
Differential Tolerance of Cowpea Cultivars to Osmotic Stress in Germinative Phase: A Multivariate Approach †

 ,
,
Abstract
:1. Introduction
2. Material and Methods
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vijaykumar, A.; Saini, A.; Jawali, N. Phylogenetic analysis of subgenus vigna species using nuclear ribosomal RNA ITS: Evidence of hybridization among Vigna unguiculata subspecies. J. Hered. 2010, 101, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vale, J.C.; Bertini, C.; Borém, A. Feijão-Caupi do Plantio à Colheita, 1st ed.; Editora UFV: Viçosa, Brazil, 2017. [Google Scholar]
- Nascimento, S.P.; Bastos, E.A.; Araújo, E.C.E.; Freire Filho, F.R.; Silva, E.M. Tolerância ao déficit hídrico em genótipos de feijão-caupi. Rev. Bras. Eng. Agrícola E Ambient. 2011, 15, 853–861. [Google Scholar] [CrossRef]
- Freitas, R.M.O.; Dombroski, J.L.D.; Freitas, F.C.L.; Nogueira, N.W.; Pinto, J.R.S. Physiological responses of cowpea under water stress and rewatering in no-tillage and conventional tillage systems. Rev. Caatinga 2017, 30, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Możdżeń, K.; Bojarski, B.; Rut, G.; Migdałek, G.; Repka, P.; Rzepka, A. Effect of drought stress induced by mannitol on physiological parameters of maize (Zea mays L.) seedlings and plants. J. Microbiol. Biotechnol. Food Sci. 2015, 4, 86–91. [Google Scholar] [CrossRef]
- Oliveira, A.B.; Gomes Filho, E.; Enéas Filho, J. O problema da salinidade na agricultura e as adaptações das plantas ao estresse salino. Enciclopédia Biosf. 2010, 6, 1–16. [Google Scholar]
- Colman, B.A.; Nunes, C.M.; Masson, G.L.; Barbosa, R.H.; Nunes, A.S. Indução de tolerância ao estresse hídrico na germinação de sementes de feijão-caupi. Comun. Sci. 2014, 5, 449–455. [Google Scholar] [CrossRef]
- Vicini, L.; Souza, A.M.; Morales, F.E.C.; Souza, F.M. Técnicas Multivariadas Exploratórias: Teorias e Aplicações No Software Statistica; Editora UFSM: Santa Maria, Brazil, 2018; p. 240. [Google Scholar]
- Manly, B.F.J.; Alberto, J.A.N. Métodos Estatísticos Multivariados: Uma Introdução, 4th ed.; Bookman: Porto Alegre, Brazil, 2019; p. 254. [Google Scholar]
- Villela, F.A.; Filho, L.D.; Sequeira, E.L. Tabela de potencial osmótico em função da concentração de polietileno-glicol 6.000 e da temperatura. Pesqui. Agropecuária Bras. 1991, 26, 1957–1968. [Google Scholar]
- Brasil Ministério da Agricultura, Pecuária e Abastecimento. Regras Para Análise de Sementes; Secretaria de Defesa Agropecuária/Mapa/ACS: Brasília, Brazil, 2009.
- Rosa, L.S.; Felippi, M.; Nogueira, A.C.; Grossi, F. Avaliação da germinação sob diferentes potenciais osmóticos e caracterização morfológica da semente e plântula de Ateleia glazioviana Baill (timbó). Cerne 2005, 11, 306–314. [Google Scholar]
- Labouriau, L.G. A Germinação das Sementes, 24th ed.; OEA: Washington, DC, USA, 1983; p. 174. [Google Scholar]
- Maguire, J.D. Speed of germination-aid in selection and evaluation for seedling emergence and vigor. Crop Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- Nakagawa, J. Testes de vigor baseados no desempenho das plântulas. In Vigor de sementes: Conceitos e Testes; KrzyzanoskI, F.C., Vieira, R.D., França Neto, J.B., Eds.; ABRATES: Londrina, Brazil, 1999; pp. 20–31. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Kaiser, H.F. The varimax criterion for analytic rotation in fator analysis. Psychometrika 1958, 23, 187–200. [Google Scholar] [CrossRef]
- Scalon, S.P.Q.; Mussury, R.M.; Euzébio, V.L.D.M.; Kodama, F.M.; Kissmann, C. Estresse hídrico no metabolismo e crescimento inicial de mudas de mutambo (Guazuma ulmifolia Lam.). Ciência Florest. 2011, 21, 655–662. [Google Scholar] [CrossRef]
- Marcos Filho, J. Fisiologia de Sementes de Plantas Cultivadas, 5th ed.; Abrates: Londrina, Brazil, 2015; p. 659. [Google Scholar]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Fisiologia e Desenvolvimento Vegetal, 6th ed.; Artmed: Porto Alegre, Brazil, 2017. [Google Scholar]
- Carneiro, M.M.L.C.; Deuner, S.; Oliveira, P.V.; Teixeira, S.B.; Sousa, C.P.; Bacarin, M.A.; Moraes, D.M. Atividade antioxidante e viabilidade de sementes de girassol após estresse hídrico e salino. Rev. Bras. Sementes 2011, 33, 752–761. [Google Scholar] [CrossRef]
- Girotto, L.; Alves, J.D.; Deuner, S.; Albuquerque, A.C.S.; Tomazoni, A.P. Tolerância à seca de genótipos de trigo utilizando agentes indutores de estresse no processo de seleção. Rev. Ceres 2012, 59, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.A.; Silva, N.V.; Walter, L.S.; Silva, E.C.A.; Nogueira, R.J.M.C. Germinação de duas espécies da caatinga sob déficit hídrico e salinidade. Pesqui. Florest. Bras. 2016, 36, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Pelegrini, L.L.; Borcioni, E.; Nogueira, A.C.; Koehler, H.S.; Quoirin, M.G.G. Efeito do estresse hídrico simulado com NaCl, manitol e PEG (6000) na germinação de sementes de Erythrina falcata Benth. Ciência Florest. 2013, 23, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.B.; Gomes Filho, E. Efeito do condicionamento osmótico na germinação e vigor de sementes de sorgo com diferentes qualidades fisiológicas. Rev. Bras. Sementes 2010, 32, 25–34. [Google Scholar] [CrossRef]
- Agostini, E.A.T.; Machado-neto, N.B.; Custódio, C.C. Induction of water deficit tolerance by cold shock and salicylic acid during germination in the common bean. Acta Scientiarum. Agron. 2013, 35, 209–219. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; p. 392. [Google Scholar]
- Machado Neto, N.B.; Saturnino, S.M.; Bomfim, D.C.; Custódio, C.C. Water stress induced by mannitol and sodium chloride in soybean cultivars. Braz. Arch. Biol. Technol. 2004, 47, 521–529. [Google Scholar] [CrossRef]
- Spadeto, C.; Lopes, J.C.; Mengarda, L.H.G.; Matheus, M.T.; Bernardes, P.M. Estresse salino e hídrico na germinação de sementes de garapa (Apuleia leiocarpa (VOGEL.) JF Macbr.). Enciclopédia Biosf. 2012, 8, 539–551. [Google Scholar]
- Ferreira, D.F. Sisvar: A Guide for its Bootstrap procedures in multiple comparisons. Ciência E Agrotecnologia 2014, 38, 109–112. [Google Scholar] [CrossRef]
- Soares, M.M.; Santos Junior, H.C.; Simões, M.G.; Pazzin, D.; Silva, L.J. Estresse hídrico e salino em sementes de soja classificadas em diferentes tamanhos. Pesqui. Agropecuária Trop. 2015, 45, 370–378. [Google Scholar] [CrossRef]


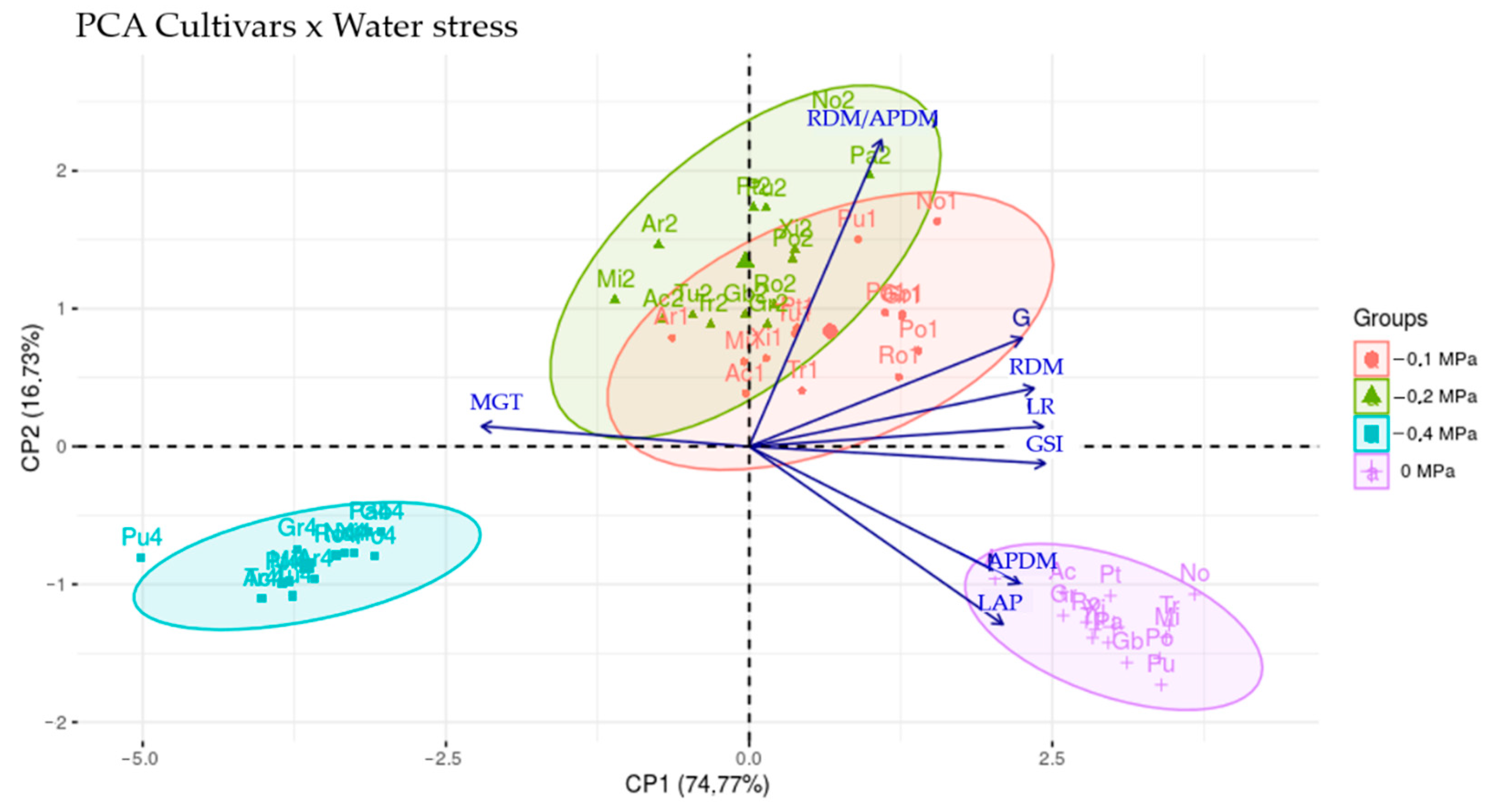{kind=link}
| Variable | ||||||||
|---|---|---|---|---|---|---|---|---|
| Variable | G | GSI | MGT | LR | LAP | RDM | APDM | RDM/ APDM |
| G | 1.00 | |||||||
| GSI | 0.93 ** | 1.00 | ||||||
| p-value | (<0.0001) | |||||||
| MGT | −0.83 ** | −0.93 ** | 1.00 | |||||
| p-value | (<0.0001) | (<0.0001) | ||||||
| LR | 0.83 ** | 0.88 ** | −0.89 ** | 1.00 | ||||
| p-value | (<0.0001) | (<0.0001) | (<0.0001) | |||||
| LAP | 0.87 ** | 0.94 ** | −0.87 ** | 0.86 ** | 1.00 | |||
| p-value | (<0.0001) | (<0.0001) | (<0.0001) | (<0.0001) | ||||
| RDM | 0.82 ** | 0.82 ** | −0.80 ** | 0.86 ** | 0.85 ** | 1.00 | ||
| p-value | (<0.0001) | (<0.0001) | (<0.0001) | (<0.0001) | (<0.0001) | |||
| APDM | 0.88 ** | 0.92 ** | −0.86 ** | 0.88 ** | 0.95 ** | 0.91 ** | 1.00 | |
| p-value | (<0.0001) | (<0.0001) | (<0.0001) | (<0.0001) | (<0.0001) | (<0.0001) | ||
| RDM/ APDM | 0.33 * | 0.25 | −0.28 * | 0.35 ** | 0.23 | 0.42 ** | 0.22 | 1.00 |
| p-value | (0.013) | (0.062) | (0.039) | (0.008) | (0.090) | (0.001) | (0.108) | |
| Variable | CP1 | CP2 | CP3 | CP4 | CP5 | CP6 | CP7 | CP8 |
|---|---|---|---|---|---|---|---|---|
| G | 0.366 | 0.270 | −0.012 | 0.688 | −0.212 | −0.085 | −0.466 | −0.225 |
| GSI | 0.397 | −0.042 | −0.157 | 0.401 | −0.060 | 0.105 | 0.735 | 0.321 |
| MGT | −0.359 | 0.050 | 0.829 | 0.358 | 0.164 | 0.011 | 0.149 | 0.065 |
| LR | 0.394 | 0.050 | 0.100 | −0.110 | 0.610 | −0.666 | −0.012 | 0.061 |
| LAP | 0.340 | −0.444 | 0.108 | 0.028 | 0.460 | 0.570 | −0.090 | −0.361 |
| RDM | 0.382 | 0.145 | 0.395 | −0.374 | −0.437 | −0.094 | 0.284 | −0.507 |
| APDM | 0.362 | −0.342 | 0.330 | −0.179 | −0.308 | 0.050 | −0.356 | 0.622 |
| RDM/ APDM | 0.177 | 0.765 | 0.070 | −0.230 | 0.234 | 0.449 | −0.068 | 0.254 |
| eigenvalues | 5.98 | 1.34 | 0.29 | 0.19 | 0.10 | 0.05 | 0.03 | 0.02 |
| EV (%) | 74.77 | 16.73 | 3.61 | 2.36 | 1.31 | 0.62 | 0.38 | 0.22 |
| AEV (%) | 74.77 | 91.50 | 95.11 | 97.47 | 98.77 | 99.39 | 99.78 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, L.K.B.d.; Silva, M.L.d.S.; Oliveira, A.B.d.; Amorim, A.V.; Dias, C.T.d.S.; Mesquita, R.O. Differential Tolerance of Cowpea Cultivars to Osmotic Stress in Germinative Phase: A Multivariate Approach. Biol. Life Sci. Forum 2022, 11, 48. https://doi.org/10.3390/IECPS2021-11994
Oliveira LKBd, Silva MLdS, Oliveira ABd, Amorim AV, Dias CTdS, Mesquita RO. Differential Tolerance of Cowpea Cultivars to Osmotic Stress in Germinative Phase: A Multivariate Approach. Biology and Life Sciences Forum. 2022; 11(1):48. https://doi.org/10.3390/IECPS2021-11994
Chicago/Turabian StyleOliveira, Letícia Kenia Bessa de, Maria Lílian dos Santos Silva, Alexandre Bosco de Oliveira, Aiala Vieira Amorim, Carlos Tadeu dos Santos Dias, and Rosilene Oliveira Mesquita. 2022. "Differential Tolerance of Cowpea Cultivars to Osmotic Stress in Germinative Phase: A Multivariate Approach" Biology and Life Sciences Forum 11, no. 1: 48. https://doi.org/10.3390/IECPS2021-11994