
Germination Performance of Different Sorghum Cultivars under Saline Conditions †

 ,
,
Abstract
:1. Introduction
2. Material and Methods
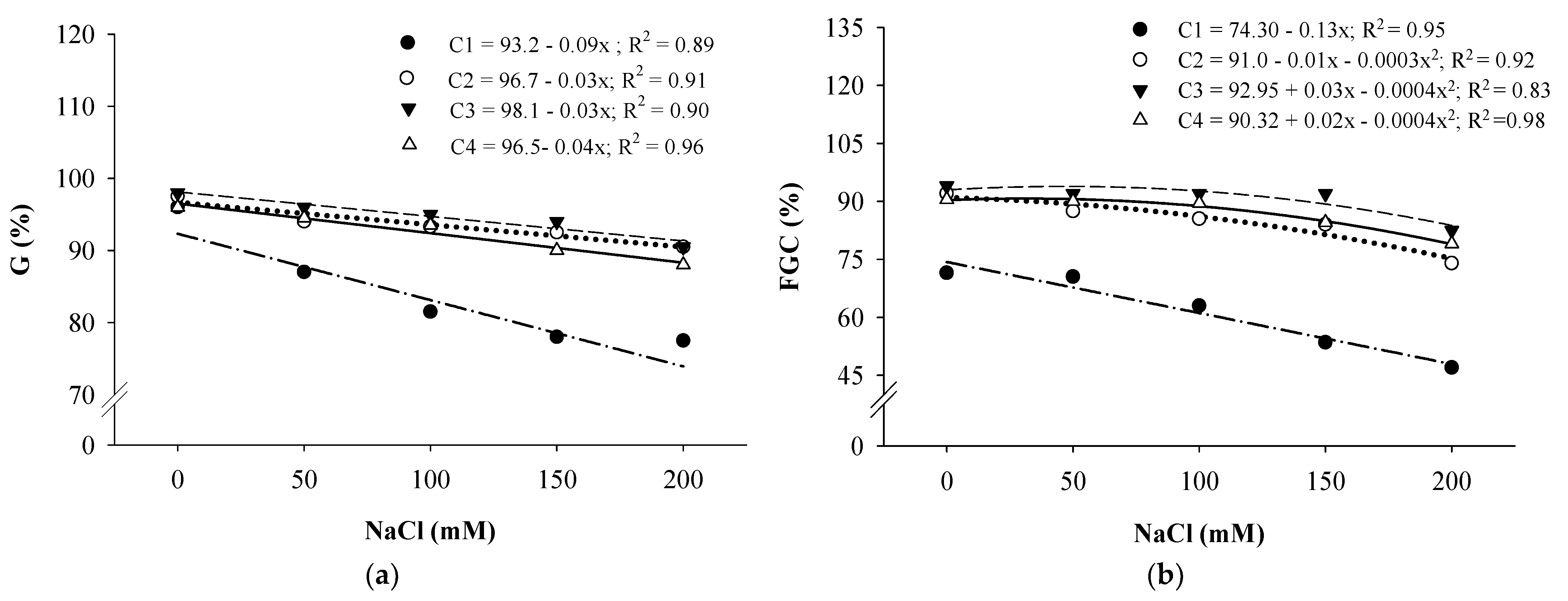
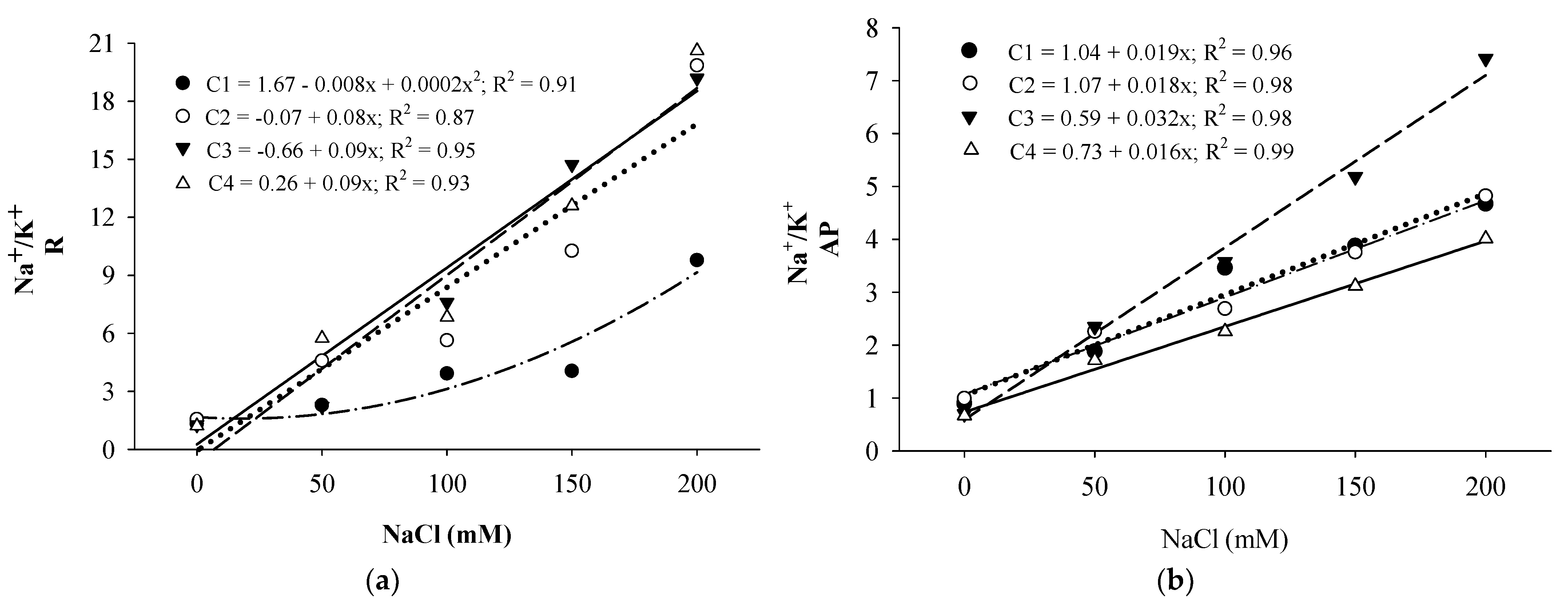
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant. Sci. 2017, 8, 1147–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrotti, A.; Chagas, R.M.; Ramos, V.C.; Prata, A.P.N.; Lucas, A.A.T.; Santos, P.B. Causas e consequên-cias do processo de salinização dos solos. Rev. Eletrônica Gestão Educ. Tecnol. Ambient. 2015, 19, 1308–1324. [Google Scholar]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Salvati, L.; Ferrara, C. The local-scale impact of soil salinization on the socioeconomic context: An exploratory analysis in Italy. Catena 2015, 127, 312–322. [Google Scholar] [CrossRef]
- Rasel, M.; Tahjib-Ul-Arif, M.; Hossain, M.A.; Hassan, L.; Farzana, S.; Brestic, M. Screening of Salt-Tolerant Rice Landraces by Seedling Stage Phenotyping and Dissecting Biochemical Determinants of Tolerance Mechanism. J. Plant Growth Regul. 2020, 40, 1853–1868. [Google Scholar] [CrossRef]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halo-phytes. Ann. Bot. 2015, 115, 419–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, D.S.; Simões, W.L.; Salviano, A.M.; De Souza, M.A.; Santos, J.E. Acúmulo e distribuição de nutrien-tes em genótipos de sorgo forrageiro sob salinidade. Braz. J. Maize Sorghum 2017, 16, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Feijão, A.R.; Silva, J.C.B.; Marques, E.C.; Prisco, J.T.; Gomes-Filho, E. Efeito da nutrição de nitrato na tolerância de plantas de sorgo sudão à salinidade. Rev. Ciência Agronômica 2011, 42, 675–683. [Google Scholar] [CrossRef]
- Lacerda, C.F.; Cambraia, J.; Oliva, M.A.; Ruiz, H.A. Changes in growth and in solute concentration in sorghum leaves and roots during salt stress recovery. Environ. Exp. Bot. 2005, 54, 69–76. [Google Scholar] [CrossRef]
- Larcher, W. Ecofisiologia Vegetal, 1st ed.; Rima: São Carlos, Brasil, 2000; p. 529. [Google Scholar]
- BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. In Regras para Análise de Sementes; Secretaria de Defesa Agropecuária/Mapa/ACS: Brasília, Brazil, 2009. [Google Scholar]
- Rosa, L.S.; Felippi, M.; Nogueira, A.C.; Grossi, F. Avaliação da ger-minação sob diferentes potenciais osmóticos e caracterização morfológica da semente e plântula de Ateleia glazioviana Baill (timbó). Cerne 2005, 11, 306–314. [Google Scholar]
- Maguire, J.D. Speed of germination-aid in selection and evaluation for seed-ling emergence and vigor. Crop. Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- Labouriau, L.G. A Germinação das Sementes, 24th ed.; OEA: Washington, DC, USA, 1983; p. 174.
- Rinne, K.T.; Saurer, M.; Streit, K.; Siegwolf, R.T.W. Evaluation of a liquid chromatography method for com-poundspecific d13C analysis of plant carbohydrates in alkaline media. Rapid Commun. Mass Spectrom. 2012, 26, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A language and environment for statistical computing. In The R Project for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Oliveira, A.B.; Gomes-Filho, E. Germinação e vigor de sementes de sorgo forrageiro sob estresse hídrico e salino. Rev. Bras. Sementes 2009, 31, 48–56. [Google Scholar] [CrossRef]
- Coelho, D.S.; Simões, W.L.; Mendes, A.; Dantas, B.F.; Rodrigues, J.A.; Souza, M.A.D. Germinação e crescimento inicial de variedades de sorgo forrageiro submetidas ao estresse salino. Rev. Bras. Eng. Agrícola E Ambient. 2014, 18, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: Adelaide, Australia, 2012; 651p. [Google Scholar]
- Sun, J.; Zou, D.T.; Luan, F.S.; Zhao, H.W.; Wang, J.G.; Liu, H.L.; Liu, Z.L. Dynamic QTL analysis of the Na+ content, K+ content, and Na+/ K+ ratio in rice roots during the feld growth under salt stress. Biol. Plant. 2014, 58, 689–696. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Yahyai, R.A.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509–528. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Kumar, S.; Saha, B.; Awasthi, J.; Dey, M.; Panda, S.K.; Sahoo, L. Crosstalk between salt, drought, and cold stress in plants: Toward genetic engineering for stress tolerance. In Abiotic Stress Response in Plants: Tuteja/Abiotic Stress Response in Plants; WileyVCH Verlag GmbH & Co., KGaA: Weinheim, Germany, 2016; pp. 55–86. [Google Scholar]
- Ma, Q.; Bao, A.; Chai, W.; Wang, W.; Zhang, J.; Li, Y.; Wang, S. Transcriptomic analysis of the succulent xerophyte Zygophyllum xanthoxylum in response to salt treatment and osmotic stress. Plant Soil 2016, 402, 343–361. [Google Scholar] [CrossRef]
- Aquino, A.J.S.; Lacerda, C.F.; Gomes-Filho, E.; Costa, R.N.T. Crescimento, partição de matéria seca e retenção de Na+, K+ e Cl− em dois genótipos de sorgo irrigados com águas salinas. Rev. Bras. Ciência Solo 2007, 31, 961–971. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sources of Variation | DF | Medium Square | |||||
|---|---|---|---|---|---|---|---|
| G | FGC | GSI | MGT | Na+/K+ R | Na+/K+ AP | ||
| Cultivares (C) | 3 | 481.65 ** | 3542.7 ** | 120.70 ** | 4.12 ** | 112.03 ** | 7.668 ** |
| Concentrações Salinas (CS) | 4 | 245.80 ** | 671.0 ** | 149.99 ** | 2.80 ** | 639.81 ** | 46.491 ** |
| Int. C × CS | 12 | 29.90 ** | 47.5 ** | 0.85 ** | 0.07 ** | 24.81 ** | 1.615 ** |
| Resíduo | 60 | 8.15 | 21.1 | 0.56 | 0.03 | 2.18 | 0.124 |
| Total | 79 | - | - | - | - | - | - |
| CV% | - | 3.13 | 5.68 | 4.85 | 4.99 | 19 | 11.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batista, A.B.P.; de Oliveira, L.K.B.; de Lima, K.V.G.; de Freitas, M.C.; Mesquita, R.O. Germination Performance of Different Sorghum Cultivars under Saline Conditions. Biol. Life Sci. Forum 2022, 11, 12. https://doi.org/10.3390/IECPS2021-11993
Batista ABP, de Oliveira LKB, de Lima KVG, de Freitas MC, Mesquita RO. Germination Performance of Different Sorghum Cultivars under Saline Conditions. Biology and Life Sciences Forum. 2022; 11(1):12. https://doi.org/10.3390/IECPS2021-11993
Chicago/Turabian StyleBatista, Ana Beatriz Pereira, Letícia Kenia Bessa de Oliveira, Késsia Vanessa Gomes de Lima, Matheus Carlos de Freitas, and Rosilene Oliveira Mesquita. 2022. "Germination Performance of Different Sorghum Cultivars under Saline Conditions" Biology and Life Sciences Forum 11, no. 1: 12. https://doi.org/10.3390/IECPS2021-11993