
Harmful Cyanobacterial Blooms: Going beyond the “Green” to Monitor and Predict HCBs


Department of Biology & CESAM (Centre for Environmental and Marine Studies), University of Aveiro, Campus Universitário de Santiago, 3810-193 Aveiro, Portugal
Hydrobiology 2024, 3(1), 11-30; https://doi.org/10.3390/hydrobiology3010002
Submission received: 9 November 2023
/
Revised: 14 December 2023
/
Accepted: 26 December 2023
/
Published: 2 January 2024
(This article belongs to the Special Issue Feature Papers in Hydrobiology 2022–2023)
{kind=link}
{kind=link}
Abstract
:Under the Climate Change scenario, the occurrence of Harmful Cyanobacterial Blooms (HCBs) is an increasingly concerning problem. Particularly for inland freshwaters, that have human populations depending on them for consumption or recreation, HCBs can lead to serious ecological damages and socio-economic impacts, but also to health risks for local communities. From satellite imagery to molecular data, there is an increasing number of methodological approaches that can help improve the monitoring and prediction of cyanobacterial blooms. However, although each methodology has its own strengths and limitations, generally there is a lack of data addressing specific and intraspecific information, which has implications for the modelling and prediction of the real dynamics and toxicity of HCBs. The present review intends to make a quick overview on current approaches to monitor cyanobacterial blooms and provide a tier-based integrative perspective for their application. A transversal monitoring at a wide scale should be enhanced but cannot rely only on pigment levels but rather include the specific and intraspecific diversity information that can be obtained from modern molecular tools. This is crucial to achieve the effective prediction, monitoring and management of HCBs under their increasing occurrence and severity trends in freshwaters.
1. Introduction
Cyanobacteria are ancient photoautotrophic bacteria, and their occurrence is ubiquitous throughout aquatic bodies, although they can also be found in terrestrial ecosystems or in symbiosis with other organisms, or even in extreme environments such as in hypersaline lakes, hot springs, polar regions, and deserts [1,2,3,4]. However, in freshwaters, the study of the cyanobacterial community has been the focus of particular interest due to the occurrence of Harmful Cyanobacterial Blooms (HCBs), resulting from the massive growth of cyanobacteria that are potentially toxic, with impacts on both aquatic and terrestrial organisms, leading to serious health impacts or even death [5,6,7,8,9,10,11,12,13,14,15,16]. Moreover, under a climate change scenario, the concern with HCBs is growing due to their spread and increased persistence worldwide [17,18,19,20,21,22], with major changes on the ecosystems [8,13,23,24] but also posing risks to human health [5,25,26] and leading to socio-economic impacts in local communities [18,27,28,29,30,31,32].
In fact, many bloom-forming cyanobacteria have been reported as toxic, with the production of a wide range of toxins generally classified into hepatotoxins, neurotoxins, cytotoxins and dermatotoxins/irritant toxins [26,33,34]. Nevertheless, the list of new cyanotoxins is still increasing [11,14,35,36]. The most reported cyanotoxins include: microcystins and nodularin (hepatotoxins); saxitoxins, anatoxins and β-methylamino-l-alanine (neurotoxins); cylindrospermopsin (cytotoxin); lyngbyatoxins and aplysiatoxins (dermatotoxins); and lipopolysaccharides (irritant toxins) [14,15,22,37,38,39,40]. For a long time, most attention has been given to microcystins, mainly due to its global spread occurrence and impacts on human health, either by acute or chronic exposure [5,39,41,42,43,44]. However, neurotoxic saxitoxins and anatoxin-a also pose significant health risks due to their rapid and acute poisonings [39,45,46] as well as the potential for chronic effects by extended exposure to low doses [47]. Cylindrospermopsin is another very frequent cyanotoxin [48,49], responsible for cytotoxic [26] and neurotoxic effects [50,51]. In fact, the proposed guideline value for long-term exposure to cylindrospermopsin in drinking water is 0.7 μg/L, which is even lower than the same guideline for microcystins (1.0 μg/L) [52]. Additionally, both hepatotoxins and neurotoxins can persist in water for long periods of time (from days up to several weeks) until its degradation [26,53], which increases the concern for extended exposure. The bioaccumulation of these toxins has also been addressed for aquatic organisms such as molluscs and fish [49,54,55,56,57] but also for crops irrigated with contaminated water [12,35,58,59,60], highlighting the risk range and potentially wider effects of cyanotoxins’ occurrence. The most common exposure routes to cyanotoxins include the consumption/drinking of contaminated water, but recreational exposure can also occur through the accidental water swallowing, skin contact or even inhalation of scum aerosols [33,45,61,62].
The most frequent cyanobacterial genera reported to have toxic strains include Microcystis, Anabaena/Dolichospermum, Aphanizomenon/Cuspidothrix/Sphaerospermopsis, Anabaenopsis, Cylindrospermopsis/Raphidiopsis, Oscillatoria/Planktothrix, Phormidium, Gloeotrichia, Lyngbya, Nodularia, Nostoc and Synechococcus [22,28,39,63,64,65,66,67,68,69,70,71,72,73]. Nonetheless, an increasing number of toxic bloom-forming species and strains has been recorded over the past two decades and it is predicted that HCBs can become more frequent and persistent under a climate change scenario, with the extension of warmer seasons [74,75,76,77,78,79], along with increased atmospheric CO2 levels and hydrological alterations due to extreme weather events such as floods or droughts [22,28,29,30,80]. The spread of invasive tropical bloom-forming species, including Raphidiopsis raciborskii, Cuspidothrix issatschenkoi and Sphaerospermopsis aphanizomenoides, has already been recorded at higher latitude regions across the globe [70,81,82,83,84,85,86,87,88,89] and with cyanotoxins’ production [90,91,92,93].
However, not all strains within harmful cyanobacterial species are toxic and species-specific and intraspecific information can thus have a crucial role for modelling and management programmes. Presently, there are numerous approaches aiming to monitor cyanobacterial blooms [7,14], but most of these are used independently, with major information limitations, and are still generally not considering detailed specific data on dominant cyanobacteria as essential to better understand the ecological dynamics of HCBs.
2. Impact of Intraspecific Variability on Blooms’ Ecology and Toxicity
2.1. Classical Taxonomy vs. Phylogenetic Approaches
Most historical records and ecological studies concerning cyanobacterial blooms have been based on the identification of bloom-forming cyanobacteria using classical taxonomy mainly sustained by phenotypic features, which can be tricky due to several reasons, namely the diversity of strains/ecotypes [94]. Moreover, with the rise of DNA-based molecular approaches (using mostly 16S rRNA gene sequences), the polyphyletic nature of some genera has become controversial [65,95,96,97] and several taxonomical revisions have been made for several species and even genera [94,98,99,100,101,102,103], using polyphasic approaches to integrate morphological, biochemical and molecular information [99,101,103]. This re-classification into new genera and species also amplified the confusion regarding the taxonomy of blooming cyanobacteria and their correct identification, particularly for Nostocaceae genera such as Anabaena and Aphanizomenon, which were divided into new genera including Dolichospermum, Chrysosporum, Cuspidothrix or Sphaerospermopsis [97,103,104,105]. For instance, Sphaerospermopsis aphanizomenoides was formerly known as Aphanizomenon aphanizomenoides (Forti) Horecká and Komárek and Anabaena aphanizomenoides Forti [98,99,103,106]. Other examples can be Raphidiopsis raciborskii and Cuspidothrix issatschenkoi, which were formerly known as Cylindrospermopsis raciborskii (Woloszynska) Seenayya and Subba Raju and Aphanizomenon issatschenkoi (Usačev) Proshkina–Lavrenko, respectively [107,108]. Therefore, many concerns have been raised related to the probability of misidentifications only relying on morphological characters under a light microscope, particularly when heterocysts and akinetes are not visible for nostocacean cyanobacteria [88,109,110,111,112,113]. In fact, although necessary, the controversy over the past decades on cyanobacterial taxonomy regarding Nostocales order, in particular, has also increased the confusion for a proper identification [65,95,96,97,99,101,103,114,115]. This can have major implications for the previous knowledge regarding the distribution and dynamics of species-specific blooms and their ecology and toxicity [63]. Therefore, polyphasic approaches are crucial for the identification and characterization of isolated cyanobacterial strains [101,116], allowing ulterior DNA sequence match and identification through phylogenetic affiliation, along with ecological and toxicological features [113,117,118]. This is the basis for a deeper exploration of global distribution patterns of cyanobacterial species by comparing strains from diverse geographical origins or from time-dispersed blooms [105,119].
2.2. Co-Dominance and Dynamics of Cyanobacterial Blooms
Cyanobacterial blooms are frequently co-dominated by several species (some of which can be very similar under a microscope and difficult to distinguish and identify correctly [88,120]). Moreover, it is known that growth requirements can vary greatly between different cyanobacterial species [19,22,52,121]. For instance, in spite of the relationship frequently reported between the occurrence of cyanobacterial blooms and eutrophication, blooms can occur under low levels of inorganic nitrogen and/or phosphorus, depending on the dominant species [22,75,113,122]. Then again, climate change conditions produce variations in the physiological responses of cyanobacterial cells and lead to important changes in the composition of bloom-dominating strains and, consequently, impact the whole dynamics and toxicity potential of a cyanobacterial bloom [30,123]. Nitrogen-fixing cyanobacteria, in particular, can take special advantage of warmer and CO2-rich conditions from climate change [123,124] and increase their geographic range of proliferation in the near future. In fact, as referred above, several Nostocales invasive diazotrophic and potentially toxic species (such as Raphidiopsis raciborskii, Cuspidothrix issatschenkoi and Sphaerospermopsis aphanizomenoides) have already been increasingly reported at higher latitudes [81,85,86,89,125,126,127]. On the other hand, during a bloom, the co-occurrence of different strains from a same species is also common, which makes it impossible to differentiate them exclusively using morphological features [88,118]. The phenotypic plasticity of some species (namely the invasive R. raciborskii and S. aphanizomenoides) can also correspond to intraspecific genetic heterogeneity even among strains from the same geographical origin [118,128], suggesting a microevolution that can provide these species an increasing expansion potential. These species-specific strains may diverse significantly in their ecological requirements and/or toxic potential [106,113,117,129,130,131], with implications not only for the bloom development and dynamics, but also for the toxicity outcomes and health risks, depending on the ratio of non-toxic vs. toxic strains. Therefore, the search for intraspecific information is becoming more and more important to understand local to global biogeographical, ecological and toxicity patterns. Here, DNA-based molecular methodologies play a crucial role not only by helping to overcome misidentifications based on morphological features but also by providing strain-specific data to track variations of cyanobacterial strains from geographically different regions [82,105,132,133] or re-incidences of blooming strains over time in the same water body [88,118,134,135].
2.3. Intraspecific Cyanotoxin Production Potential
As discussed above, the taxonomic confusion and controversy has also had an impact on the characterization of which cyanotoxins can be produced by each cyanobacterial species [65]. Additionally, each species can have toxic and non-toxic genotypes [69,136,137,138], which brings even more entropy into toxicological studies. The synthesis of different cyanotoxins relies on the presence of specific gene clusters [139,140,141], and some cyanobacteria can also have biosynthesis gene clusters for more than one cyanotoxin [11], meaning that cyanobacterial blooms can be also co-dominated by multiple-toxin-producing strains, as previously reported [142,143]. Additionally, the regulation of cyanotoxins production varies at the strain level and can be affected by environmental factors such as temperature, light, pH and concentration of nutrients such as iron and nitrogen [139] as well as the bacteria present in water [144,145]. For instance, increased water temperature can influence the toxic potential of a bloom by promoting a shift of dominance from non-toxic to toxic cyanobacterial strains; at the same time, it can also increase toxin expression in the toxic strains [136,146]. All these genetic and physiological variations at the species and strain levels have implications on the overall toxicity of a bloom and toxicity assessment for a particular species [147]. This highlights the need for more studies using polyphasic approaches at the strain level combining at least a molecular characterization along with geographical distribution, as the genotype differentiation in specific niches can be also attributed to a selection from environmental factors [69].
3. Current Monitoring and Assessment Tools for Cyanobacterial Blooms
3.1. Methodologies Not Considering the Diversity of Cyanobacteria Nor Cyanotoxicity
3.1.1. Remote Sensing
The revolution of spatial exploration and satellites has brought an important possibility to provide a monitoring approach at a wide geographical and temporal range for cyanobacterial blooms, using indexes that deliver estimates for photosynthetic pigments such as chlorophyll a (Chl-a) and phycocyanin (PC) [148,149]. As PC is a cyanobacteria-specific pigment (despite its punctual presence in some microalgae such as cryptophytes [150]), this information should provide evidence on the occurrence of cyanobacteria-dominated blooms rather than blooms of microalgae, as previously depicted from estimates using only Chl-a (as this pigment is shared by other phytoplanktonic groups) in the beginning of the 2000s [151]. In fact, recent algorithms are further trying to reduce the interference of Chl-a on the estimation of PC [152,153]. Despite the limitations [154], studies using remote sensing imagery for PC estimations have been hugely increasing over the past decade making use of the opportunity to retrieve more directly cyanobacterial-related biomass information from: extensive water surface areas, including entire water bodies; remote and inaccessible regions; and historical data sets, in a time-saving and cost-effective way [151].
The most-used satellite data series have been provided by Landsat series, Moderate Resolution Imaging Spectroradiometer (MODIS), Medium Resolution Imaging Spectrometer (MERIS), and, more recently, Sentinel-3 Ocean Land Colour Instrument (OLCI). However, as satellite imagery is based on optical properties, there are limitations for the estimation of pigment concentrations [155,156,157]. Moreover, the use of different satellite source data for modelling is also risky due to the lack of data homogeneity as a consequence of the diversity of resolution and spectra from the different sensors, as well as the interferences from clouds, atmosphere and water reflectance [158]. For inland waters, Landsat satellite imagery provides valuable continuous time-series data sets to estimate PC and turbidity, already available for almost four decades and have been used to study cyanobacterial blooms at big lakes such as Lake Erie [159], Taihu Lake [160] or Lake Champlain [161], taking advantage of the fine resolution (about 30 m). However, under a rapid change into a cyanobacterial bloom development, a coarse revisit time of 16 days is not sufficient. There are also some limitations regarding the use of empirical models. On the other hand, MODIS, MERIS and OLCI imagery data, although possessing a wider range of spectral bands and a shorter revisit time (up to 1 day), have only more recent data (since 2000, 2003 and 2016, respectively) and are not adequate to monitor small waterbodies due to their low spatial resolution (about 300 m). There are also limitations in PC estimation for blooms of cyanobacteria in inland waters, namely under oligotrophic conditions or during blooms with other phytoplankton (where other pigments’ signatures overlap in the signature curves) [162,163,164]. More recently, the launch of Sentinel-2 MSI (Multiple Spectral Instrument) and Lansat-8 OLI (Operational Land Imager) has provided the acquisition of data that could be comparable and with higher spatial resolution (10–60 m and 30 m, respectively), which is more adequate to monitor medium to small lakes and reservoirs [165,166,167,168]. However, the design of their imagery sensors was not originally thought for aquatic applications and the lack of an orange band (~620 nm) has been challenging and has required the development of complex algorithms for PC estimation [169,170].
Nevertheless, despite all limitations, remote sensing PC estimation has proven to provide a valuable proxy to cyanobacterial biomass, which can be a powerful tool to be complemented by field data, namely concerning bloom aggregations in the surface water. Yet, by itself, it is not sufficient to take further conclusions on the dominant cyanobacteria or real toxic potential of a bloom.
3.1.2. In Situ Sensors and Automated Unmanned Vehicles (AUVs)
Fluorescence probes have proven to be the most suitable approach for wide real-time monitoring to detect the development of a cyanobacterial bloom [171]. In situ and portable devices are now commercially available [32] to fulfil the primary screening and early alert for any bloom formation through the real-time monitoring of pigments such as Chl-a and PC, allowing to differentiate the occurrence of microalgal and cyanobacterial blooms through submersible fluorometers or laboratory spectrofluorometers. However, in situ phycocyanin fluorescence probes can also have some interferences for a correct measurement of PC due to factors such as natural organic matter, water temperature or natural light [172]. Another issue to consider is, once again, the species-specific variation, resulting in different PC concentrations due to disparities in cell PC content and cyanobacterial cell biovolume among the different bloom-forming species [172,173]. This hinders a correct definition of general thresholds for PC concentration. The spatial range of in situ sensors is also limited but the outcomes are worth it for their routine use instead of the pigment extraction procedures.
The interference from clouds, atmosphere and land reflectance at the nearshore zones when using remote sensing can also be surpassed by using imagery from automated unmanned vehicles (AUVs) including drones, boats, underwater vehicles and water samplers that can provide information from spatial or vertical depth profiles, hotspots, nearshore zones or critical health risk areas during the bloom development [174,175,176,177] and help calibrate and validate the algorithms for PC estimation from remote imagery data in lakes and reservoirs [170]. Additionally, data from low orbiting satellites such as CubeSats [178] can also be an interesting alternative. Despite the limitations previously recorded for samples dominated by filamentous bloom-forming strains [179], flow cytometry has also been developed to study the phytoplankton community structure and abundance as well as viability and metabolic activity of cyanobacteria [180]. This makes submersible flow cytometers also a valuable source of real-time data, and a semi-automated classification of cyanobacteria by devices such as FlowCAM or Imaging FlowCytobot, despite its limitations to distinguish between species [181,182], can develop as a promising tool for routine real-time monitoring of bloom-forming cyanobacteria in water bodies.
Information from environmental parameters and nutrients is also crucial to understand the drivers behind the variation of community structure and toxic potential of a bloom at the strain level, as discussed above. A huge variety of affordable portable sensors are now also accessible for real-time quantification of environmental parameters such as temperature, conductivity, pH and dissolved oxygen. More accessible and portable fluorometry equipment are likewise available and can also be used for citizen science programmes without needing a high degree of expertise to use it, extending the range of source data in small lakes and reservoirs for which remote sensing methodologies are not yet optimised.
3.1.3. Turbidity and Visual Inspection—The Contribution from Citizen Science
The change in water transparency and colour (including the potential formation of scum) during cyanobacterial blooms is visible and easily recordable through an image capture (e.g., BloomWatch and Bloomin’ Algae apps, created by U.S. Environmental Protection Agency and the U.K. Centre for Ecology and Hydrology, respectively) or by using simple methodologies. In fact, affordable turbidity sensors or Secchi-disk depth approaches have been widely used for water quality assessment even through citizen science without the need for deep expertise [183,184,185]. This information has been used as an additional data source for monitoring the occurrence of cyanobacterial blooms [186]. Of course, depending on the blooming species, there will be a diverse colour palette as well as a different aspect at the surface water, as not all species form the characteristic scum during blooms and there is no guarantee that the bloom is from cyanobacteria without further analyses. For instance, strains belonging to Microcystis, Dolichospermum, Cuspidothrix or Aphanizomenon usually form surface blooms with visible streaks or even scum at the water surface, but for genera such as Plantothrix or Raphidiopsis, blooms can also occur spread across the water column and the water does not seem to have an occurring bloom taking place which can cause increased risks [14,52].
Nevertheless, despite these limitations from visual inspection and turbidity in water bodies and the potential errors/bias of citizen science-based approaches, the originated data may be of great importance as a first “real-time” screening for cyanobacterial blooms’ development and should be promoted, particularly for small lakes and reservoirs, where remote sensing is not yet suitable to use, also increasing the awareness of citizens for this global societal problematic.
3.2. Methodologies Considering the Diversity of Cyanobacteria or Cyanotoxins
3.2.1. Microscopic Identification and Cell Counting
The use of microscopy is the traditional analysis to identify and count cyanobacteria in water samples, namely during cyanobacterial blooming events. Cell counts are usually made using sedimentation chambers in an inverted light microscope [14] but can also make use of epifluorescence by staining samples with selective dyes to distinguish autotrophic from heterotrophic cells [187]. However, these microscopy techniques are limited to a time and spatial point, requiring highly skilled personnel with taxonomical expertise and, by using light microscopy, there are major limitations to identify and count small cyanobacteria as well as dense spherical colonies (such as in most Microcystis spp.). This can lead to bias in total cyanobacterial cell and biovolume estimations which are parameters used by the WHO to recommend an Alert Levels Frameworks for drinking and recreational waters, indicating the potential for toxic risks [14]. Moreover, misidentifications are frequently detected, either due to the numerous recent taxonomical revisions or the limitations of morphological identification under a microscope. In particular, the high controversy on the taxonomy of Anabaena/Aphanizomenon-like cyanobacteria has a long background but modern cyanobacterial taxonomy based on polyphasic approaches have brought consensus and are providing a solid basis for the correct identification of Nostocales cyanobacteria [95,98,99,106,188]. Then again, there is the limitation that microscopy observation techniques do not allow the differentiation between toxic and non-toxic strains of a same species using morphological features.
Over the past three decades, there have also been some proposed methods for digital image recognition of cyanobacteria [189,190], but even with the recent technological innovation of digital image processing and machine learning, there is still much to develop until it can be a feasible tool to correctly recognise and identify cyanobacteria up to the species level in complex environmental mixtures [191,192,193,194]. The set of taxa used for training, calibration and validation of the models are still very limited which hinders the potential for its effective application in environmental water samples. Nevertheless, this kind of automatised methodologies using artificial intelligence (AI) will surely be the basis of future monitoring for HCBs by simultaneously integrating and processing data from diverse sources and correcting it according to the historical occurrence data from the particular water body geographical origin, highlighting the importance of creating local databases for cyanobacterial blooms.
3.2.2. DNA-Based Molecular Approaches
Several community profiling techniques based on DNA have been used over the past decades. Methodologies such as Denaturing Gradient Gel Electrophoresis (DGGE) and Fluorescent In Situ Hybridization (FISH) allowed to target and study the diversity of cyanobacterial communities directly from environmental samples without the need for cyanobacteria isolation and culture in the laboratory [195,196]. DGGE, in particular, was widely used to study variations in bacterial–cyanobacterial communities through space and time [56,197,198,199,200,201,202]. However, limitations mainly linked to effort and time consumption to obtain only a limited number of sequences have reduced its use as High-Throughput Sequencing (HTS) techniques became less expensive and more accessible, providing a huge amount of data to explore. This rapid development of sequencing analyses has allowed metagenomic approaches to bring a new perspective into cyanobacterial blooms dynamics and ecology by studying whole cyanobacterial communities directly from environmental samples and providing a huge amount of sequencing data, namely supplementary information on non-cultivable and/or low-density cyanobacteria. HTS analysis has helped to unravel the diversity within a cyanobacterial bloom, using 16S rRNA gene or 16S rRNA-23S rRNA ITS (internal transcribed spacer) sequences, directly from environmental samples, providing an overview of spatial and/or temporal variations of cyanobacterial bloom-forming phylotypes in space and/or time [88,134,135,203,204,205], but also cyanotoxin assessment and dispersal [63,66,92,206]. Moreover, HTS can simultaneously provide information on the overall microbial communities during the occurrence of cyanobacterial blooms and help explore their role in the blooms’ dynamics [88,135,207,208]. Depending on the dominant cyanobacterial species/strain during a bloom, significant variations can occur at the level of bacterial community composition, with reciprocal impacts on the bloom dynamics and toxicity, as some bacteria can promote/inhibit growth [209,210] whereas others can lyse cyanobacteria [211] and/or degrade cyanotoxins [144,145]. Therefore, this complementary information could also have major implications for subsequent modelling and water management strategies.
Nevertheless, despite all the advantages of studying the overall cyanobacterial community directly from environmental samples, it is crucial to increase the isolation and characterisation of dominating strains from cyanobacterial blooms in order to explore the intraspecific genetic variation of blooming species as well as their potential differences in physiology and toxicity, contributing to the creation of local database sets for each target water body. Several techniques have been used to genotypically distinguish strains from the same species, from fingerprinting to sequencing of phylogenetically relevant DNA fragments [82,105,118,132,133]. Once the different strains are characterised, their re-incidence would become easier to detect, even when using molecular approaches targeting the whole cyanobacterial community (such as HTS). In fact, over the past decade, a wide variety of studies using omic approaches (from genomics to metabolomics) has provided the establishment of increasingly robust databases which did not exist in the beginning of the omic analyses for cyanobacteria (about 25 years ago) due to the lack of species-specific data. The number of deposited genomes from bloom-forming cyanobacteria has increased, regarding genera such as Microcystis, Cylindrospermopsis/Raphidiopsis, Aphanizomenon, Dolichospermum/Anabaena, Lyngbya, Nostoc, Nodularia and Oscillatoria/Planktothrix [62,212]. This expansion in genomic information is not only in quantity but also in reviews and annotation, allowing an increasing robustness of genetic databases for the comparative analysis of sequencing results. Although these molecular methods are not yet suitable for a routine monitoring basis, as the procedures require time, expertise and demanding laboratory procedures besides the high cost for regular analyses that correspond to a one time and space sample each, they provide crucial information that should be considered for future modelling and management approaches.
3.2.3. Detection and Quantification of Cyanotoxins
Presently, cost-effective molecular approaches based on PCR and qPCR (quantitative Polymerase Chain Reaction) to detect genes from the operons involved in the biosynthesis of cyanotoxins can provide preliminary information on the toxic potential of a bloom [213,214]. Moreover, there are simple qualitative cyanotoxins assays and qPCR portable devices that can be a valuable screening tool for water managers or can be even used in citizen science projects. Other more complex methodologies such as FISH have also been used to detect specific toxic cyanobacteria targeting ribosomal sequences [215] or cyanotoxins synthesis-related genes [213,216,217,218]. HTS has also been applied to match genes of cyanotoxins to the corresponding toxigenic cyanobacterial taxa present in environmental samples [219]. However, the presence of a gene cluster does not mean the cyanotoxin is actually being synthesised which hinders to conclude about the actual toxic risk of a cyanobacterial bloom.
Therefore, biochemical detection techniques such as Enzyme-Linked Immunosorbent Assay (ELISA) or Protein Phosphatase Inhibition Assay (PPIA), and analytical methods including High-Performance Liquid Chromatography—Ultraviolet (HPLC-UV) or Liquid Chromatography—Mass Spectrometry (LC-MS) are still the most used methodologies for cyanotoxins quantification. However, these have advantages (namely the quantification of cyanotoxins), but also limitations mainly regarding the costs, time-consumption, and technical expertise [32]. Another issue to consider is that these methodologies analyse only one cyanotoxin at a time. On the other hand, in spite their importance to detect simultaneously the presence cyanotoxins and their effects, bioassays have been less used to the ethical issues associated with its wide application, and new methods have been developed such as aptamer-biosensors [220] although these are still not much used.
Although the quantification of toxins can be considered the ultimate goal of the monitoring strategy for cyanobacterial blooms, in order to manage immediate health risks for the water users, it is crucial to understand under which conditions these cyanotoxins are being produced and by which cyanobacteria. This could lead to better prediction modelling and management but requires a tier-based and integrated approach with methodologies that also provide information at the specific and intraspecific levels, in order to also consider the prevalence of toxic over non-toxic genotypes. Once again, there is the need to isolate bloom-forming strains to characterize them using polyphasic approaches (with morphological, molecular, and toxic features) to allow future re-incidences and related toxic risks. More studies are needed regarding the dynamics of toxic vs. non-toxic strains within a same species.
4. Integrative Perspective for an Effective Monitoring and Modelling of HCBs
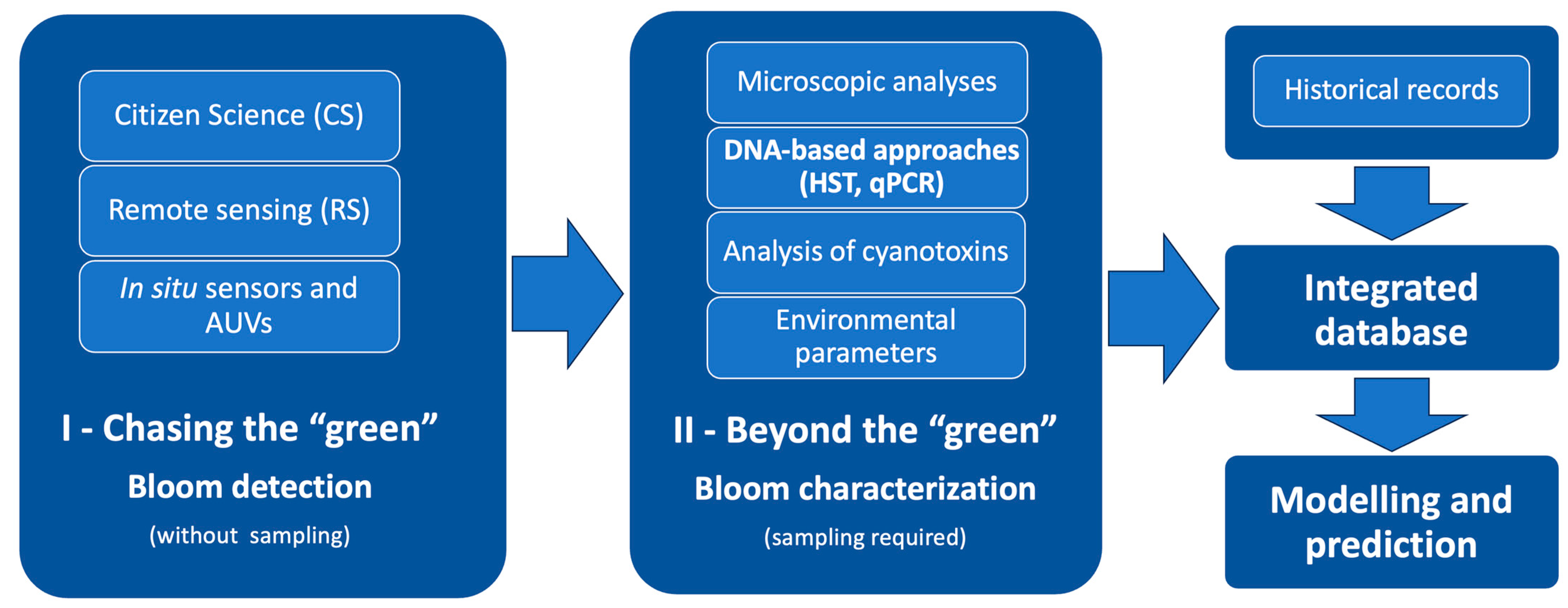
The perspective for the future monitoring and assessment of cyanobacterial blooms must be based on the integration of different data sources used for diverse but complementary monitoring purposes (Figure 1), allowing the development of robust predictive models based not only on metadata but also on scattered information, namely from historical records. The recovery, compilation, and integration of historical data in modelling is crucial to go further in the completion of this “puzzle”, even if it is dispersed in time and space. However, caution must be considered regarding nomenclature issues when dealing with historical data, in particular. Classical microscopic identification based only on morphological features can be hindered by factors such as the similarity between some taxa and the numerous taxonomical reviews over the past two decades, which may have led to misidentifications of cyanobacterial organisms or nomenclature mismatches, compromising a wider ecological and toxicological research on species-specific cyanobacterial blooms dynamics. Presently, DNA-based molecular methodologies play here a crucial role not only by helping to overcome misidentifications based on morphological features but also by providing strain-specific data that could be used in blooms’ prediction models and water management programmes. HTS analysis, in particular, as it becomes more affordable, should be considered for routine monitoring programmes, with the advantage of providing quantitative information and tracking re-incidence of bloom-forming cyanobacterial strains directly from water samples, minimizing constrains of culture-dependent approaches and classical ambiguous microscopic identifications. Furthermore, it should be raised the possibility of creating thresholds under the Alert Levels Frameworks for these molecular data (based on relative percentages of cyanobacterial OTUs recovered from HTS analysis through an equation where PC concentration should be considered as a proxy for total cyanobacterial biomass).
As discussed in this review, the toxic potential of an HCB depends on intraspecific genetic and physiological features of dominant cyanobacteria but also on environmental factors that impact the dominance of some strains over the others, as well as gene expression and toxin synthesis. Therefore, it is essential to crossover information of cyanotoxins occurrence with the producing cyanobacterial species/strains (as there are toxic and non-toxic genotypes) in order to go further in the understanding of HCBs and also the environmental context of their occurrence. Monitoring must aim not only to report the presence of cyanobacterial blooms but also to collect data to understand their occurrence and dynamics (see Figure 2).
The creation of global, regional, and local cross-referenced databases is also urgent, where multilevel source data could be easily assembled and made available for scientists, water managers, and civil society in a collaborative way.
As represented in Figure 2, the first level of information (phase I), concerning water colour and/or pigments (Chl a and PC) concentration estimation at a wide geographical scale and not depending on sampling and processing samples, can be provided by remote sensing but also by increasing citizen science programmes, where reliable information cannot be retrieved from satellite imagery. In fact, although remote sensing has provided an important basis for monitoring Harmful Algal Blooms (HABs) in the ocean and big lakes across the world, major drawbacks are encountered to monitor small reservoirs and lakes from space. However, these small freshwater bodies, mostly used for agriculture or recreational purposes, are also more prone to develop recurrent and severe cyanobacterial blooms (most of which dominated by toxic strains, leading to higher cyanotoxin levels), with immediate impacts for water users through skin contact, accidental swallowing of contaminated water, or inhalation of aerosols from scum, making urgent their monitoring on a daily or at least weekly basis. Therefore, at a regional scale, when alternative approaches based on local in situ fluorometric sensors or UAVs for a real-time monitoring are not available, the potential contribution of citizen science for “real-time” visual inspection or turbidity measurements should also be considered increasingly relevant at small water bodies (namely recreational urban lakes). Alternatively, whenever possible, particularly in large water bodies used for drinking water supply or recreation, in situ sensors and AUVs should complement the first level data by providing spatial information on the growing bloom distribution and pigments’ concentrations (Chl a and PC). In the presence of PC and Chl a levels near the lowest thresholds recommended by the WHO [14], action should be taken to phase II to further characterise the bloom through water sampling and the exploration of specific information on dominant cyanobacteria and toxic potential. In the lack of specialized operators for microscopic identification and cell counting, samples should be collected, filtered using 0.22 µm polycarbonate filters, and frozen (following a protocol using sterile conditions), for ulterior DNA extraction and analysis, namely through HTS (to search for cyanobacterial diversity) and qPCR (to search for cyanotoxin producers). Due to the cyanobacterial inter- and intra-specific genetic and physiological diversity, the massive promotion of molecular DNA-based approaches (such as HTS) must be the main direction in the near future to effectively approach and study HCBs dynamics. Following this strategy, the amount of information that could be cross-linked would tremendously increase the databases with more robust information that could better feed predictive models for HCBs.
Multi-tiered based approaches for monitoring HCBs have been proposed over the last years by integrating several methodologies with different purposes at different stages [32,221,222,223,224] and most modelling approaches are already integrating field sampling data with remote sensing imagery providing models considering hydrodynamics, wind/wave, sediment transport, and biogeochemistry. However, although Chl a and PC-based models and studies are essential, they will not help per se to unravel species-specific cyanobacterial blooms dynamics. Robust calibration and validation of the models with independent quality data are crucial and data from methodologies providing primary real-time information on blooms development must be combined with approaches that can accurately identify and provide quantitative information on dominant cyanobacteria (at least at the specific level, but preferably at the infra-specific level, using molecular data instead of microscopic analysis). Otherwise, the models become “blind”, no matter the complexity of algorithms and machine learning approaches, as the source data possess such a high degree of variability that could be drastically explained if using specific and intraspecific data. Multidisciplinary studies and models must therefore be promoted to integrate the different disciplines in a same approach. The development of models to understand HCBs distribution at a regional or even at a global scale must start incorporating data from diverse sources, from satellite imagery to dominant species/strains and cyanotoxins, along with environmental parameters and hydrological features, but all of these under an ecological perspective. Some models are also beginning to incorporate information at the cellular level (from transcriptomics, proteomics, and metabolomics) [212,225], which could help understanding why and how some cyanobacterial strains are dominant, but there is still a long way to go until an overall integration of data is feasible. As proposed in the model above and also discussed by Thompson et al. [226], a global programme for cyanobacterial monitoring should include the recommendation to collect and preserve samples for future genomic analysis when the conditions are not suitable at the moment, for several reasons. This can be a low time-consuming and cost-effective alternative to keep historical “freezed” information that can be analysed a posteriori if considered relevant but that does not become lost due to expertise and/or cost constraints.
On the other hand, automated methodologies using artificial intelligence (AI) will surely be the basis of future monitoring for HCBs and models will simultaneously integrate multisource data from diverse sources (remote sensing, in situ pigment sensors, UAVs, microscopy image recognition, molecular data from HTS and cyanotoxins, along with environmental parameters) and make real-time corrections according to the geographical historical database. Predictions will also become more realistic and consequently, the prevention of toxic cyanobacterial blooms outcomes will also be more effective. However, instead of the generalisation trend on the development of robust algorithms that can have a wide application throughout diverse water bodies across the globe to improve the output from remote sense data, maybe a new perspective is needed for the differentiation and variation arising from the ecological diversity of dominant cyanobacteria in blooms. The diversification of models is inevitable, as different blooming-species (and strains) can have major differences in ecological dynamics and toxicity. This is crucial to go deeper on the global ecology of HCBs and successfully understand, model, and predict their future occurrence.
Ultimately, national/regional collaborative programmes should be thought and evaluated, where diverse partners can provide different inputs, depending on their level of expertise (from scientific laboratories to water managers), but definitely not forgetting the involvement of the civil society (namely through citizen science approaches).
Funding
This work had the financial support from CESAM (UID/AMB/50017/2020), FCT/MCTES and FEDER (PT2020 Partnership Agreement and Compete 2020). FCT (Fundação para a Ciência e Tecnologia) funded a research contract for D.R.F. supported by national funds (OE), according to the DL 57/2016 (https://doi.org/10.54499/DL57/2016/CP1482/CT0034).
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Xu, H.-F.; Raanan, H.; Dai, G.-Z.; Oren, N.; Berkowicz, S.; Murik, O.; Kaplan, A.; Qiu, B.-S. Reading and Surviving the Harsh Conditions in Desert Biological Soil Crust: The Cyanobacterial Viewpoint. FEMS Microbiol. Rev. 2021, 45, fuab036. [Google Scholar] [CrossRef]
- Jasser, I.; Panou, M.; Khomutovska, N.; Sandzewicz, M.; Panteris, E.; Niyatbekov, T.; Łach, Ł.; Kwiatowski, J.; Kokociński, M.; Gkelis, S. Cyanobacteria in Hot Pursuit: Characterization of Cyanobacteria Strains, Including Novel Taxa, Isolated from Geothermal Habitats from Different Ecoregions of the World. Mol. Phylogenet. Evol. 2022, 170, 107454. [Google Scholar] [CrossRef] [PubMed]
- Perera, I.; Subashchandrabose, S.R.; Venkateswarlu, K.; Naidu, R.; Megharaj, M. Consortia of Cyanobacteria/Microalgae and Bacteria in Desert Soils: An Underexplored Microbiota. Appl. Microbiol. Biotechnol. 2018, 102, 7351–7363. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Pichel, F. Cyanobacteria. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 107–124. [Google Scholar]
- de Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.M.; Pereira, M.J. Microcystin-Producing Blooms—A Serious Global Public Health Issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Wood, R. Acute Animal and Human Poisonings from Cyanotoxin Exposure—A Review of the Literature. Environ. Int. 2016, 91, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Cotterill, V.; Hamilton, D.P.; Puddick, J.; Suren, A.; Wood, S.A. Phycocyanin Sensors as an Early Warning System for Cyanobacteria Blooms Concentrations: A Case Study in the Rotorua Lakes. N. Z. J. Mar. Freshw. Res. 2019, 53, 555–570. [Google Scholar] [CrossRef]
- Hilborn, E.D.; Beasley, V.R. One Health and Cyanobacteria in Freshwater Systems: Animal Illnesses and Deaths Are Sentinel Events for Human Health Risks. Toxins 2015, 7, 1374–1395. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Boyer, G.L. Health Impacts from Cyanobacteria Harmful Algae Blooms: Implications for the North American Great Lakes. Harmful Algae 2016, 54, 194–212. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Felpeto, A.B.; Latała, A. Allelopathic and Bloom-Forming Picocyanobacteria in a Changing World. Toxins 2018, 10, 48. [Google Scholar] [CrossRef]
- Janssen, E.M.L. Cyanobacterial Peptides beyond Microcystins—A Review on Co-Occurrence, Toxicity, and Challenges for Risk Assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef]
- Redouane, E.M.; El Amrani Zerrifi, S.; El Khalloufi, F.; Oufdou, K.; Oudra, B.; Lahrouni, M.; Campos, A.; Vasconcelos, V. Mode of Action and Fate of Microcystins in the Complex Soil-Plant Ecosystems. Chemosphere 2019, 225, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, V. Cyanobacteria Toxins: Diversity and Ecological Effects. Limnetica 2001, 20, 45–58. [Google Scholar] [CrossRef]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press: Boca Raton, FL, USA, 2021; ISBN 9788490225370. [Google Scholar]
- Cheung, M.Y.; Liang, S.; Lee, J. Toxin-Producing Cyanobacteria in Freshwater: A Review of the Problems, Impact on Drinking Water Safety, and Efforts for Protecting Public Health. J. Microbiol. 2013, 51, 1–10. [Google Scholar] [CrossRef]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful Freshwater Algal Blooms, with an Emphasis on Cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Barnard, M.A. Mitigating the Global Expansion of Harmful Cyanobacterial Blooms: Moving Targets in a Human- and Climatically-Altered World. Harmful Algae 2020, 96, 101845. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial Blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Duelling “CyanoHABs”: Unravelling the Environmental Drivers Controlling Dominance and Succession among Diazotrophic and Non-N2-Fixing Harmful Cyanobacteria. Environ. Microbiol. 2016, 18, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Havens, K.E.; Paerl, H.W. Climate Change at a Crossroad for Control of Harmful Algal Blooms. Environ. Sci. Technol. 2015, 49, 12605–12606. [Google Scholar] [CrossRef]
- Scholz, S.N.; Esterhuizen-Londt, M.; Pflugmacher, S. Rise of Toxic Cyanobacterial Blooms in Temperate Freshwater Lakes: Causes, Correlations and Possible Countermeasures. Toxicol. Environ. Chem. 2017, 99, 543–577. [Google Scholar] [CrossRef]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The Rise of Harmful Cyanobacteria Blooms: The Potential Roles of Eutrophication and Climate Change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Mehinto, A.C.; Smith, J.; Wenger, E.; Stanton, B.; Linville, R.; Brooks, B.W.; Sutula, M.A.; Howard, M.D.A. Synthesis of Ecotoxicological Studies on Cyanotoxins in Freshwater Habitats—Evaluating the Basis for Developing Thresholds Protective of Aquatic Life in the United States. Sci. Total Environ. 2021, 795, 148864. [Google Scholar] [CrossRef] [PubMed]
- Weralupitiya, C.; Wanigatunge, R.P.; Gunawardana, D.; Vithanage, M.; Magana-Arachchi, D. Cyanotoxins Uptake and Accumulation in Crops: Phytotoxicity and Implications on Human Health. Toxicon 2022, 211, 21–35. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial Toxins: Modes of Actions, Fate in Aquatic and Soil Ecosystems, Phytotoxicity and Bioaccumulation in Agricultural Crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ger, K.A.; Urrutia-Cordero, P.; Frost, P.C.; Hansson, L.A.; Sarnelle, O.; Wilson, A.E.; Lürling, M. The Interaction between Cyanobacteria and Zooplankton in a More Eutrophic World. Harmful Algae 2016, 54, 128–144. [Google Scholar] [CrossRef]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating Cyanobacterial Harmful Algal Blooms in Aquatic Ecosystems Impacted by Climate Change and Anthropogenic Nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef]
- Paerl, H.W. Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures. Toxins 2018, 10, 76. [Google Scholar] [CrossRef]
- Visser, P.M.; Verspagen, J.M.H.; Sandrini, G.; Stal, L.J.; Matthijs, H.C.P.; Davis, T.W.; Paerl, H.W.; Huisman, J. How Rising CO2 and Global Warming May Stimulate Harmful Cyanobacterial Blooms. Harmful Algae 2016, 54, 145–159. [Google Scholar] [CrossRef]
- Watson, S.B.; Miller, C.; Arhonditsis, G.; Boyer, G.L.; Carmichael, W.; Charlton, M.N.; Confesor, R.; Depew, D.C.; Höök, T.O.; Ludsin, S.A.; et al. The Re-Eutrophication of Lake Erie: Harmful Algal Blooms and Hypoxia. Harmful Algae 2016, 56, 44–66. [Google Scholar] [CrossRef]
- Bullerjahn, G.S.; McKay, R.M.; Davis, T.W.; Baker, D.B.; Boyer, G.L.; D’Anglada, L.V.; Doucette, G.J.; Ho, J.C.; Irwin, E.G.; Kling, C.L.; et al. Global Solutions to Regional Problems: Collecting Global Expertise to Address the Problem of Harmful Cyanobacterial Blooms. A Lake Erie Case Study. Harmful Algae 2016, 54, 223–238. [Google Scholar] [CrossRef]
- Stewart, I.; Webb, P.M.; Schluter, P.J.; Shaw, G.R. Recreational and Occupational Field Exposure to Freshwater Cyanobacteria—A Review of Anecdotal and Case Reports, Epidemiological Studies and the Challenges for Epidemiologic Assessment. Environ. Health 2006, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W. Cyanobacteria Secondary Metabolites—The Cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Manning, S.R.; Nobles, D.R. Impact of Global Warming on Water Toxicity: Cyanotoxins. Curr. Opin. Food Sci. 2017, 18, 14–20. [Google Scholar] [CrossRef]
- Huang, I.S.; Zimba, P.V. Cyanobacterial Bioactive Metabolites—A Review of Their Chemistry and Biology. Harmful Algae 2019, 83, 42–94. [Google Scholar] [CrossRef]
- Wiegand, C.; Pflugmacher, S. Ecotoxicological Effects of Selected Cyanobacterial Secondary Metabolites a Short Review. Toxicol. Appl. Pharmacol. 2005, 203, 201–218. [Google Scholar] [CrossRef]
- Smith, J.L.; Boyer, G.L.; Zimba, P. V A Review of Cyanobacterial Odorous and Bioactive Metabolites: Impacts and Management Alternatives in Aquaculture. Aquaculture 2008, 208, 5–20. [Google Scholar] [CrossRef]
- Svirčev, Z.; Lalić, D.; Bojadžija Savić, G.; Tokodi, N.; Drobac Backović, D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global Geographical and Historical Overview of Cyanotoxin Distribution and Cyanobacterial Poisonings; Springer: Berlin/Heidelberg, Germany, 2019; Volume 93, ISBN 0123456789. [Google Scholar]
- He, X.; Liu, Y.L.; Conklin, A.; Westrick, J.; Weavers, L.K.; Dionysiou, D.D.; Lenhart, J.J.; Mouser, P.J.; Szlag, D.; Walker, H.W. Toxic Cyanobacteria and Drinking Water: Impacts, Detection, and Treatment. Harmful Algae 2016, 54, 174–193. [Google Scholar] [CrossRef]
- Cao, L.; Massey, I.Y.; Feng, H.; Yang, F. A Review of Cardiovascular Toxicity of Microcystins. Toxins 2019, 11, 507. [Google Scholar] [CrossRef]
- Svirčev, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of Microcystins with Reference to Cases of Human Intoxications and Epidemiological Investigations of Exposures to Cyanobacteria and Cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef]
- Schreidah, C.M.; Ratnayake, K.; Senarath, K.; Karunarathne, A. Microcystins: Biogenesis, Toxicity, Analysis, and Control. Chem. Res. Toxicol. 2020, 33, 2225–2246. [Google Scholar] [CrossRef]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The Diversity of Cyanobacterial Toxins on Structural Characterization, Distribution and Identification: A Systematic Review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef] [PubMed]
- Koreiviene, J.; Anne, O.; Kasperovičiene, J.; Burškyte, V. Cyanotoxin Management and Human Health Risk Mitigation in Recreational Waters. Environ. Monit. Assess. 2014, 186, 4443–4459. [Google Scholar] [CrossRef] [PubMed]
- Testai, E.; Scardala, S.; Vichi, S.; Buratti, F.M.; Funari, E. Risk to Human Health Associated with the Environmental Occurrence of Cyanobacterial Neurotoxic Alkaloids Anatoxins and Saxitoxins. Crit. Rev. Toxicol. 2016, 46, 385–419. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, K.; Musgrave, I.F.; Humpage, A. Low Dose Extended Exposure to Saxitoxin and Its Potential Neurodevelopmental Effects: A Review. Environ. Toxicol. Pharmacol. 2016, 48, 7–16. [Google Scholar] [CrossRef]
- Moreira, C.; Azevedo, J.; Antunes, A.; Vasconcelos, V. Cylindrospermopsin: Occurrence, Methods of Detection and Toxicology. J. Appl. Microbiol. 2013, 114, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Scarlett, K.R.; Kim, S.; Lovin, L.M.; Chatterjee, S.; Scott, J.T.; Brooks, B.W. Global Scanning of Cylindrospermopsin: Critical Review and Analysis of Aquatic Occurrence, Bioaccumulation, Toxicity and Health Hazards. Sci. Total Environ. 2020, 738, 139807. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, M.G.; Prieto, A.I.; Gutiérrez-Praena, D.; Moreno, F.J.; Cameán, A.M.; Jos, A. Neurotoxic Assessment of Microcystin-LR, Cylindrospermopsin and Their Combination on the Human Neuroblastoma SH-SY5Y Cell Line. Chemosphere 2019, 224, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, M.G.; Gutiérrez-Praena, D.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Neurotoxicity Induced by Microcystins and Cylindrospermopsin: A Review. Sci. Total Environ. 2019, 668, 547–565. [Google Scholar] [CrossRef]
- Chorus, I.; Fastner, J.; Welker, M. Cyanobacteria and Cyanotoxins in a Changing Environment: Concepts, Controversies, Challenges. Water 2021, 13, 2463. [Google Scholar] [CrossRef]
- Edwards, C.; Graham, D.; Fowler, N.; Lawton, L.A. Biodegradation of Microcystins and Nodularin in Freshwaters. Chemosphere 2008, 73, 1315–1321. [Google Scholar] [CrossRef]
- Bi, X.; Dai, W.; Wang, X.; Dong, S.; Zhang, S.; Zhang, D.; Wu, M. Microcystins Distribution, Bioaccumulation, and Microcystis Genotype Succession in a Fish Culture Pond. Sci. Total Environ. 2019, 688, 380–388. [Google Scholar] [CrossRef]
- Onyango, D.M.; Orina, P.S.; Ramkat, R.C.; Kowenje, C.; Githukia, C.M.; Lusweti, D.; Lung’ayia, H.B.O. Review of Current State of Knowledge of Microcystin and Its Impacts on Fish in Lake Victoria. Lakes Reserv. 2020, 25, 350–361. [Google Scholar] [CrossRef]
- Vareli, K.; Zarali, E.; Zacharioudakis, G.S.A.; Vagenas, G.; Varelis, V.; Pilidis, G.; Briasoulis, E.; Sainis, I. Microcystin Producing Cyanobacterial Communities in Amvrakikos Gulf (Mediterranean Sea, NW Greece) and Toxin Accumulation in Mussels (Mytilus galloprovincialis). Harmful Algae 2012, 15, 109–118. [Google Scholar] [CrossRef]
- Preece, E.P.; Hardy, F.J.; Moore, B.C.; Bryan, M. A Review of Microcystin Detections in Estuarine and Marine Waters: Environmental Implications and Human Health Risk. Harmful Algae 2017, 61, 31–45. [Google Scholar] [CrossRef]
- Zhu, J.; Ren, X.; Liu, H.; Liang, C. Effect of Irrigation with Microcystins-Contaminated Water on Growth and Fruit Quality of Cucumis sativus L. and the Health Risk. Agric. Water Manag. 2018, 204, 91–99. [Google Scholar] [CrossRef]
- Romero-Oliva, C.S.; Contardo-Jara, V.; Block, T.; Pflugmacher, S. Accumulation of Microcystin Congeners in Different Aquatic Plants and Crops—A Case Study from Lake Amatitlán, Guatemala. Ecotoxicol. Environ. Saf. 2014, 102, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.L.; Utsumi, M. An Overview of the Accumulation of Microcystins in Aquatic Ecosystems. J. Environ. Manag. 2018, 213, 520–529. [Google Scholar] [CrossRef]
- Weirich, C.A.; Miller, T.R. Freshwater Harmful Algal Blooms: Toxins and Children’s Health. Curr. Probl. Pediatr. Adolesc. Health Care 2014, 44, 2–24. [Google Scholar] [CrossRef]
- Otten, T.G.; Paerl, H.W. Health Effects of Toxic Cyanobacteria in U.S. Drinking and Recreational Waters: Our Current Understanding and Proposed Direction. Curr. Environ. Health Rep. 2015, 2, 75–84. [Google Scholar] [CrossRef]
- Österholm, J.; Popin, R.V.; Fewer, D.P.; Sivonen, K. Phylogenomic Analysis of Secondary Metabolism in the Toxic Cyanobacterial Genera Anabaena, Dolichospermum and Aphanizomenon. Toxins 2020, 12, 248. [Google Scholar] [CrossRef]
- Jiang, Y.; Song, G.; Pan, Q.; Yang, Y.; Li, R. Identification of Genes for Anatoxin-a Biosynthesis in Cuspidothrix issatschenkoi. Harmful Algae 2015, 46, 43–48. [Google Scholar] [CrossRef]
- Cirés, S.; Ballot, A. A Review of the Phylogeny, Ecology and Toxin Production of Bloom-Forming Aphanizomenon spp. and Related Species within the Nostocales (Cyanobacteria). Harmful Algae 2016, 54, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Dreher, T.W.; Li, R. An Overview of Diversity, Occurrence, Genetics and Toxin Production of Bloom-Forming Dolichospermum (Anabaena) Species. Harmful Algae 2016, 54, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Puddick, J.; Fleming, R.; Heussner, A.H. Detection of Anatoxin-Producing Phormidium in a New Zealand Farm Pond and an Associated Dog Death. N. Z. J. Bot. 2017, 55, 36–46. [Google Scholar] [CrossRef]
- Chernova, E.; Sidelev, S.; Russkikh, I.; Voyakina, E.; Babanazarova, O.; Romanov, R.; Kotovshchikov, A.; Mazur-Marzec, H. Dolichospermum and Aphanizomenon as Neurotoxins Producers in Some Russian Freshwaters. Toxicon 2017, 130, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Capelli, C.; Ballot, A.; Cerasino, L.; Papini, A.; Salmaso, N. Biogeography of Bloom-Forming Microcystin Producing and Non-Toxigenic Populations of Dolichospermum lemmermannii (Cyanobacteria). Harmful Algae 2017, 67, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Antunes, J.T.; Leão, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: Review of the Distribution, Phylogeography, and Ecophysiology of a Global Invasive Species. Front. Microbiol. 2015, 6, 473. [Google Scholar] [CrossRef]
- Ballot, A.; Scherer, P.I.; Wood, S.A. Variability in the Anatoxin Gene Clusters of Cuspidothrix issatschenkoi from Germany, New Zealand, China and Japan. PLoS ONE 2018, 13, e0200774. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Bullerjahn, G.S.; McKay, R.M.L. The Complicated and Confusing Ecology of Microcystis Blooms. mBio 2020, 11, 1–5. [Google Scholar] [CrossRef]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing Organisms, Occurrence, Toxicity, Mechanism of Action and Human Health Toxicological Risk Evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- Deng, J.; Qin, B.; Paerl, H.W.; Zhang, Y.; Ma, J.; Chen, Y. Earlier and Warmer Springs Increase Cyanobacterial (Microcystis spp.) Blooms in Subtropical Lake Taihu, China. Freshw. Biol. 2014, 59, 1076–1085. [Google Scholar] [CrossRef]
- de Figueiredo, D.R.; Reboleira, A.S.S.P.; Antunes, S.C.; Abrantes, N.; Azeiteiro, U.; Gonçalves, F.; Pereira, M.J.; Gonçalves, F.; Figueiredo, D.R. The Effect of Environmental Parameters and Cyanobacterial Blooms on Phytoplankton Dynamics of a Portuguese Temperate Lake. Hydrobiologia 2006, 568, 145–157. [Google Scholar] [CrossRef]
- Wang, P.; Ma, J.; Wang, X.; Tan, Q. Rising Atmospheric CO2 Levels Result in an Earlier Cyanobacterial Bloom-Maintenance Phase with Higher Algal Biomass. Water Res. 2020, 185, 116267. [Google Scholar] [CrossRef] [PubMed]
- Vanderley, R.F.; Ger, K.A.; Becker, V.; Bezerra, M.G.T.A.; Panosso, R. Abiotic Factors Driving Cyanobacterial Biomass and Composition under Perennial Bloom Conditions in Tropical Latitudes. Hydrobiologia 2021, 848, 943–960. [Google Scholar] [CrossRef]
- Qin, B.; Deng, J.; Shi, K.; Wang, J.; Brookes, J.; Zhou, J.; Zhang, Y.; Zhu, G.; Paerl, H.W.; Wu, L. Extreme Climate Anomalies Enhancing Cyanobacterial Blooms in Eutrophic Lake Taihu, China. Water Resour. Res. 2021, 57, e2020WR029371. [Google Scholar] [CrossRef]
- Hayes, N.M.; Haig, H.A.; Simpson, G.L.; Leavitt, P.R. Effects of Lake Warming on the Seasonal Risk of Toxic Cyanobacteria Exposure. Limnol. Oceanogr. Lett. 2020, 5, 393–402. [Google Scholar] [CrossRef]
- Liu, X.; Lu, X.; Chen, Y. The Effects of Temperature and Nutrient Ratios on Microcystis Blooms in Lake Taihu, China: An 11-Year Investigation. Harmful Algae 2011, 10, 337–343. [Google Scholar] [CrossRef]
- Rzymski, P.; Brygider, A.; Kokociński, M. On the Occurrence and Toxicity of Cylindrospermopsis raciborskii in Poland. Limnol. Rev. 2017, 17, 23–29. [Google Scholar] [CrossRef]
- Moreira, C.; Fathalli, A.; Vasconcelos, V.; Antunes, A. Genetic Diversity and Structure of the Invasive Toxic Cyanobacterium Cylindrospermopsis raciborskii. Curr. Microbiol. 2011, 62, 1590–1595. [Google Scholar] [CrossRef]
- Kokociński, M.; Gagala, I.; Jasser, I.; Karosiene, J.; Kasperovičiene, J.; Kobos, J.; Koreiviene, J.; Soininen, J.; Szczurowska, A.; Woszczyk, M.; et al. Distribution of Invasive Cylindrospermopsis raciborskii in the East-Central Europe Is Driven by Climatic and Local Environmental Variables. FEMS Microbiol. Ecol. 2017, 93, fix035. [Google Scholar] [CrossRef]
- Kokocinski, M.; Dziga, D.; Spoof, L.; Stefaniak, K.; Jurczak, T.; Mankiewicz-Boczek, J.; Meriluoto, J. First Report of the Cyanobacterial Toxin Cylindrospermopsin in the Shallow, Eutrophic Lakes of Western Poland. Chemosphere 2009, 74, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Rücker, J.; Stüken, A.; Preußel, K.; Nixdorf, B.; Chorus, I.; Köhler, A.; Wiedner, C. Occurrence of the Cyanobacterial Toxin Cylindrospermopsin in Northeast Germany. Environ. Toxicol. 2007, 22, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Stüken, A.; Rücker, J.; Endrulat, T.; Preussel, K.; Hemm, M.; Nixdorf, B.; Karsten, U.; Wiedner, C. Distribution of Three Alien Cyanobacterial Species (Nostocales) in Northeast Germany: Cylindrospermopsis raciborskii, Anabaena bergii and Aphanizomenon aphanizomenoides. Phycologia 2006, 45, 696–703. [Google Scholar] [CrossRef]
- Kaštovský, J.; Hauer, T.; Mareš, J.; Krautová, M.; Bešta, T.; Komárek, J.; Desortová, B.; Heteša, J.; Hindáková, A.; Houk, V.; et al. A Review of the Alien and Expansive Species of Freshwater Cyanobacteria and Algae in the Czech Republic. Biol. Invasions 2010, 12, 3599–3625. [Google Scholar] [CrossRef]
- de Figueiredo, D.R.; Lopes, A.R.; Pereira, M.J.; Polónia, A.R.; Castro, B.B.; Gonçalves, F.; Gomes, N.C.M.; Cleary, D.F.R. Bacterioplankton Community Shifts during a Spring Bloom of Aphanizomenon gracile and Sphaerospermopsis aphanizomenoides at a Temperate Shallow Lake. Hydrobiology 2022, 1, 499–517. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Park, H.-K.; Kim, I.-S. Invasion and Toxin Production by Exotic Nostocalean cyanobacteria (Cuspidothrix, Cylindrospermopsis, and Sphaerospermopsis) in the Nakdong River, Korea. Harmful Algae 2020, 100, 101954. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.K.; Costa, L.D.F.; Yunes, J.S.; Resgalla, C.; Barufi, J.B.; de Oliveira Bastos, E.; Horta, P.A.; Rörig, L.R. Saxitoxins from the Freshwater Cyanobacterium Raphidiopsis raciborskii Can Contaminate Marine Mussels. Harmful Algae 2021, 103, 102004. [Google Scholar] [CrossRef]
- Tawong, W.; Pongcharoen, P.; Nishimura, T.; Adachi, M. Molecular Characterizations of Thai Raphidiopsis raciborskii (Nostocales, Cyanobacteria) Based on 16s rDNA, RbcLX, and Cylindrospermopsin Synthetase Genes. Plankton Benthos Res. 2019, 14, 211–223. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velázquez, D.; Casero, M.C.; Quesada, A. Phylogeography of Cylindrospermopsin and Paralytic Shellfish Toxin-Producing Nostocales Cyanobacteria from Mediterranean Europe (Spain). Appl. Environ. Microbiol. 2014, 80, 1359–1370. [Google Scholar] [CrossRef]
- Wood, S.A.; Rasmussen, J.P.; Holland, P.T.; Campbell, R.; Crowe, A.L.M. First Report of the Cyanotoxin Anatoxin-a from Aphanizomenon issatschenkoi (Cyanobacteria). J. Phycol. 2007, 43, 356–365. [Google Scholar] [CrossRef]
- Palinska, K.A.; Surosz, W. Taxonomy of Cyanobacteria: A Contribution to Consensus Approach. Hydrobiologia 2014, 740, 1–11. [Google Scholar] [CrossRef]
- Hindák, F. Morphological Variation of Four Planktic Nostocalean Cyanophytes—Members of the Genus Aphanizomenon or Anabaena? Hydrobiologia 2000, 438, 107–116. [Google Scholar] [CrossRef]
- Li, R.; Carmichael, W.W.; Liu, Y.; Watanabe, M.M. Taxonomic Re-Evaluation of Aphanizomenon flos-aquae NH-5 Based on Morphology and 16S rRNA Gene Sequences. Hydrobiologia 2000, 438, 99–105. [Google Scholar] [CrossRef]
- Komárek, J.; Zapomelová, E. Planktic Morphospecies of the Cyanobacterial Genus Anabaena = Subg. Dolichospermum—2. Part: Straight Types. Fottea 2008, 8, 1–14. [Google Scholar]
- Zapomělová, E.; Skácelová, O.; Pumann, P.; Kopp, R.; Janeček, E. Polyphasic Characterization of Three Strains of Anabaena reniformis and Aphanizomenon aphanizomenoides (Cyanobacteria) and Their Reclassification To Sphaerospermum gen. nov. (Incl. Anabaena kisseleviana) (45:1363–73). J. Phycol. 2010, 46, 415. [Google Scholar] [CrossRef] [PubMed]
- Zapomělová, E.; Hrouzek, P.; Řezanka, T.; Jezberová, J.; Řeháková, K.; Hisem, D.; Komárková, J. Polyphasic Characterization of Dolichospermum spp. and Sphaerospermopsis spp. (Nostocales, Cyanobacteria): Morphology, 16S rRNA Gene Sequences and Fatty Acid and Secondary Metabolite Profiles. J. Phycol. 2011, 47, 1152–1163. [Google Scholar] [CrossRef] [PubMed]
- Gama, W.A.; Rigonato, J.; Fiore, M.F.; Sant’Anna, C.L. New Insights into Chroococcus (Cyanobacteria) and Two Related Genera: Cryptococcum gen. nov. and Inacoccus gen. nov. Eur. J. Phycol. 2019, 54, 315–325. [Google Scholar] [CrossRef]
- Komárek, J. A Polyphasic Approach for the Taxonomy of Cyanobacteria: Principles and Applications. Eur. J. Phycol. 2016, 51, 346–353. [Google Scholar] [CrossRef]
- Komárková, J.; Jezberová, J.; Komárek, O.; Zapomělová, E. Variability of Chroococcus (Cyanobacteria) Morphospecies with Regard to Phylogenetic Relationships. Hydrobiologia 2010, 639, 69–83. [Google Scholar] [CrossRef]
- Zapomělová, E.; Jezberová, J.; Hrouzek, P.; Hisem, D.; Řeháková, K.; Komárková, J. Polyphasic Characterization of Three Strains of Anabaena reniformis and Aphanizomenon aphanizomenoides (Cyanobacteria) and Their Reclassification to Sphaerospermum gen. nov. (Incl. Anabaena kisseleviana). J. Phycol. 2009, 45, 1363–1373. [Google Scholar] [CrossRef]
- Komárek, J. The Cyanobacterial Genus Macrospermum. Fottea 2008, 8, 79–86. [Google Scholar] [CrossRef]
- Zapomělová, E.; Skácelová, O.; Pumann, P.; Kopp, R.; Janeček, E. Biogeographically Interesting Planktonic Nostocales (Cyanobacteria) in the Czech Republic and Their Polyphasic Evaluation Resulting in Taxonomic Revisions of Anabaena bergii Ostenfeld 1908 (Chrysosporum gen. nov.) and A. tenericaulis Nygaard 1949 (Dolichospermum tenericaule comb. nova). Hydrobiologia 2012, 698, 353–365. [Google Scholar] [CrossRef]
- Zapomělová, E.; Řeháková, K.; Jezberová, J.; Komárková, J. Polyphasic Characterization of Eight Planktonic Anabaena Strains (Cyanobacteria) with Reference to the Variability of 61 Anabaena Populations Observed in the Field. Hydrobiologia 2010, 639, 99–113. [Google Scholar] [CrossRef]
- Aguilera, A.; Gómez, E.B.; Kaštovský, J.; Echenique, R.O.; Salerno, G.L. The Polyphasic Analysis of Two Native Raphidiopsis Isolates Supports the Unification of the Genera Raphidiopsis and Cylindrospermopsis (Nostocales, Cyanobacteria). Phycologia 2018, 57, 130–146. [Google Scholar] [CrossRef]
- Rajaniemi, P.; Komárek, J.; Willame, R.; Hrouzek, P.; Kaštovská, K.; Hoffmann, L.; Sivonen, K. Taxonomic Consequences from the Combined Molecular and Phenotype Evaluation of Selected Anabaena and Aphanizomenon Strains. Algol. Stud. Arch. Für Hydrobiol. 2005, 117, 371–391. [Google Scholar] [CrossRef]
- Choi, H.J.; Joo, J.H.; Kim, J.H.; Wang, P.; Ki, J.S.; Han, M.S. Morphological Characterization and Molecular Phylogenetic Analysis of Dolichospermum hangangense (Nostocales, Cyanobacteria) sp. nov. from Han River, Korea. Algae 2018, 33, 143–156. [Google Scholar] [CrossRef]
- Johansen, J.R.; Kovacik, L.; Casamatta, D.A.; Fučiková, K.; Kaštovský, J. Utility of 16S-23S ITS Sequence and Secondary Structure for Recognition of Intrageneric and Intergeneric Limits within Cyanobacterial Taxa: Leptolyngbya corticola sp. nov. (Pseudanabaenaceae, Cyanobacteria). Nova Hedwig. 2011, 92, 283–302. [Google Scholar] [CrossRef]
- Moore, D.; McGregor, G.B.; Shaw, G. Morphological Changes during Akinete Germination in Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria). J. Phycol. 2004, 40, 1098–1105. [Google Scholar] [CrossRef]
- Ryu, H.S.; Shin, R.Y.; Lee, J.H. Morphology and Taxonomy of the Aphanizomenon spp. (Cyanophyceae) and Related Species in the Nakdong River, South Korea. J. Ecol. Environ. 2016, 41, 6. [Google Scholar] [CrossRef]
- de Figueiredo, D.R.; Gonçalves, A.M.M.; Castro, B.B.; Gonçalves, F.; Pereira, M.J.; Correia, A. Differential Inter-and Intra-Specific Responses of Aphanizomenon Strains to Nutrient Limitation and Algal Growth Inhibition. J. Plankton Res. 2011, 33, 1606–1616. [Google Scholar] [CrossRef]
- Komárek, J. Modern Taxonomic Revision of Planktic Nostocacean Cyanobacteria: A Short Review of Genera. Hydrobiologia 2010, 639, 231–243. [Google Scholar] [CrossRef]
- Komárek, J. Cyanobacterial Taxonomy: Current Problems and Prospects for the Integration of Traditional and Molecular Approaches. Algae 2006, 21, 349–375. [Google Scholar] [CrossRef]
- Komárek, J. Quo Vadis, Taxonomy of Cyanobacteria (2019). Fottea 2020, 20, 104–110. [Google Scholar] [CrossRef]
- Sanchis, D.; Carrasco, D.; Quesada, A. The Genus Microcystis (Microcystaceae/Cyanobacteria) from a Spanish Reservoir: A Contribution to the Definition of Morphological Variations. Nova Hedwig. 2004, 79, 479–495. [Google Scholar] [CrossRef]
- de Figueiredo, D.R.; Alves, A.; Pereira, M.J.; Correia, A. Molecular Characterization of Bloom-Forming Aphanizomenon Strains Isolated from Vela Lake (Western Central Portugal). J. Plankton Res. 2010, 32, 239–252. [Google Scholar] [CrossRef]
- Werner, V.R.; Laughinghouse, H.D., IV; Fiore, M.F.; Sant’anna, C.L.; Hoff, C.; de Souza Santos, K.R.; Neuhaus, E.B.; Molica, R.J.R.; Honda, R.Y.; Echenique, R.O.; et al. Morphological and Molecular Studies of Sphaerospermopsis torques-reginae (Cyanobacteria, Nostocales) from South American Water Blooms. Phycologia 2012, 51, 228–238. [Google Scholar] [CrossRef]
- Lv, X.; Cheng, Y.; Zhang, S.; Hu, Z.; Xiao, P.; Zhang, H.; Geng, R.; Li, R. Polyphasic Characterization and Taxonomic Evaluation of a Bloom-Forming Strain Morphologically Resembling Radiocystis fernandoi (Chroococcales, Cyanobacteria) from Lake Erhai, China. Diversity 2022, 14, 816. [Google Scholar] [CrossRef]
- Tanvir, R.U.; Hu, Z.; Zhang, Y.; Lu, J. Cyanobacterial Community Succession and Associated Cyanotoxin Production in Hypereutrophic and Eutrophic Freshwaters. Environ. Pollut. 2021, 290, 118056. [Google Scholar] [CrossRef]
- Gobler, C.J.; Burkholder, J.A.M.; Davis, T.W.; Harke, M.J.; Johengen, T.; Stow, C.A.; Van de Waal, D.B. The Dual Role of Nitrogen Supply in Controlling the Growth and Toxicity of Cyanobacterial Blooms. Harmful Algae 2016, 54, 87–97. [Google Scholar] [CrossRef]
- Ma, J.; Wang, P. Effects of Rising Atmospheric CO2 Levels on Physiological Response of Cyanobacteria and Cyanobacterial Bloom Development: A Review. Sci. Total Environ. 2021, 754, 141889. [Google Scholar] [CrossRef]
- Symes, E.; van Ogtrop, F.F. The Effect of Pre-Industrial and Predicted Atmospheric CO2 Concentrations on the Development of Diazotrophic and Non-Diazotrophic Cyanobacterium: Dolichospermum circinale and Microcystis aeruginosa. Harmful Algae 2019, 88, 101536. [Google Scholar] [CrossRef] [PubMed]
- Budzyńska, A.; Gołdyn, R. Domination of Invasive Nostocales (Cyanoprokaryota) at 52°N Latitude. Phycol. Res. 2017, 65, 322–332. [Google Scholar] [CrossRef]
- Budzyńska, A.; Rosińska, J.; Pełechata, A.; Toporowska, M.; Napiórkowska-Krzebietke, A.; Kozak, A.; Messyasz, B.; Pęczuła, W.; Kokociński, M.; Szeląg-Wasielewska, E.; et al. Environmental Factors Driving the Occurrence of the Invasive Cyanobacterium Sphaerospermopsis aphanizomenoides (Nostocales) in Temperate Lakes. Sci. Total Environ. 2019, 650, 1338–1347. [Google Scholar] [CrossRef] [PubMed]
- Kokociński, M.; Soininen, J. New Insights into the Distribution of Alien Cyanobacterium Chrysosporum bergii (Nostocales, Cyanobacteria). Phycol. Res. 2019, 67, 208–214. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, Y.; Li, R.; Yang, Y.; Jiang, Y. Recent Advances in the Ecology of Bloom-Forming Raphidiopsis (Cylindrospermopsis) raciborskii: Expansion in China, Intraspecific Heterogeneity and Critical Factors for Invasion. Int. J. Environ. Res. Public Health 2023, 20, 1984. [Google Scholar] [CrossRef] [PubMed]
- Galvanese, E.F.; Padial, A.A.; Aubriot, L. Acclimation at High Temperatures Increases the Ability of Raphidiopsis raciborskii (Cyanobacteria) to Withstand Phosphate Deficiency and Reveals Distinct Strain Responses. Eur. J. Phycol. 2019, 54, 359–368. [Google Scholar] [CrossRef]
- Joung, S.-H.; Oh, H.-M.; Ko, S.-R.; Ahn, C.-Y. Correlations between Environmental Factors and Toxic and Non-Toxic Microcystis Dynamics during Bloom in Daechung Reservoir, Korea. Harmful Algae 2011, 10, 188–193. [Google Scholar] [CrossRef]
- Yoshida, M.; Yoshida, T.; Satomi, M.; Takashima, Y.; Hosoda, N.; Hiroishi, S. Intra-Specific Phenotypic and Genotypic Variation in Toxic Cyanobacterial Microcystis Strains. J. Appl. Microbiol. 2008, 105, 407–415. [Google Scholar] [CrossRef]
- Haande, S.; Rohrlack, T.; Ballot, A.; Røberg, K.; Skulberg, R.; Beck, M.; Wiedner, C. Genetic Characterisation of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) Isolates from Africa and Europe. Harmful Algae 2008, 7, 692–701. [Google Scholar] [CrossRef]
- Gugger, M.; Molica, R.; Le Berre, B.; Dufour, P.; Bernard, C.; Humbert, J.-F. Genetic Diversity of Cylindrospermopsis Strains (Cyanobacteria) Isolated from Four Continents. Appl. Environ. Microbiol. 2005, 71, 1097–1100. [Google Scholar] [CrossRef]
- Hur, M.; Lee, I.; Tak, B.-M.; Lee, H.J.; Yu, J.J.; Cheon, S.U.; Kim, B.-S. Temporal Shifts in Cyanobacterial Communities at Different Sites on the Nakdong River in Korea. Water Res. 2013, 47, 6973–6982. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.W.; Farnsley, S.E.; LeCleir, G.R.; Layton, A.C.; Satchwell, M.F.; DeBruyn, J.M.; Boyer, G.L.; Zhu, G.; Paerl, H.W. The Relationships between Nutrients, Cyanobacterial Toxins and the Microbial Community in Taihu (Lake Tai), China. Harmful Algae 2011, 10, 207–215. [Google Scholar] [CrossRef]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The Effects of Temperature and Nutrients on the Growth and Dynamics of Toxic and Non-Toxic Strains of Microcystis during Cyanobacteria Blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Kurmayer, R.; Christiansen, G.; Fastner, J.; Börner, T. Abundance of Active and Inactive Microcystin Genotypes in Populations of the Toxic Cyanobacterium Planktothrix spp. Environ. Microbiol. 2004, 6, 831–841. [Google Scholar] [CrossRef]
- Johansson, E.; Legrand, C.; Björnerås, C.; Godhe, A.; Mazur-Marzec, H.; Säll, T.; Rengefors, K. High Diversity of Microcystin Chemotypes within a Summer Bloom of the Cyanobacterium Microcystis botrys. Toxins 2019, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The Genetics, Biosynthesis and Regulation of Toxic Specialized Metabolites of Cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a Synthetase Gene Cluster of the Cyanobacterium Anabaena sp. strain 37 and Molecular Methods to Detect Potential Producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [PubMed]
- Méjean, A.; Paci, G.; Gautier, V.; Ploux, O. Biosynthesis of Anatoxin-a and Analogues (Anatoxins) in Cyanobacteria. Toxicon 2014, 91, 15–22. [Google Scholar] [CrossRef]
- Burford, M.A.; Davis, T.W.; Orr, P.T.; Sinha, R.; Willis, A.; Neilan, B. Nutrient-Related Changes in the Toxicity of Field Blooms of the Cyanobacterium, Cylindrospermopsis raciborskii. FEMS Microbiol. Ecol. 2014, 89, 135–148. [Google Scholar] [CrossRef]
- Lei, L.; Lei, M.; Cheng, N.; Chen, Z.; Xiao, L.; Han, B.P.; Lin, Q. Nutrient Regulation of Relative Dominance of Cylindrospermopsin-Producing and Non-Cylindrospermopsin-Producing Raphidiopsis raciborskii. Front. Microbiol. 2021, 12, 793544. [Google Scholar] [CrossRef]
- Shimizu, K.; Maseda, H.; Okano, K.; Itayama, T.; Kawauchi, Y.; Chen, R.; Utsumi, M.; Zhang, Z.; Sugiura, N. How Microcystin-Degrading Bacteria Express Microcystin Degradation Activity. Lakes Reserv. 2011, 16, 169–178. [Google Scholar] [CrossRef]
- Kormas, K.A.; Lymperopoulou, D.S. Cyanobacterial Toxin Degrading Bacteria: Who Are They? Biomed. Res. Int. 2013, 2013, 1463894. [Google Scholar] [CrossRef] [PubMed]
- Dziallas, C.; Grossart, H.P. Increasing Oxygen Radicals and Water Temperature Select for Toxic Microcystis sp. PLoS ONE 2011, 6, e25569. [Google Scholar] [CrossRef] [PubMed]
- Omidi, A.; Esterhuizen-Londt, M.; Pflugmacher, S. Still Challenging: The Ecological Function of the Cyanobacterial Toxin Microcystin–What We Know so Far. Toxin Rev. 2018, 37, 87–105. [Google Scholar] [CrossRef]
- Ogashawara, I.; Mishra, D.; Mishra, S.; Curtarelli, M.; Stech, J. A Performance Review of Reflectance Based Algorithms for Predicting Phycocyanin Concentrations in Inland Waters. Remote Sens. 2013, 5, 4774–4798. [Google Scholar] [CrossRef]
- Borges, H.D.; Cicerelli, R.E.; De Almeida, T.; Roig, H.L.; Olivetti, D. Monitoring Cyanobacteria Occurrence in Freshwater Reservoirs Using Semi-Analytical Algorithms and Orbital Remote Sensing. Mar. Freshw. Res. 2020, 71, 569–578. [Google Scholar] [CrossRef]
- Greenwold, M.J.; Cunningham, B.R.; Lachenmyer, E.M.; Pullman, J.M.; Richardson, T.L.; Dudycha, J.L. Diversification of Light Capture Ability Was Accompanied by the Evolution of Phycobiliproteins in Cryptophyte Algae. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190655. [Google Scholar] [CrossRef]
- Ogashawara, I. Determination of Phycocyanin from Space—A Bibliometric Analysis. Remote Sens. 2020, 12, 567. [Google Scholar] [CrossRef]
- Mishra, S.; Mishra, D.R.; Lee, Z.; Tucker, C.S. Quantifying Cyanobacterial Phycocyanin Concentration in Turbid Productive Waters: A Quasi-Analytical Approach. Remote Sens. Environ. 2013, 133, 141–151. [Google Scholar] [CrossRef]
- Li, L.; Li, L.; Song, K. Remote Sensing of Freshwater Cyanobacteria: An Extended IOP Inversion Model of Inland Waters (IIMIW) for Partitioning Absorption Coefficient and Estimating Phycocyanin. Remote Sens. Environ. 2015, 157, 9–23. [Google Scholar] [CrossRef]
- Yan, Y.; Bao, Z.; Shao, J. Phycocyanin Concentration Retrieval in Inland Waters: A Comparative Review of the Remote Sensing Techniques and Algorithms. J. Great Lakes Res. 2018, 44, 748–755. [Google Scholar] [CrossRef]
- Witter, D.L.; Ortiz, J.D.; Palm, S.; Heath, R.T.; Budd, J.W. Assessing the Application of SeaWiFS Ocean Color Algorithms to Lake Erie. J. Great Lakes Res. 2009, 35, 361–370. [Google Scholar] [CrossRef]
- Pirasteh, S.; Mollaee, S.; Fatholahi, S.N.; Li, J. Estimation of Phytoplankton Chlorophyll-a Concentrations in the Western Basin of Lake Erie Using Sentinel-2 and Sentinel-3 Data|Estimation Des Concentrations de Chlorophylle-a Du Phytoplancton Dans Le Bassin Ouest Du Lac Érié à l’aide Des Données Sentin. Can. J. Remote Sens. 2020, 46, 585–602. [Google Scholar] [CrossRef]
- Binding, C.E.; Greenberg, T.A.; Bukata, R.P. The MERIS Maximum Chlorophyll Index; Its Merits and Limitations for Inland Water Algal Bloom Monitoring. J. Great Lakes Res. 2013, 39, 100–107. [Google Scholar] [CrossRef]
- Wynne, T.T.; Stumpf, R.P.; Tomlinson, M.C.; Fahnenstiel, G.L.; Dyble, J.; Schwab, D.J.; Joshi, S.J. Evolution of a Cyanobacterial Bloom Forecast System in Western Lake Erie: Development and Initial Evaluation. J. Great Lakes Res. 2013, 39, 90–99. [Google Scholar] [CrossRef]
- Vincent, R.K.; Qin, X.; McKay, R.M.L.; Miner, J.; Czajkowski, K.; Savino, J.; Bridgeman, T. Phycocyanin Detection from LANDSAT TM Data for Mapping Cyanobacterial Blooms in Lake Erie. Remote Sens. Environ. 2004, 89, 381–392. [Google Scholar] [CrossRef]
- Yin, Z.; Li, J.; Liu, Y.; Xie, Y.; Zhang, F.; Wang, S.; Sun, X.; Zhang, B. Water Clarity Changes in Lake Taihu over 36 Years Based on Landsat TM and OLI Observations. Int. J. Appl. Earth Obs. Geoinf. 2021, 102, 102457. [Google Scholar] [CrossRef]
- Isenstein, E.M.; Kim, D.; Park, M.H. Modeling for Multi-Temporal Cyanobacterial Bloom Dominance and Distributions Using Landsat Imagery. Ecol. Inform. 2020, 59, 101119. [Google Scholar] [CrossRef]
- Shi, K.; Zhang, Y.; Qin, B.; Zhou, B. Remote Sensing of Cyanobacterial Blooms in Inland Waters: Present Knowledge and Future Challenges. Sci. Bull. 2019, 64, 1540–1556. [Google Scholar] [CrossRef]
- Simis, S.G.H.; Peters, S.W.M.; Gons, H.J. Remote Sensing of the Cyanobacterial Pigment Phycocyanin in Turbid Inland Water. Limnol. Oceanogr. 2005, 50, 237–245. [Google Scholar] [CrossRef]
- Ruiz-Verdú, A.; Simis, S.G.H.; de Hoyos, C.; Gons, H.J.; Peña-Martínez, R. An Evaluation of Algorithms for the Remote Sensing of Cyanobacterial Biomass. Remote Sens. Environ. 2008, 112, 3996–4008. [Google Scholar] [CrossRef]
- Liu, M.; Ling, H.; Wu, D.; Su, X.; Cao, Z. Sentinel-2 and Landsat-8 Observations for Harmful Algae Blooms in a Small Eutrophic Lake. Remote Sens. 2021, 13, 4479. [Google Scholar] [CrossRef]
- Veerman, J.; Kumar, A.; Mishra, D.R. Exceptional Landscape-Wide Cyanobacteria Bloom in Okavango Delta, Botswana in 2020 Coincided with a Mass Elephant Die-off Event. Harmful Algae 2022, 111, 102145. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, E.; Coimbra, K.; Ogashawara, I.; Rodrigues, T.; Mantovani, J.; Rotta, L.H.; Park, E.; Fernandes Cunha, D.G. A Satellite-Based Investigation into the Algae Bloom Variability in Large Water Supply Urban Reservoirs during COVID-19 Lockdown. Remote Sens. Appl. 2021, 23, 100555. [Google Scholar] [CrossRef]
- Sòria-Perpinyà, X.; Vicente, E.; Urrego, P.; Pereira-Sandoval, M.; Ruíz-Verdú, A.; Delegido, J.; Soria, J.M.; Moreno, J. Remote Sensing of Cyanobacterial Blooms in a Hypertrophic Lagoon (Albufera of València, Eastern Iberian Peninsula) Using Multitemporal Sentinel-2 Images. Sci. Total Environ. 2020, 698, 134305. [Google Scholar] [CrossRef] [PubMed]
- Niroumand-Jadidi, M.; Bovolo, F. Deep Learning-Based Retrieval of an Orange Band Sensitive to Cyanobacteria for Landsat-8/9 and Sentinel-2. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2023, 16, 3929–3937. [Google Scholar] [CrossRef]
- Pérez-González, R.; Sòria-Perpinyà, X.; Soria, J.M.; Delegido, J.; Urrego, P.; Sendra, M.D.; Ruíz-Verdú, A.; Vicente, E.; Moreno, J. Phycocyanin Monitoring in Some Spanish Water Bodies with Sentinel-2 Imagery. Water 2021, 13, 2866. [Google Scholar] [CrossRef]
- Zamyadi, A.; Choo, F.; Newcombe, G.; Stuetz, R.; Henderson, R.K. A Review of Monitoring Technologies for Real-Time Management of Cyanobacteria: Recent Advances and Future Direction. TrAC-Trends Anal. Chem. 2016, 85, 83–96. [Google Scholar] [CrossRef]
- Ma, L.; Moradinejad, S.; Guerra Maldonado, J.F.; Zamyadi, A.; Dorner, S.; Prévost, M. Factors Affecting the Interpretation of Online Phycocyanin Fluorescence to Manage Cyanobacteria in Drinking Water Sources. Water 2022, 14, 3749. [Google Scholar] [CrossRef]
- Macário, I.P.; Castro, B.B.; Nunes, I.M.S.; Antunes, S.C.S.C.; Pizarro, C.; Coelho, C.; Gonçalves, F.; de Figueiredo, D.R. New Insights towards the Establishment of Phycocyanin Concentration Thresholds Considering Species-Specific Variability of Bloom-Forming Cyanobacteria. Hydrobiologia 2015, 757, 155–165. [Google Scholar] [CrossRef]
- Bunyon, C.L.; Fraser, B.T.; McQuaid, A.; Congalton, R.G. Using Imagery Collected by an Unmanned Aerial System to Monitor Cyanobacteria in New Hampshire, USA, Lakes. Remote Sens. 2023, 15, 2839. [Google Scholar] [CrossRef]
- Pershin, S.M.; Katsnelson, B.G.; Grishin, M.Y.; Lednev, V.N.; Zavozin, V.A.; Ostrovsky, I. Laser Remote Sensing of Lake Kinneret by Compact Fluorescence LiDAR. Sensors 2022, 22, 7307. [Google Scholar] [CrossRef]
- Wu, D.; Li, R.; Zhang, F.; Liu, J. A Review on Drone-Based Harmful Algae Blooms Monitoring. Environ. Monit. Assess. 2019, 191, 211. [Google Scholar] [CrossRef]
- Becker, R.H.; Sayers, M.; Dehm, D.; Shuchman, R.; Quintero, K.; Bosse, K.; Sawtell, R. Unmanned Aerial System Based Spectroradiometer for Monitoring Harmful Algal Blooms: A New Paradigm in Water Quality Monitoring. J. Great Lakes Res. 2019, 45, 444–453. [Google Scholar] [CrossRef]
- Johansen, R.A.; Beck, R.; Stumpf, R.; Lekki, J.; Tokars, R.; Tolbert, C.; McGhan, C.; Black, T.; Ma, O.; Xu, M.; et al. HABSat-1: Assessing the Feasibility of Using CubeSats for the Detection of Cyanobacterial Harmful Algal Blooms in Inland Lakes and Reservoirs. Lake Reserv. Manag. 2019, 35, 193–207. [Google Scholar] [CrossRef]
- Azevedo, R.; Rodriguez, E.; Figueiredo, D.; Peixoto, F.; Santos, C. Methodologies for the Study of Filamentous Cyanobacteria by Flow Cytometry. Fresenius Environ. Bull. 2012, 21, 679–684. [Google Scholar]
- Dashkova, V.; Malashenkov, D.; Poulton, N.; Vorobjev, I.; Barteneva, N.S. Imaging Flow Cytometry for Phytoplankton Analysis. Methods 2017, 112, 188–200. [Google Scholar] [CrossRef]
- Mirasbekov, Y.; Zhumakhanova, A.; Zhantuyakova, A.; Sarkytbayev, K.; Malashenkov, D.V.; Baishulakova, A.; Dashkova, V.; Davidson, T.A.; Vorobjev, I.A.; Jeppesen, E.; et al. Semi-Automated Classification of Colonial Microcystis by FlowCAM Imaging Flow Cytometry in Mesocosm Experiment Reveals High Heterogeneity during Seasonal Bloom. Sci. Rep. 2021, 11, 9377. [Google Scholar] [CrossRef]
- Kraft, K.; Seppälä, J.; Hällfors, H.; Suikkanen, S.; Ylöstalo, P.; Anglès, S.; Kielosto, S.; Kuosa, H.; Laakso, L.; Honkanen, M.; et al. First Application of IFCB High-Frequency Imaging-in-Flow Cytometry to Investigate Bloom-Forming Filamentous Cyanobacteria in the Baltic Sea. Front. Mar. Sci. 2021, 8, 594144. [Google Scholar] [CrossRef]
- George, G.; Menon, N.N.; Abdulaziz, A.; Brewin, R.J.W.; Pranav, P.; Gopalakrishnan, A.; Mini, K.G.; Kuriakose, S.; Sathyendranath, S.; Platt, T. Citizen Scientists Contribute to Real-Time Monitoring of Lake Water Quality Using 3D Printed Mini Secchi Disks. Front. Water 2021, 3, 662142. [Google Scholar] [CrossRef]
- Ho, S.Y.F.; Xu, S.J.; Lee, F.W.F. Citizen Science: An Alternative Way for Water Monitoring in Hong Kong. PLoS ONE 2020, 15, e0238349. [Google Scholar] [CrossRef]
- Siano, R.; Chapelle, A.; Antoine, V.; Michel-Guillou, E.; Rigaut-Jalabert, F.; Guillou, L.; Hégaret, H.; Leynaert, A.; Curd, A. Citizen Participation in Monitoring Phytoplankton Seawater Discolorations. Mar. Policy 2020, 117, 103039. [Google Scholar] [CrossRef]
- Cunha, D.G.F.; Casali, S.P.; de Falco, P.B.; Thornhill, I.; Loiselle, S.A. The Contribution of Volunteer-Based Monitoring Data to the Assessment of Harmful Phytoplankton Blooms in Brazilian Urban Streams. Sci. Total Environ. 2017, 584–585, 586–594. [Google Scholar] [CrossRef]
- Sanseverino, I.; Conduto António, D.; Loos, R.; Lettieri, T. Cyanotoxins: Methods and Approaches for Their Analysis and Detection; Joint Research Centre: Luxembourg, 2017. [Google Scholar]
- Komárek, J.; Mareš, J. An Update to Modern Taxonomy (2011) of Freshwater Planktic Heterocytous Cyanobacteria. Hydrobiologia 2012, 698, 327–351. [Google Scholar] [CrossRef]
- Thiel, S.U.; Wiltshire, R.J.; Davies, L.J. Automated Object Recognition of Blue-Green Algae for Measuring Water Quality—A Preliminary Study; Elsevier: Amsterdam, The Netherlands, 1995; Volume 29. [Google Scholar]
- Almesjö, L.; Rolff, C. Automated Measurements of Filamentous Cyanobacteria by Digital Image Analysis. Limnol. Oceanogr. Methods 2007, 5, 217–224. [Google Scholar] [CrossRef]
- Geronimo, J.O.-N.; Arguelles, E. Automated Classification and Identification System for Freshwater Algae Using Convolutional Neural Networks. Philipine J. Sci. 2023, 152, 325–335. [Google Scholar] [CrossRef]
- Chong, J.W.R.; Khoo, K.S.; Chew, K.W.; Vo, D.V.N.; Balakrishnan, D.; Banat, F.; Munawaroh, H.S.H.; Iwamoto, K.; Show, P.L. Microalgae Identification: Future of Image Processing and Digital Algorithm. Bioresour. Technol. 2023, 369, 128418. [Google Scholar] [CrossRef]
- Gaur, A.; Pant, G.; Jalal, A.S. Computer-Aided Cyanobacterial Harmful Algae Blooms (CyanoHABs) Studies Based on Fused Artificial Intelligence (AI) Models. Algal Res. 2022, 67, 102842. [Google Scholar] [CrossRef]
- Kraft, K.; Velhonoja, O.; Eerola, T.; Suikkanen, S.; Tamminen, T.; Haraguchi, L.; Ylöstalo, P.; Kielosto, S.; Johansson, M.; Lensu, L.; et al. Towards Operational Phytoplankton Recognition with Automated High-Throughput Imaging, near-Real-Time Data Processing, and Convolutional Neural Networks. Front. Mar. Sci. 2022, 9, 867695. [Google Scholar] [CrossRef]
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR Primers to Amplify 16S rRNA Genes from Cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef]
- Biegala, I.C.; Raimbault, P. High Abundance of Diazotrophic Picocyanobacteria (<3 μm) in a Southwest Pacific Coral Lagoon. Aquat. Microb. Ecol. 2008, 51, 45–53. [Google Scholar] [CrossRef]
- Wang, X.; Sun, M.; Wang, J.; Yang, L.; Luo, L.; Li, P.; Kong, F. Microcystis Genotype Succession and Related Environmental Factors in Lake Taihu during Cyanobacterial Blooms. Microb. Ecol. 2012, 64, 986–999. [Google Scholar] [CrossRef]
- de Figueiredo, D.R.; Pereira, M.J.; Castro, B.B.; Correia, A. Bacterioplankton Community Composition in Portuguese Water Bodies under a Severe Summer Drought. Community Ecol. 2012, 13, 185–193. [Google Scholar] [CrossRef]
- Yu, Z.; Zhou, J.; Yang, J.; Yu, X.; Liu, L. Vertical Distribution of Diazotrophic Bacterial Community Associated with Temperature and Oxygen Gradients in a Subtropical Reservoir. Hydrobiologia 2014, 741, 69–77. [Google Scholar] [CrossRef]
- Touzet, N.; McCarthy, D.; Gill, A.; Fleming, G.T.A. Comparative Summer Dynamics of Surface Cyanobacterial Communities in Two Connected Lakes from the West of Ireland. Sci. Total Environ. 2016, 553, 416–428. [Google Scholar] [CrossRef]
- Feng, L.; Liu, S.; Wu, W.; Ma, J.; Li, P.; Xu, H.; Li, N.; Feng, Y. Dominant Genera of Cyanobacteria in Lake Taihu and Their Relationships with Environmental Factors. J. Microbiol. 2016, 54, 468–476. [Google Scholar] [CrossRef]
- Kolmonen, E.; Sivonen, K.; Rapala, J.; Haukka, K. Diversity of Cyanobacteria and Heterotrophic Bacteria in Cyanobacterial Blooms in Lake Joutikas, Finland. Aquat. Microb. Ecol. 2004, 36, 201–211. [Google Scholar] [CrossRef]
- Eiler, A.; Drakare, S.; Bertilsson, S.; Pernthaler, J.; Peura, S.; Rofner, C.; Simek, K.; Yang, Y.; Znachor, P.; Lindström, E.S. Unveiling Distribution Patterns of Freshwater Phytoplankton by a next Generation Sequencing Based Approach. PLoS ONE 2013, 8, e53516. [Google Scholar] [CrossRef]
- Steffen, M.M.; Li, Z.; Effler, T.C.; Hauser, L.J.; Boyer, G.L.; Wilhelm, S.W. Comparative Metagenomics of Toxic Freshwater Cyanobacteria Bloom Communities on Two Continents. PLoS ONE 2012, 7, e44002. [Google Scholar] [CrossRef]
- Steven, B.; McCann, S.; Ward, N.L. Pyrosequencing of Plastid 23S RRNA Genes Reveals Diverse and Dynamic Cyanobacterial and Algal Populations in Two Eutrophic Lakes. FEMS Microbiol. Ecol. 2012, 82, 607–615. [Google Scholar] [CrossRef]
- Stüken, A.; Campbell, R.J.; Quesada, A.; Sukenik, A.; Dadheech, P.K.; Wiedner, C. Genetic and Morphologic Characterization of Four Putative Cylindrospermopsin Producing Species of the Cyanobacterial Genera Anabaena and Aphanizomenon. J. Plankton Res. 2009, 31, 465–480. [Google Scholar] [CrossRef]
- Lezcano, M.Á.; Velázquez, D.; Quesada, A.; El-Shehawy, R. Diversity and Temporal Shifts of the Bacterial Community Associated with a Toxic Cyanobacterial Bloom: An Interplay between Microcystin Producers and Degraders. Water Res. 2017, 125, 52–61. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Z.; Ding, A.; Wu, J.; Xiao, J.; Sun, Y.; Cheng, C.; Zaichao, Z.; Aizhong, D.; Jiayan, W.; et al. Bar-Coded Pyrosequencing Reveals the Bacterial Community during Microcystis Water Bloom in Guanting Reservoir, Beijing. Procedia Eng. 2011, 18, 341–346. [Google Scholar] [CrossRef]
- Salomon, P.S.; Janson, S.; Grane?li, E. Molecular Identification of Bacteria Associated with Filaments of Nodularia spumigena and Their Effect on the Cyanobacterial Growth. Harmful Algae 2003, 2, 261–272. [Google Scholar] [CrossRef]
- Berg, K.A.; Lyra, C.; Sivonen, K.; Paulin, L.; Suomalainen, S.; Tuomi, P.; Rapala, J. High Diversity of Cultivable Heterotrophic Bacteria in Association with Cyanobacterial Water Blooms. ISME J. 2008, 3, 314–325. [Google Scholar] [CrossRef]
- Shunyu, S.; Yongding, L.; Yinwu, S.; Genbao, L.; Dunhai, L. Lysis of Aphanizomenon flos-aquae (Cyanobacterium) by a Bacterium Bacillus cereus. Biol. Control. 2006, 39, 345–351. [Google Scholar] [CrossRef]
- Hennon, G.M.M.; Dyhrman, S.T. Progress and Promise of Omics for Predicting the Impacts of Climate Change on Harmful Algal Blooms. Harmful Algae 2020, 91, 101587. [Google Scholar] [CrossRef]
- Dziallas, C.; Pinnow, S.; Grossart, H.P. Quantification of Toxic and Toxin-Producing Cyanobacterial Cells by RING-FISH in Combination with Flow Cytometry. Limnol. Oceanogr. Methods 2011, 9, 67–73. [Google Scholar] [CrossRef]
- Legrand, B.; Lesobre, J.; Colombet, J.; Latour, D.; Sabart, M. Molecular Tools to Detect Anatoxin-a Genes in Aquatic Ecosystems: Toward a New Nested PCR-Based Method. Harmful Algae 2016, 58, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Caracciolo, A.B.; Dejana, L.; Fajardo, C.; Grenni, P.; Martin, M.; Mengs, G.; Sánchez-Fortún, S.; Lettieri, T.; Saccà, M.L.; Medlin, L.K. A New Fluorescent Oligonucleotide Probe for In-Situ Identification of Microcystis aeruginosa in Freshwater. Microchem. J. 2019, 148, 503–513. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Reilly, M.; Young, F.M.; Codd, G.A. Localization of Microcystin Synthetase Genes in Colonies of the Cyanobacterium Microcystis Using Fluorescence in Situ Hybridization. J. Phycol. 2009, 45, 1400–1404. [Google Scholar] [CrossRef]
- Zeller, P.; Méjean, A.; Biegala, I.; Contremoulins, V.; Ploux, O. Fluorescence in Situ Hybridization of Microcystis Strains Producing Microcystin Using Specific mRNA Probes. Lett. Appl. Microbiol. 2016, 63, 376–383. [Google Scholar] [CrossRef]
- Brient, L.; Ben Gamra, N.; Periot, M.; Roumagnac, M.; Zeller, P.; Bormans, M.; Méjean, A.; Ploux, O.; Biegala, I.C. Rapid Characterization of Microcystin-Producing Cyanobacteria in Freshwater Lakes by TSA-FISH (Tyramid Signal Amplification-Fluorescent in Situ Hybridization). Front. Environ. Sci. 2017, 5, 43. [Google Scholar] [CrossRef]
- Casero, M.C.; Velázquez, D.; Medina-Cobo, M.; Quesada, A.; Cirés, S. Unmasking the Identity of Toxigenic Cyanobacteria Driving a Multi-Toxin Bloom by High-Throughput Sequencing of Cyanotoxins Genes and 16S rRNA Metabarcoding. Sci. Total Environ. 2019, 665, 367–378. [Google Scholar] [CrossRef]
- Cunha, I.; Biltes, R.; Sales, M.G.F.; Vasconcelos, V. Aptamer-Based Biosensors to Detect Aquatic Phycotoxins and Cyanotoxins. Sensors 2018, 18, 2367. [Google Scholar] [CrossRef]
- Macário, I.P.E.; Castro, B.B.; Nunes, I.M.S.; Pizarro, C.; Coelho, C.; Gonçalves, F.; de Figueiredo, D.R. Stepwise Strategy for Monitoring Toxic Cyanobacterial Blooms in Lentic Water Bodies. Environ. Monit. Assess. 2017, 189, 620. [Google Scholar] [CrossRef]
- Ho, J.C.; Michalak, A.M. Challenges in Tracking Harmful Algal Blooms: A Synthesis of Evidence from Lake Erie. J. Great Lakes Res. 2015, 41, 317–325. [Google Scholar] [CrossRef]
- Srivastava, A.; Singh, S.; Ahn, C.Y.; Oh, H.M.; Asthana, R.K. Monitoring Approaches for a Toxic Cyanobacterial Bloom. Environ. Sci. Technol. 2013, 47, 8999–9013. [Google Scholar] [CrossRef] [PubMed]
- Douglas Greene, S.B.; LeFevre, G.H.; Markfort, C.D. Improving the Spatial and Temporal Monitoring of Cyanotoxins in Iowa Lakes Using a Multiscale and Multi-Modal Monitoring Approach. Sci. Total Environ. 2020, 760, 143327. [Google Scholar] [CrossRef] [PubMed]
- Steffen, M.M.; Belisle, B.S.; Watson, S.B.; Boyer, G.L.; Bourbonniere, R.A.; Wilhelm, S.W. Metatranscriptomic Evidence for Co-Occurring Top-down and Bottom-up Controls on Toxic Cyanobacterial Communities. Appl. Environ. Microbiol. 2015, 81, 3268–3276. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.A.; Carstensen, J. Global Observing for Phytoplankton? A Perspective. J. Plankton Res. 2023, 45, 221–234. [Google Scholar] [CrossRef]
Figure 1.
Main methodologies used in monitoring cyanobacterial blooms and their source type information as well as some associated limitations.
Figure 1.
Main methodologies used in monitoring cyanobacterial blooms and their source type information as well as some associated limitations.

Figure 2.
General scheme on a possible multilevel cross-referenced integrated database that gathers multi-tiered inputs from different information sources to feed models.
Figure 2.
General scheme on a possible multilevel cross-referenced integrated database that gathers multi-tiered inputs from different information sources to feed models.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
de Figueiredo, D.R. Harmful Cyanobacterial Blooms: Going beyond the “Green” to Monitor and Predict HCBs. Hydrobiology 2024, 3, 11-30. https://doi.org/10.3390/hydrobiology3010002
AMA Style
de Figueiredo DR. Harmful Cyanobacterial Blooms: Going beyond the “Green” to Monitor and Predict HCBs. Hydrobiology. 2024; 3(1):11-30. https://doi.org/10.3390/hydrobiology3010002
Chicago/Turabian Stylede Figueiredo, Daniela R. 2024. "Harmful Cyanobacterial Blooms: Going beyond the “Green” to Monitor and Predict HCBs" Hydrobiology 3, no. 1: 11-30. https://doi.org/10.3390/hydrobiology3010002