
Inhibitory Effect of Aqueous Extracts from Egeria densa Planch. on Cyanobacteria Microcystis aeruginosa (Kützing) Lemmermann Growth

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Stock Cultures
2.1.1. Microcystis aeruginosa (Kützing) Lemmermann Culture
2.1.2. Egeria densa Planch. Culture
E. densa Monoculture and Combined Culture with M. aeruginosa
2.2. E. densa Fresh Tissue Extraction Processes
2.3. Experimental Design
2.3.1. Application of E. densa Live Plants
2.3.2. Application of E. densa Extracts
2.4. Analytical Methods
2.4.1. Cell Density
Inhibition Rate (IR)
2.4.2. Chlorophyll-A
2.4.3. Nitrate (NO3−) and Phosphate (PO43−)
2.4.4. Three-Dimensional Excitation–Emission Matrix (3D EEM)
2.5. Data Analysis
3. Results
3.1. Effect of E. densa Live Plants on M. aeruginosa Growth during Coexistence
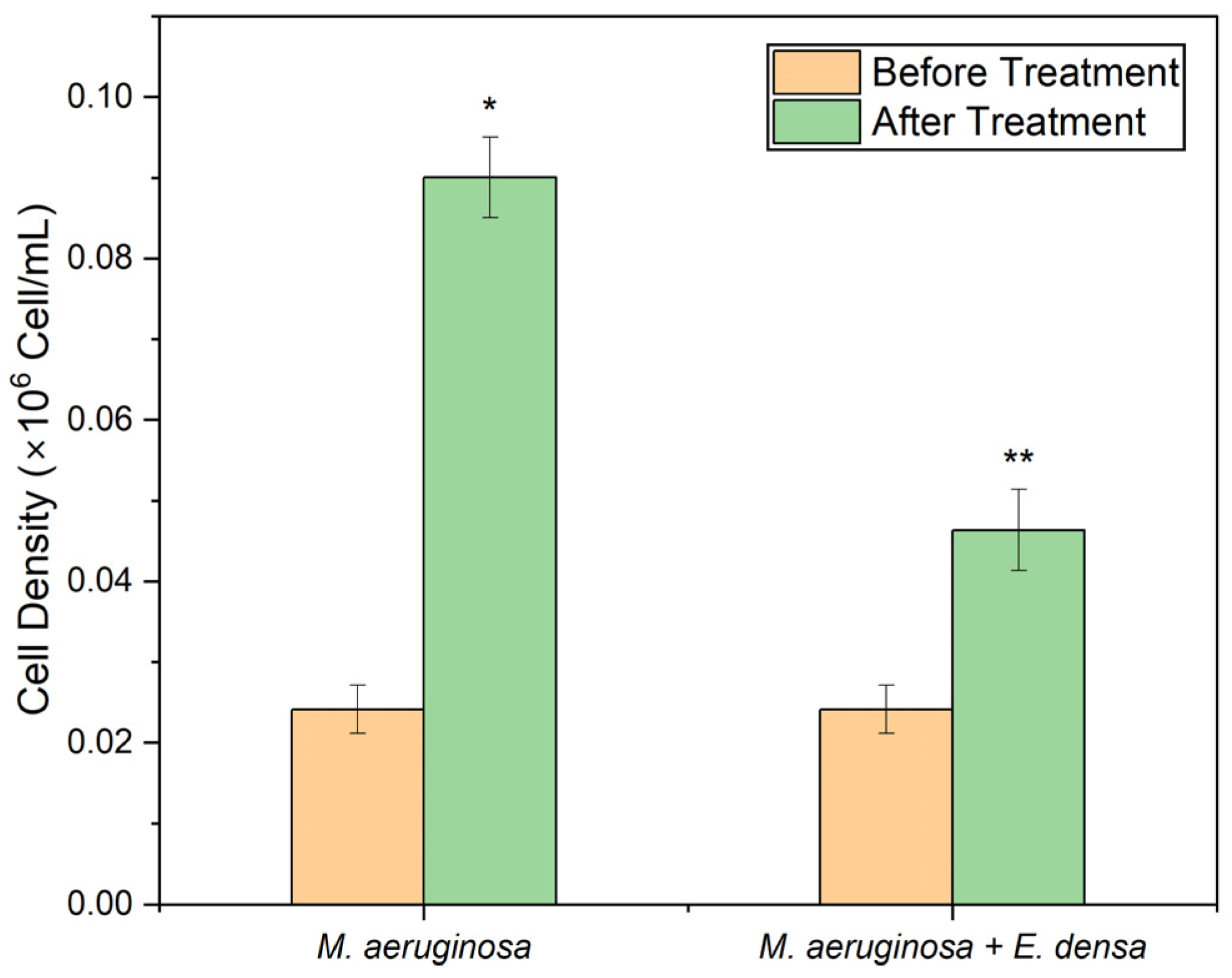
3.1.1. Cell Density
3.1.2. Protein Concentration
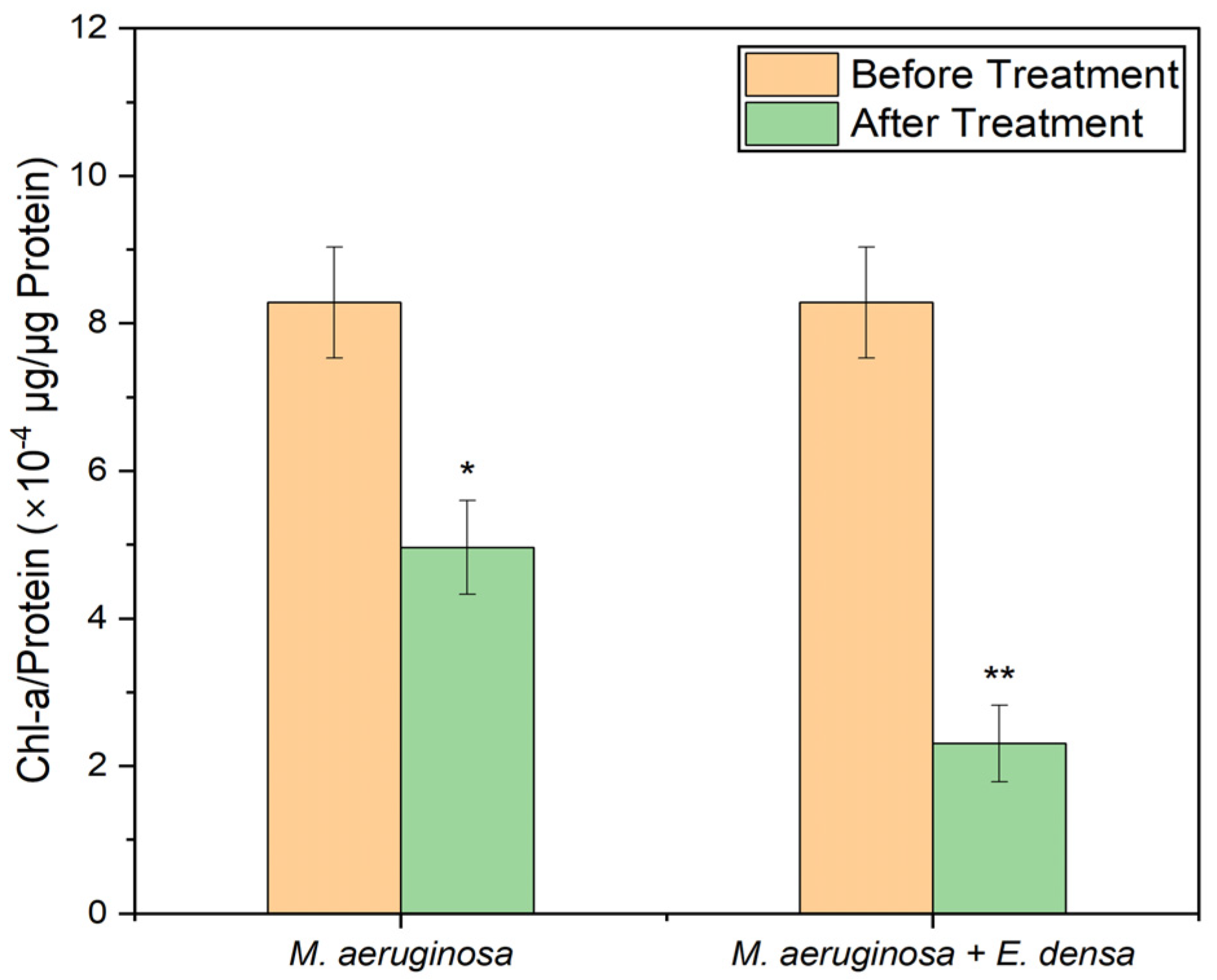
3.1.3. Chl-A Concentration
3.2. Effect of Different E. densa Extracts on Cyanobacteria Growth
3.2.1. Cell Density
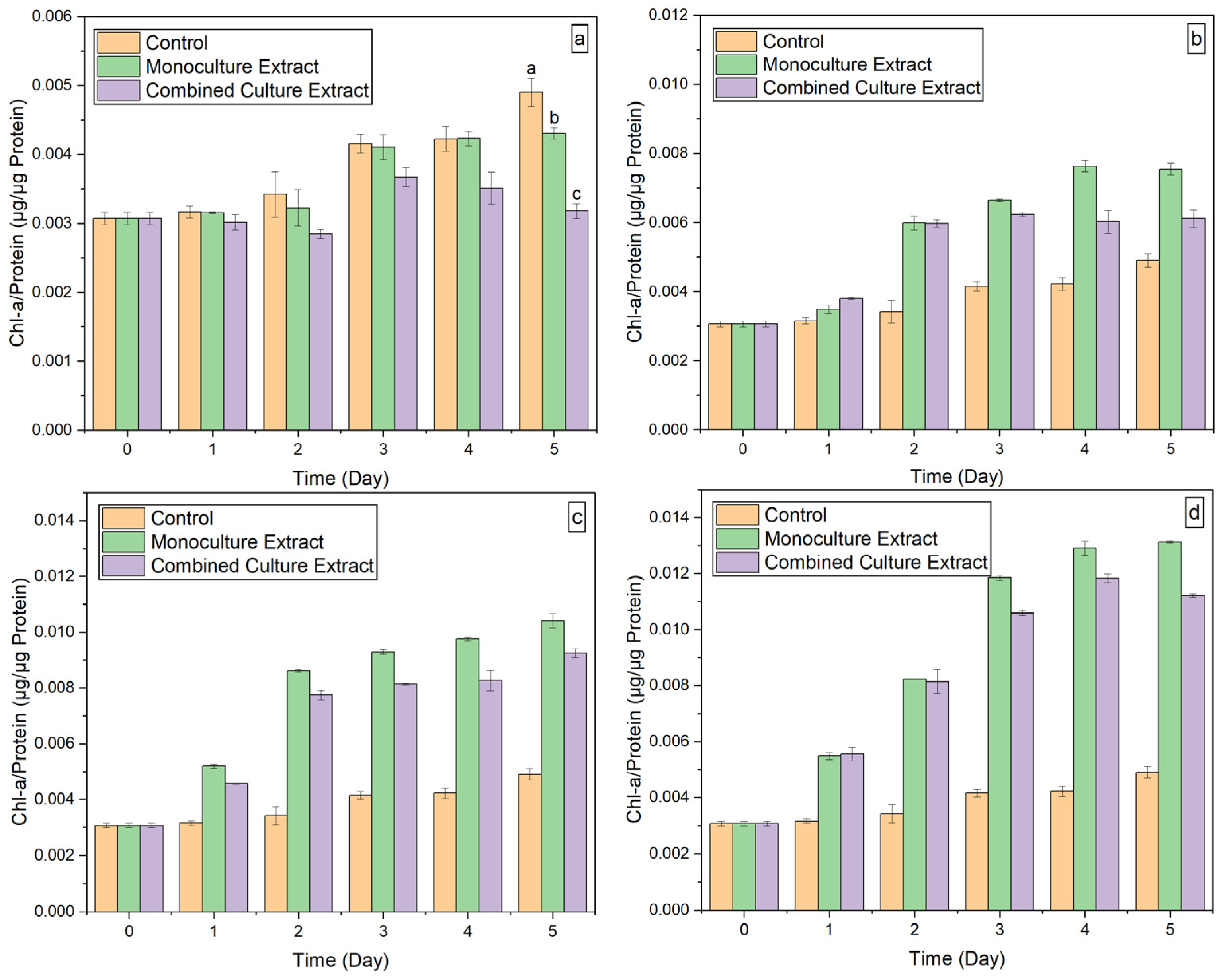
3.2.2. Chl-A Concentration
3.3. E. densa Extracts Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Mitigating Harmful Cyanobacterial Blooms in a Human- and Climatically-Impacted World. Life 2014, 4, 988–1012. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A Review of the Global Ecology, Genomics, and Biogeography of the Toxic Cyanobacterium, Microcystis Spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed]
- House, W.A. Geochemical Cycling of Phosphorus in Rivers. Appl. Geochem. 2003, 18, 739–748. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency Health Effects from Cyanotoxins. Available online: https://www.epa.gov/cyanohabs/health-effects-cyanotoxins (accessed on 21 November 2022).
- Dolman, A.M.; Rücker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and Cyanotoxins: The Influence of Nitrogen versus Phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef]
- Pokrzywinski, K.L.; Bishop, W.M.; Grasso, C.R.; Fernando, B.M.; Sperry, B.P.; Berthold, D.E.; Laughinghouse, H.D.; Van Goethem, E.M.; Volk, K.; Heilman, M.; et al. Evaluation of a Peroxide-Based Algaecide for Cyanobacteria Control: A Mesocosm Trial in Lake Okeechobee, FL, USA. Water 2022, 14, 169. [Google Scholar] [CrossRef]
- Sample, D.J.; Fox, L.J. Innovative Best Management Fact Sheet No.1: Floating Treatment Wetlands. Va. Polytech. Inst. State Univ. 2013, 1–5. [Google Scholar]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Taylor & Francis: Abingdon, UK, 2021; ISBN 9780367533311. [Google Scholar]
- Chen, L.; Giesy, J.P.; Adamovsky, O.; Svirčev, Z.; Meriluoto, J.; Codd, G.A.; Mijovic, B.; Shi, T.; Tuo, X.; Li, S.C.; et al. Challenges of Using Blooms of Microcystis Spp. in Animal Feeds: A Comprehensive Review of Nutritional, Toxicological and Microbial Health Evaluation. Sci. Total Environ. 2021, 764, 142319. [Google Scholar] [CrossRef]
- Nakai, S.; Inoue, Y.; Hosomi, M.; Murakami, A. Growth Inhibition of Blue-Green Algae by Allelopathic Effects of Macrophytes. Water Sci. Technol. 1999, 39, 47–53. [Google Scholar] [CrossRef]
- Tazart, Z.; Manganelli, M.; Scardala, S.; Buratti, F.M.; Di Gregorio, F.N.; Douma, M.; Mouhri, K.; Testai, E.; Loudiki, M. Remediation Strategies to Control Toxic Cyanobacterial Blooms: Effects of Macrophyte Aqueous Extracts on Microcystis aeruginosa (Growth, Toxin Production and Oxidative Stress Response) and on Bacterial Ectoenzymatic Activities. Microorganisms 2021, 9, 1782. [Google Scholar] [CrossRef]
- Kurashov, E.; Krylova, J.; Protopopova, E. The Use of Allelochemicals of Aquatic Macrophytes to Suppress the Development of Cyanobacterial “Blooms”. In Plankton Communities; Pereira, L., Gonçalves, A.M., Eds.; Intech Open: London, UK, 2021. [Google Scholar] [CrossRef]
- Maredová, N.; Altman, J.; Kaštovský, J. The Effects of Macrophytes on the Growth of Bloom-Forming Cyanobacteria: Systematic Review and Experiment. Sci. Total Environ. 2021, 792, 148413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shang, Y.; Lyu, T.; Chen, J.; Pan, G. Switching Harmful Algal Blooms to Submerged Macrophytes in Shallow Waters Using Geo-Engineering Methods: Evidence from a 15N Tracing Study. Environ. Sci. Technol. 2018, 52, 11778–11785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.H.; Guo, L.; Cao, J.Y.; Chang, J.J. Allelopathic Activities of Three Emergent Macrophytes on Several Monospecific Cyanobacterial Species and Natural Phytoplankton Assemblages. Polish J. Environ. Stud. 2015, 24, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Amorim, C.A.; de Moura-Falcão, R.H.; Valença, C.R.; de Souza, V.R.; Moura, A.D.N. Allelopathic Effects of the Aquatic Macrophyte Ceratophyllum demersum L. on Phytoplankton Species: Contrasting Effects between Cyanobacteria and Chlorophytes. Acta Limnol. Bras. 2019, 31, e21. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, B.; Wang, J.; Gao, Y.; Wu, Z. Study on the Mechanism of Allelopathic Influence on Cyanobacteria and Chlorophytes by Submerged Macrophyte (Myriophyllum spicatum) and Its Secretion. Aquat. Toxicol. 2010, 98, 196–203. [Google Scholar] [CrossRef]
- Wang, H.; Liu, F.; Luo, P.; Li, Z.; Zheng, L.; Wang, H.; Zou, D.; Wu, J. Allelopathic Effects of Myriophyllum aquaticum on Two Cyanobacteria of Anabaena flos-aquae and Microcystis aeruginosa. Bull. Environ. Contam. Toxicol. 2017, 98, 556–561. [Google Scholar] [CrossRef]
- Park, M.H.; Chung, I.M.; Ahmad, A.; Kim, B.H.; Hwang, S.J. Growth Inhibition of Unicellular and Colonial Microcystis Strains (Cyanophyceae) by Compounds Isolated from Rice (Oryza sativa) Hulls. Aquat. Bot. 2009, 90, 309–314. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, J.C.; Lee, S.; Lee, B.H.; Cho, K.Y. Biological Activity of L-2-Azetidinecarboxylic Acid, Isolated from Polygonatum odoratum Var. Pluriflorum, against Several Algae. Aquat. Bot. 2006, 85, 1–6. [Google Scholar] [CrossRef]
- Declerck, S.; Vanderstukken, M.; Pals, A.; Muylaert, K.; De Meester, L. Plankton Biodiversity along a Gradient of Productivity and Its Mediation by Macrophytes. Ecology 2007, 88, 2199–2210. [Google Scholar] [CrossRef]
- Vanderstukken, M.; Mazzeo, N.; van Colen, W.; Declerck, S.A.J.; Muylaert, K. Biological Control of Phytoplankton by the Subtropical Submerged Macrophytes Egeria densa and Potamogeton illinoensis: A Mesocosm Study. Freshw. Biol. 2011, 56, 1837–1849. [Google Scholar] [CrossRef]
- Senavirathna, M.D.H.J.; Muhetaer, G.; Atapaththu, K.S.S.; Fujino, T. Egeria densa Allelopathy on Microcystis aeruginosa Under Different Light Intensities and Preliminary Insight into Inter-Parameter Relationships. Water. Air. Soil Pollut. 2021, 232, 135. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, H.; Han, Z.; Ye, J.; Liu, Z. The Influence of Aquatic Macrophytes on Microcystis aeruginosa Growth. Ecol. Eng. 2012, 42, 130–133. [Google Scholar] [CrossRef]
- Ghobrial, M.G.; Nassr, H.S.; Kamil, A.W. Bioactivity Effect of Two Macrophyte Extracts on Growth Performance of Two Bloom-Forming Cyanophytes. Egypt. J. Aquat. Res. 2015, 41, 69–81. [Google Scholar] [CrossRef]
- Gross, E.M.; Erhard, D.; Iványi, E. Allelopathic Activity of Ceratophyllum demersum L. and Najas marina Ssp. Intermedia (Wolfgang) Casper. Hydrobiologia 2003, 506–509, 583–589. [Google Scholar] [CrossRef]
- Mulderij, G.; Smolders, A.J.P.; Van Donk, E. Allelopathic Effect of the Aquatic Macrophyte, Stratiotes aloides, on Natural Phytoplankton. Freshw. Biol. 2006, 51, 554–561. [Google Scholar] [CrossRef]
- Senavirathna, M.D.H.J.; Wijesinghe, N.A.; Liu, Z.; Fujino, T. Effects of Short-Term Exposure to Different Salinity Levels on Myriophyllum spicatum and Ceratophyllum demersum and Suitability of Biomarkers to Evaluate Macrophyte Responses to Salinity Stress. Ann. Limnol. 2020, 56, 23. [Google Scholar] [CrossRef]
- Hu, H.; Hong, Y. Algal-Bloom Control by Allelopathy of Aquatic Macrophytes—A Review. Front. Environ. Sci. Eng. China 2008, 2, 421–438. [Google Scholar] [CrossRef]
- Espinosa-Rodríguez, C.A.; Rivera-De La Parra, L.; Martínez-Téllez, A.; Gómez-Cabral, G.C.; Sarma, S.S.S.; Nandini, S. Allelopathic Interactions between the Macrophyte Egeria densa and Plankton (Alga, Scenedesmus acutus and Cladocerans, Simocephalus Spp.): A Laboratory Study. J. Limnol. 2016, 75, 151–160. [Google Scholar] [CrossRef]
- Virginia, V.; Ranielle, B.; Silva, S.; Severiano, S. Can the Presence of Green Microalgae Reverse the Allelopathic Effects of the Submerged Macrophyte Egeria densa on the Toxin-Producing Cyanobacterium Raphidiopsis raciborskii? Hydrobiologia 2022, 849, 4391–4406. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Zhang, H.; Ye, Y.K.; Wang, S.H.; Luo, J.P.; Tang, J.; Ma, D.F. Hydrogen Sulfide Counteracts Chlorophyll Loss in Sweetpotato Seedling Leaves and Alleviates Oxidative Damage against Osmotic Stress. Plant Growth Regul. 2009, 58, 243–250. [Google Scholar] [CrossRef]
- Pilotto, L.S.; Douglas, R.M.; Burch, M.D.; Cameron, S.; Beers, M.; Rouch, G.J.; Robinson, P.; Kirk, M.; Cowie, C.T.; Hardiman, S.; et al. Health Effects of Exposure to Cyanobacteria (Blue-Green Algae) during Recreational Water-Related Activities. Aust. N. Z. J. Public Health 1997, 21, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Hua, Q.; Liu, Y.G.; Yan, Z.L.; Zeng, G.M.; Liu, S.B.; Wang, W.J.; Tan, X.F.; Deng, J.Q.; Tang, X.; Wang, Q.P. Allelopathic Effect of the Rice Straw Aqueous Extract on the Growth of Microcystis aeruginosa. Ecotoxicol. Environ. Saf. 2018, 148, 953–959. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Y.; Yin, H.; Wang, Y.; Li, H.; Wang, Z.; Geng, Y.; Liang, W.; Wang, H. Effect of Aquatic Macrophyte Growth on Landscape Water Quality Improvement. Environ. Sci. Pollut. Res. 2019, 26, 33791–33803. [Google Scholar] [CrossRef]
- Hu, L.; Yin, C. Development of a New Three-Dimensional Fluorescence Spectroscopy Method Coupling with Multilinear Pattern Recognition to Discriminate the Variety and Grade of Green Tea. Food Anal. Methods 2017, 10, 2281–2292. [Google Scholar] [CrossRef]
- Körner, S.; Nicklisch, A. Allelopathic Growth Inhibition of Selected Phytoplankton Species by Submerged Macrophytes. J. Phycol. 2002, 871, 862–871. [Google Scholar] [CrossRef]
- Norling, B.; Zak, E.; Andersson, B.; Pakrasi, H. 2D-Isolation of Pure Plasma and Thylakoid Membranes from the Cyanobacterium Synechocystis Sp. PCC 6803. Fed. Eur. Biochem. Soc. Lett. 1998, 436, 189–192. [Google Scholar] [CrossRef]
- Li, H.; Singh, A.K.; McIntyre, L.M.; Sherman, L.A. Differential Gene Expression in Response to Hydrogen Peroxide and the Putative PerR Regulon of Synechocystis Sp. Strain PCC 6803. J. Bacteriol. 2004, 186, 3331–3345. [Google Scholar] [CrossRef]
- Mohamed, Z.A. Macrophytes-Cyanobacteria Allelopathic Interactions and Their Implications for Water Resources Management—A Review. Limnologica 2017, 63, 122–132. [Google Scholar] [CrossRef]
- Wang, F.; Zhao, W.; Chen, J.; Zhou, Y. Allelopathic Inhibitory Effect on the Growth of Microcystis aeruginosa by Improved Ultrasonic-Cellulase Extract of Vallisneria. Chemosphere 2022, 298, 134245. [Google Scholar] [CrossRef]
- Traversa, A.; Loffredo, E.; Gattullo, C.E.; Senesi, N. Water-Extractable Organic Matter of Different Composts: A Comparative Study of Properties and Allelochemical Effects on Horticultural Plants. Geoderma 2010, 156, 287–292. [Google Scholar] [CrossRef]
- Rossi, F.; De Philippis, R. Role of Cyanobacterial Exopolysaccharides in Phototrophic Biofilms and in Complex Microbial Mats. Life 2015, 5, 1218–1238. [Google Scholar] [CrossRef] [PubMed]
- Tazart, Z.; Douma, M.; Tebaa, L.; Loudiki, M. Use of Macrophytes Allelopathy in the Biocontrol of Harmful Microcystis aeruginosa Blooms. Water Sci. Technol. Water Supply 2019, 19, 245–253. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wijesinghe, A.; Senavirathna, M.D.H.J.; Fujino, T. Inhibitory Effect of Aqueous Extracts from Egeria densa Planch. on Cyanobacteria Microcystis aeruginosa (Kützing) Lemmermann Growth. Hydrobiology 2023, 2, 491-506. https://doi.org/10.3390/hydrobiology2030033
Wijesinghe A, Senavirathna MDHJ, Fujino T. Inhibitory Effect of Aqueous Extracts from Egeria densa Planch. on Cyanobacteria Microcystis aeruginosa (Kützing) Lemmermann Growth. Hydrobiology. 2023; 2(3):491-506. https://doi.org/10.3390/hydrobiology2030033
Chicago/Turabian StyleWijesinghe, Ashika, Mudalige Don Hiranya Jayasanka Senavirathna, and Takeshi Fujino. 2023. "Inhibitory Effect of Aqueous Extracts from Egeria densa Planch. on Cyanobacteria Microcystis aeruginosa (Kützing) Lemmermann Growth" Hydrobiology 2, no. 3: 491-506. https://doi.org/10.3390/hydrobiology2030033