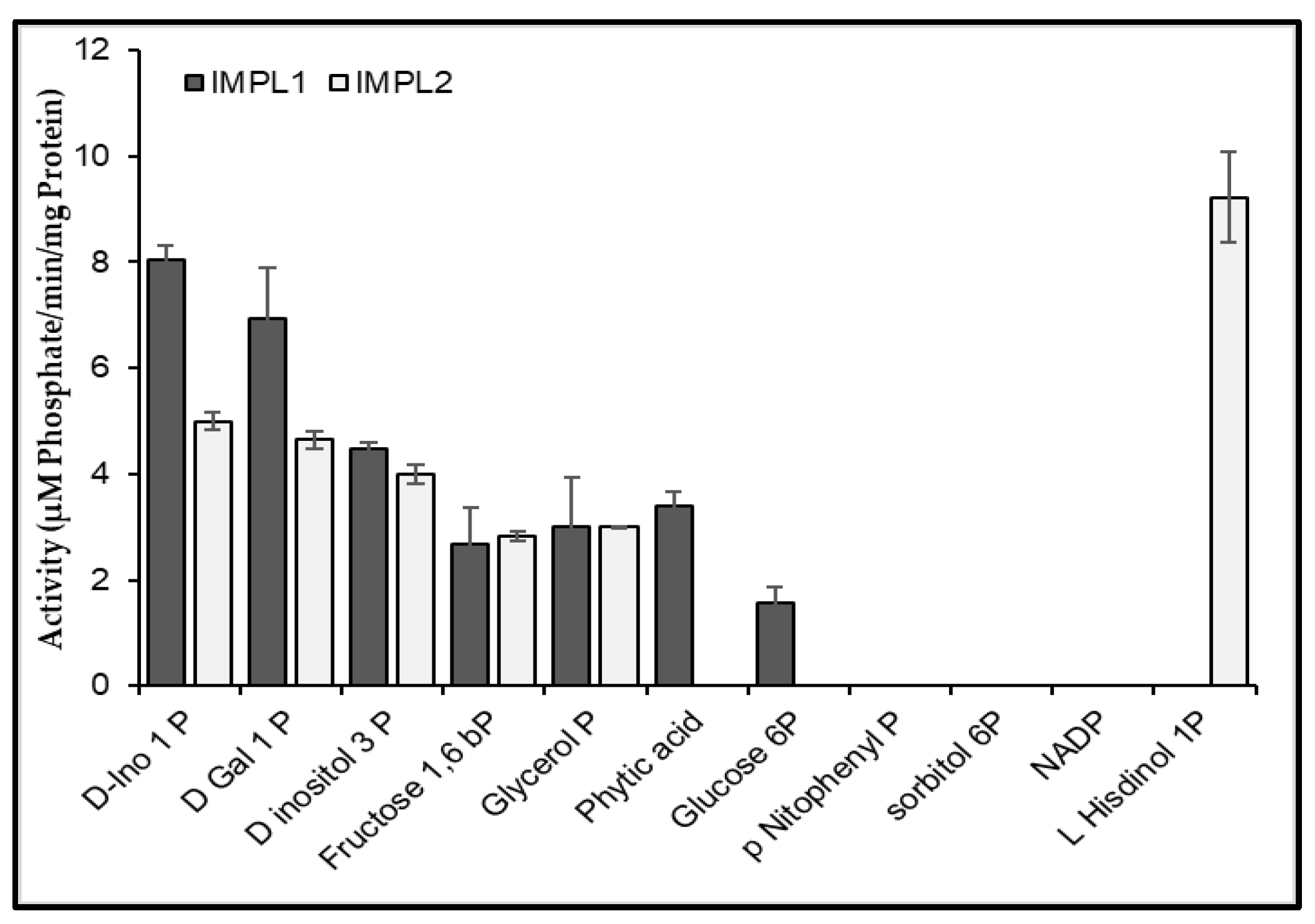
3.3. CaIMPL1 and CaIMPL2 Proteins Exhibit Distinct Biochemical Properties
4. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer No. | Primer Sequence (5′-3′) | Purpose |
|---|---|---|
| CaSS1F | ATGTTGTCACAGTGCCATCT | For cloning full-length CDS of CaIMPL2 sequence |
| CaSS2R | GCTCTAGATTCATTACAACGGTAA | |
| CaSS3F | ATGTCAATTGTATTCTCCGCAGC | For cloning full-length CDS of CaIMPL2 sequence |
| CaSS4R | GAGGACTACAGGGCAGACGTTTAA | |
| CaSS5F | GAATTCATGTTGTCACAGTGCCATCT | To clone full-length CaIMPL2 in bacterial expression vector |
| CaSS6R | CTCGAGCCGTTGTAATGAATCTAGAGC | |
| CaSS7F | GAATTCATGTCAATTGTATTCTCCGCAGC | To clone full-length CDS of CaIMPL1 in bacterial expression vector |
| CaSS8R | CTCGAGAACGTCTGCCCTGTAGTCCTC | |
| CaSS9F | GTGTACGCCAATCCTTGTGAAC | For real-time PCR in chickpea for CaIMPL2 |
| CaSS10R | CGACTACTCACGATTCCGCATA | |
| CaSS11F | CTCGGATGGATGGAGGAAAA | For real-time PCR in chickpea for CaIMPL1 |
| CaSS12R | GGAGGACGCCGTTTGAAA | |
| CaSS13F | AGCGTGTAGCTGCTTCAAACC | For real-time PCR in chickpea for CaIMP |
| CaSS14R | GTTTGGCGCAGAGCATCA | |
| CaSS15F | GCCCGCGACGTTGTGA | For real-time PCR for chickpea 18S (Endogenous control) |
| CaSS16R | CCTTGTTACGACTTCTCCTTCCTCTA |
References
- Boominathan, P.; Shukla, R.; Kumar, A.; Manna, D.; Negi, D.; Verma, P.K.; Chattopadhyay, D. Long Term Transcript Accumulation during the Development of Dehydration Adaptation in Cicer arietinum. Plant Physiol. 2004, 135, 1608–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.J.; Jauhar, P.P. Genetic Resources, Chromosome Engineering, and Crop Improvement; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar] [CrossRef]
- Saxena, S.C.; Salvi, P.; Kaur, H.; Verma, P.; Petla, B.P.; Rao, V.; Kamble, N.U.; Majee, M. Differentially expressed myo-inositol monophosphatase gene (CaIMP) in chickpea (Cicer arietinum L.) encodes a lithium-sensitive phosphatase enzyme with broad substrate specificity and improves seed germination and seedling growth under abiotic stresses. J. Exp. Bot. 2013, 64, 5623–5639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Russell, D.W. Expression of cloned genes in Escherichia coli. In Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001; Volume 3, pp. 15.23–15.24. [Google Scholar]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Majee, M.; Maitra, S.; Dastidar, K.G.; Pattnaik, S.; Chatterjee, A.; Hait, N.C.; Das, K.P.; Majumder, A.L. A Novel Salt-tolerant l-myo-Inositol-1-phosphate Synthase from Porteresia coarctata (Roxb.) Tateoka, a Halophytic Wild Rice. J. Biol. Chem. 2004, 279, 28539–28552. [Google Scholar] [CrossRef] [PubMed] [Green Version]






| Enzyme (Substrate) | Km (μM) | Vmax (μmol min−1 mg−1) | MgCl2 (mM) | Optimum Temperature | Optimum pH | |
|---|---|---|---|---|---|---|
| 1 | CaIMPL2 (Histidinol 1-P) | 29 | 3.8 | 3 | 37 °C | 8 |
| 2 | CaIMPL1 (D-Inositol 1-Phosphate) | 24 | 4.1 | 3 | 37 °C | 8 |
| 3 | CaIMPL1 (D-Galactose 1-Phosphate) | 18 | 4.9 | 3 | 37 °C | 8 |
| 4 | CaIMP (D-Inositol 1-Phosphate) | 25 | 4.4 | 3 | 37 °C | 8 |
| 5 | CaIMP (D-Galactose 1-Phosphate) | 16 | 5.3 | 3 | 37 °C | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saxena, S.C. Elucidating the Role of Inositol Monophosphatases Gene Family in Abiotic Stress Management. Biol. Life Sci. Forum 2022, 11, 70. https://doi.org/10.3390/IECPS2021-12055
Saxena SC. Elucidating the Role of Inositol Monophosphatases Gene Family in Abiotic Stress Management. Biology and Life Sciences Forum. 2022; 11(1):70. https://doi.org/10.3390/IECPS2021-12055
Chicago/Turabian StyleSaxena, Saurabh Chandra. 2022. "Elucidating the Role of Inositol Monophosphatases Gene Family in Abiotic Stress Management" Biology and Life Sciences Forum 11, no. 1: 70. https://doi.org/10.3390/IECPS2021-12055