
Evaluation of Biogeochemical Changes in Channelized and Restored Portions of a Subtropical Floodplain
Abstract
:1. Introduction
2. Materials and Methods
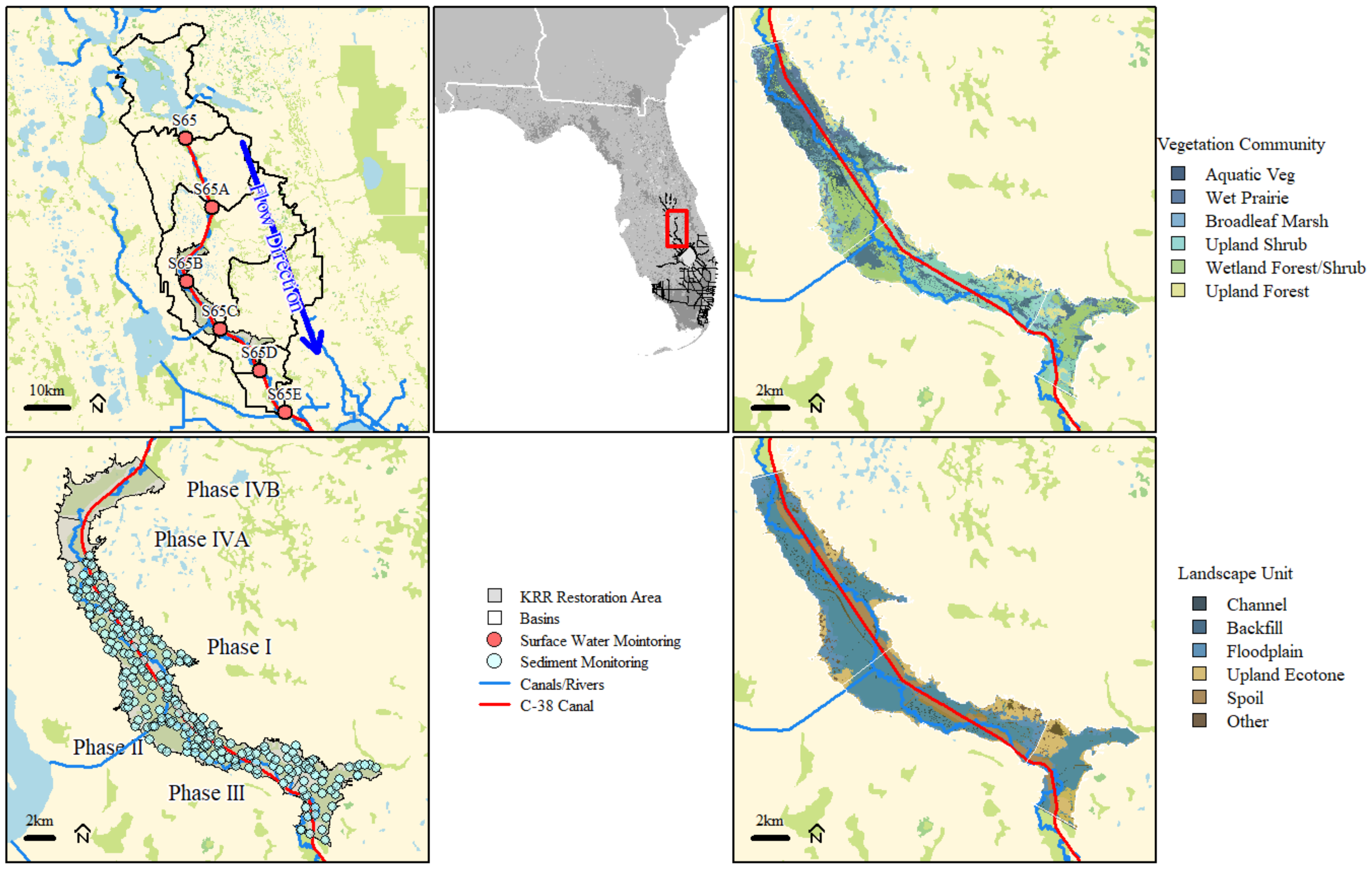
2.1. Study Area
2.2. Data Sources
2.3. Data Analysis
3. Results
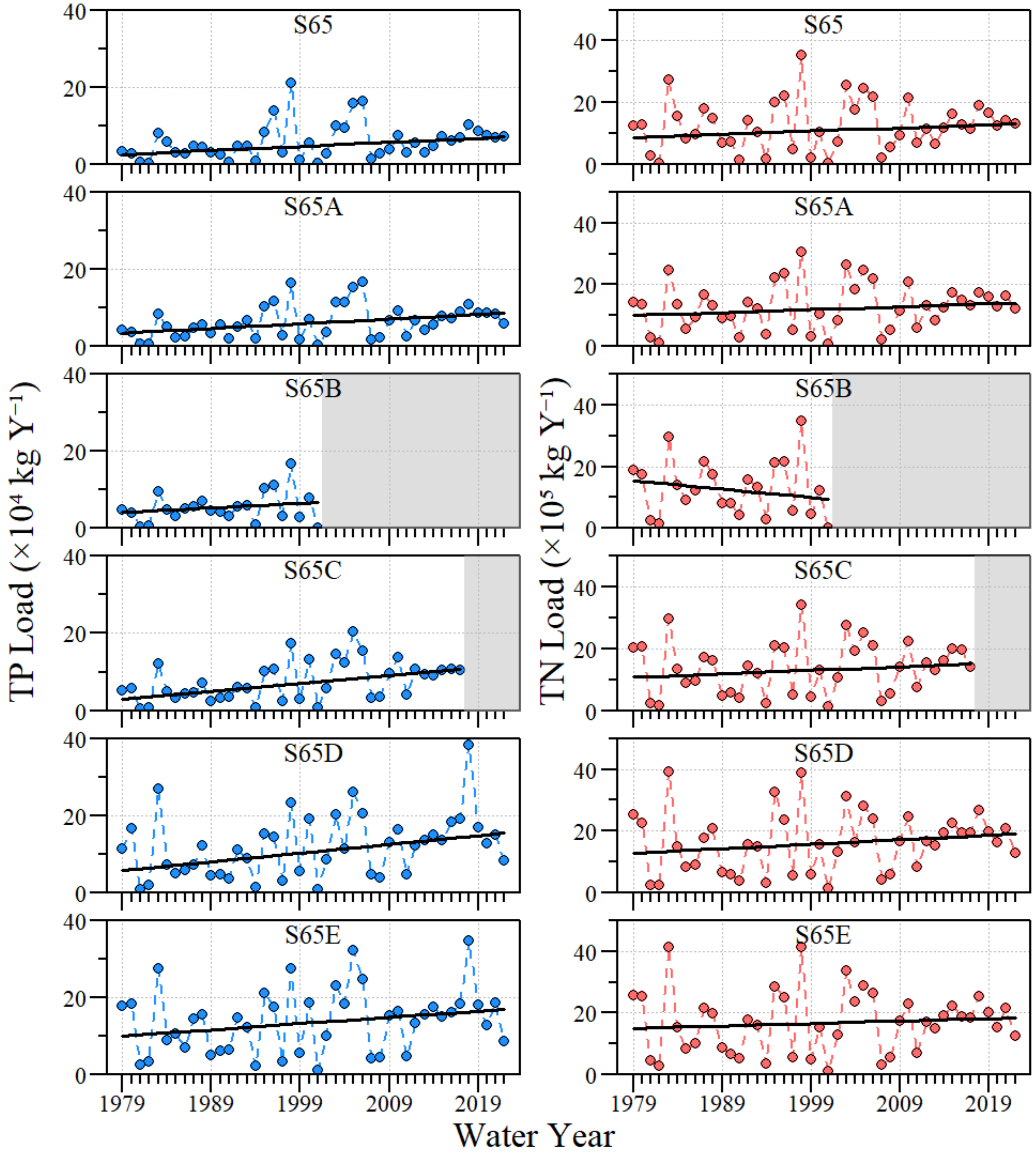
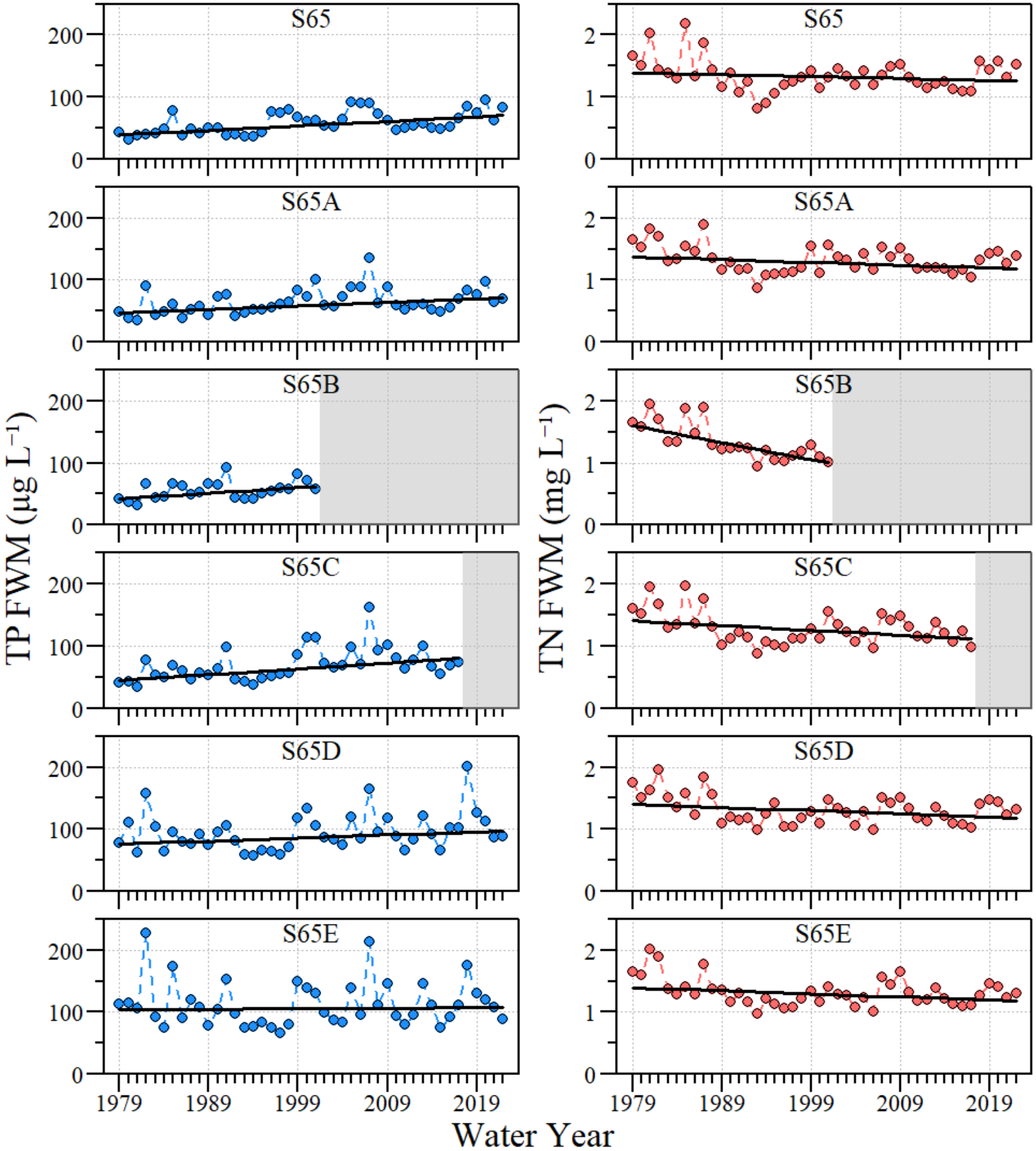
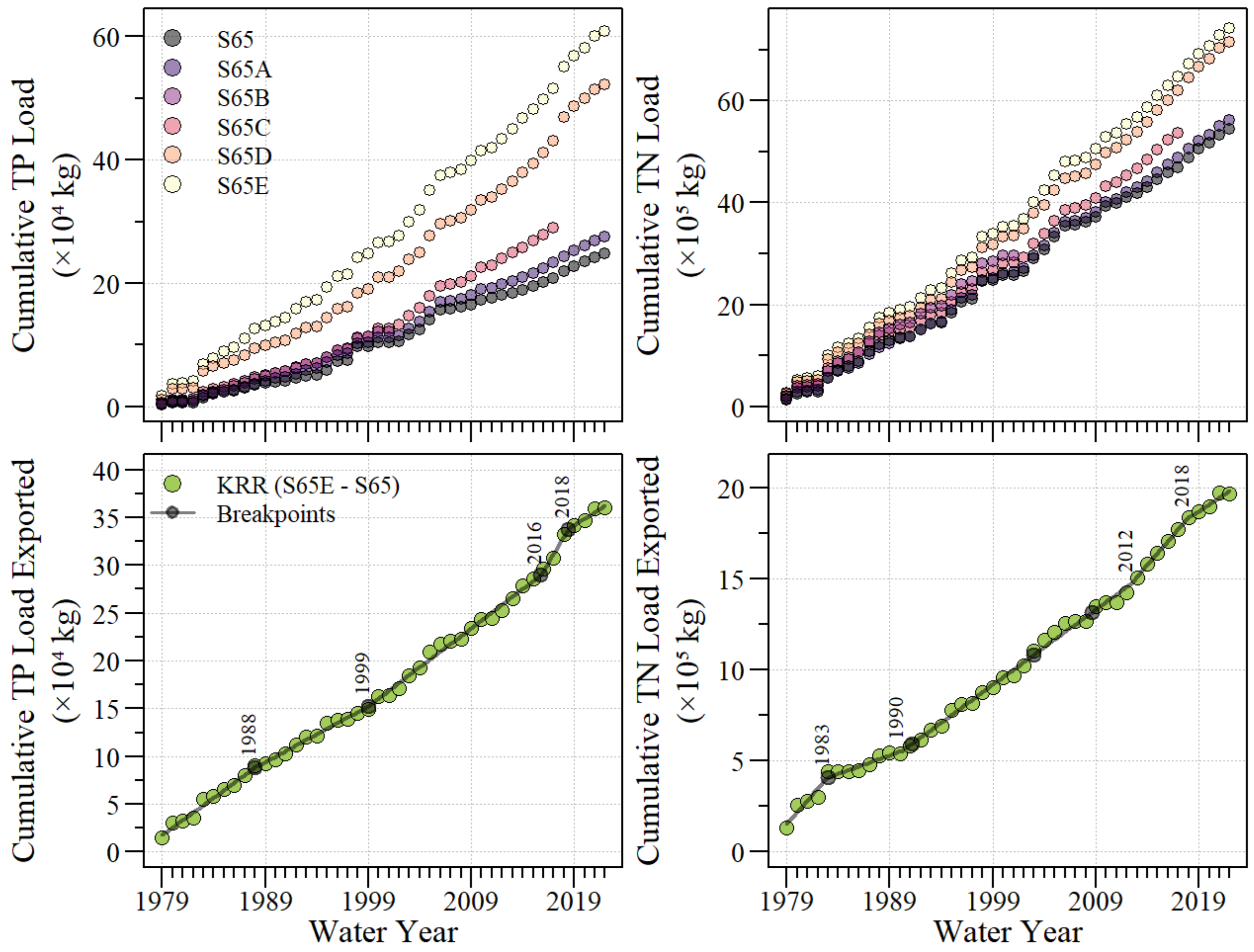
3.1. Water Quality Conditions
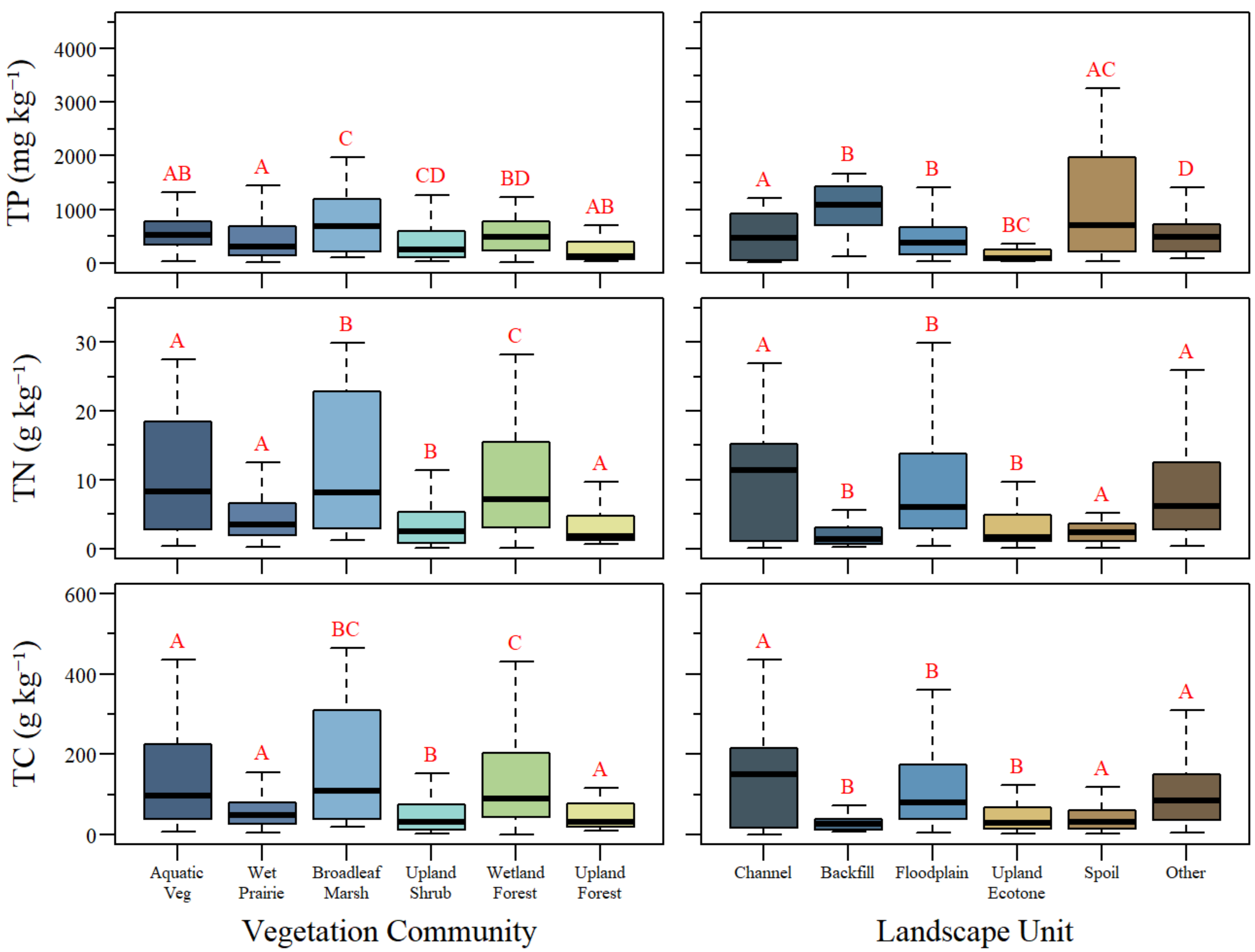
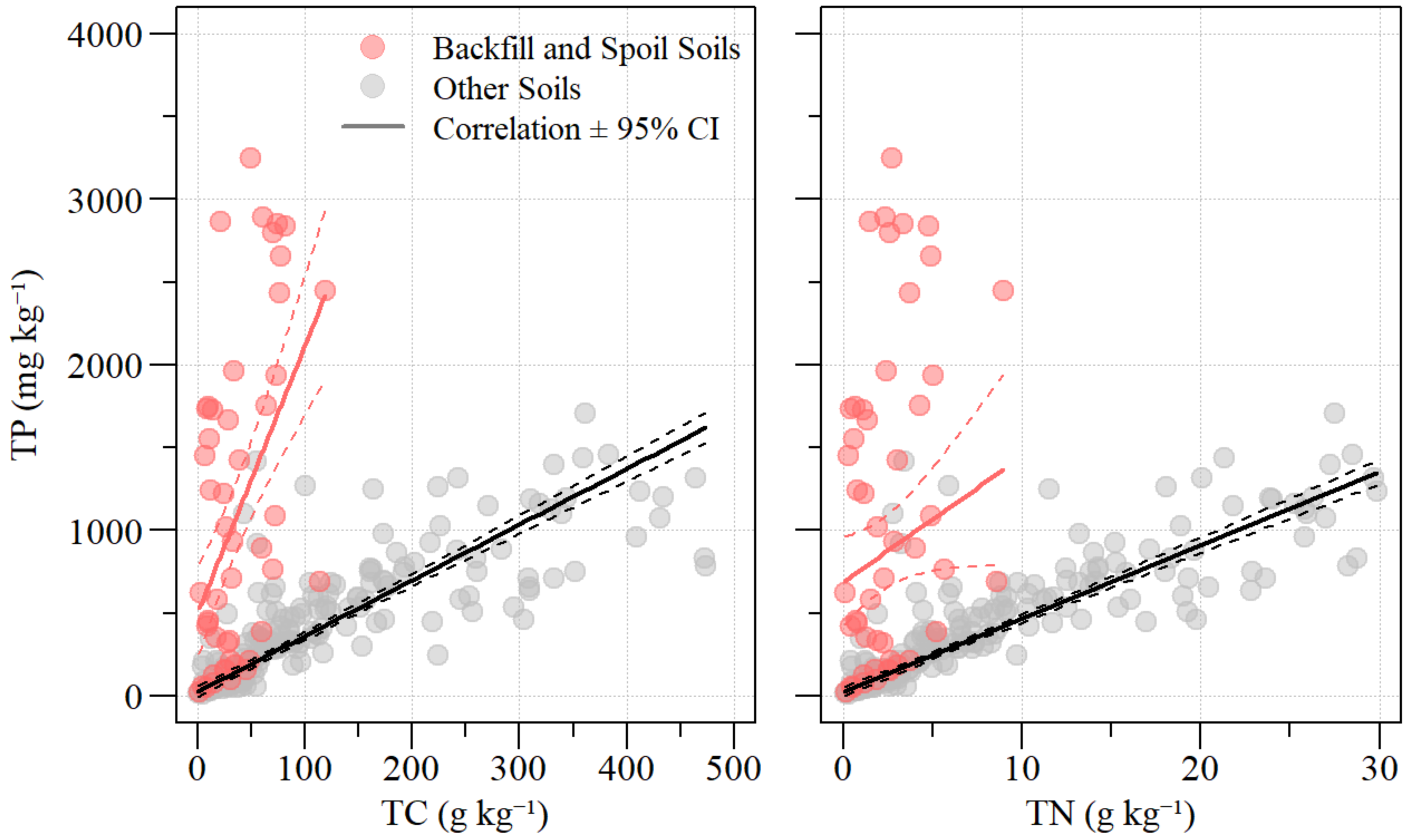
3.2. Soil Nutrient Distribution
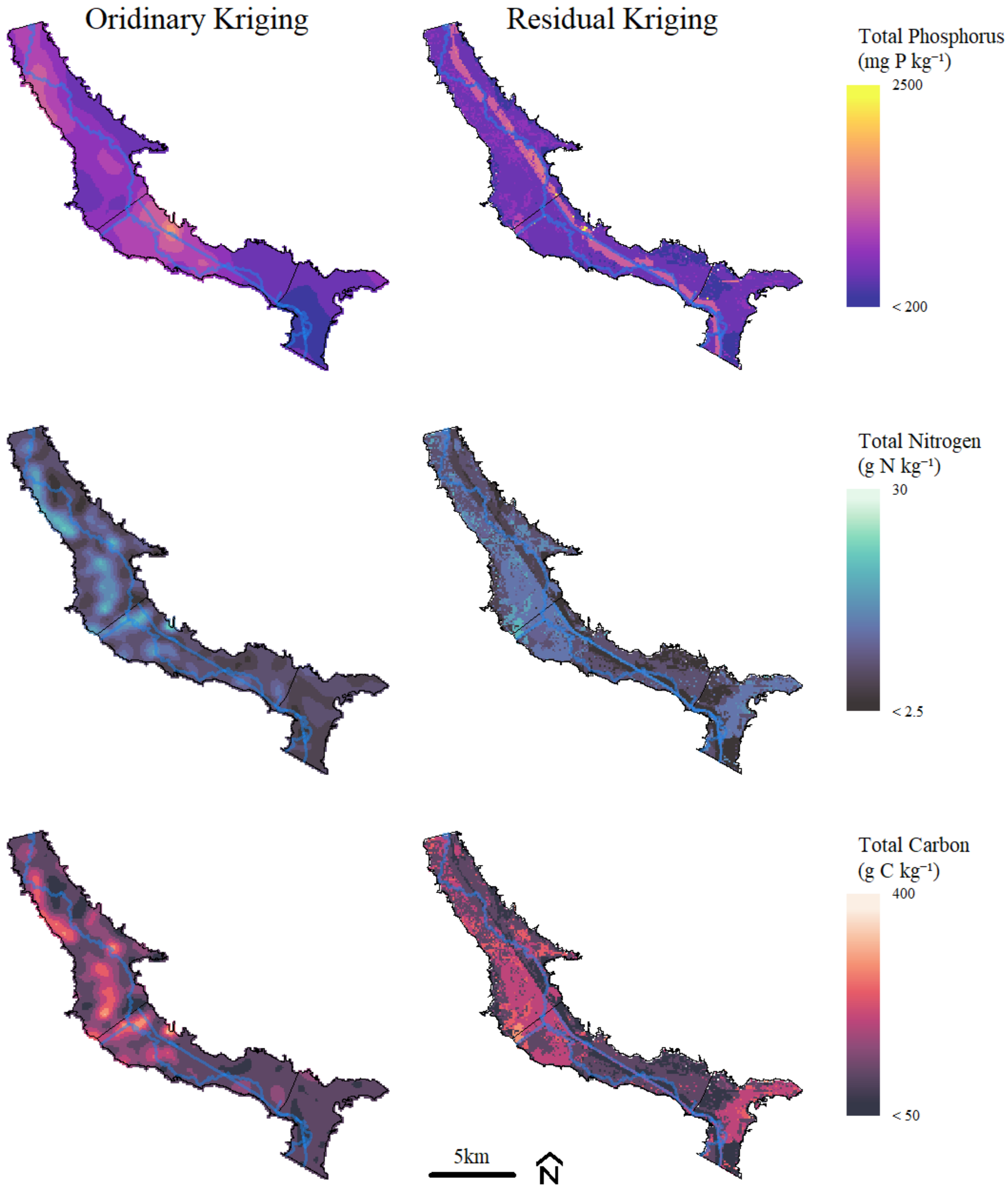
3.3. Geostatistical Analysis
4. Discussion
4.1. River–Floodplain Nutrient Loading
4.2. Floodplain Soils
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wohl, E. An Integrative Conceptualization of Floodplain Storage. Rev. Geophys. 2021, 59, e2020RG000724. [Google Scholar] [CrossRef]
- Nanson, G.C.; Croke, J.C. A Genetic Classification of Floodplains. Geomorphology 1992, 4, 459–486. [Google Scholar] [CrossRef] [Green Version]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The Flood Pulse Concept in River-Floodplain Systems. Can. Spec. Publ. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Harvey, J.; Gooseff, M. River Corridor Science: Hydrologic Exchange and Ecological Consequences from Bedforms to Basins. Water Resour. Res. 2015, 51, 6893–6922. [Google Scholar] [CrossRef] [Green Version]
- Xenopoulos, M.A.; Barnes, R.T.; Boodoo, K.S.; Butman, D.; Catalán, N.; D’Amario, S.C.; Fasching, C.; Kothawala, D.N.; Pisani, O.; Solomon, C.T.; et al. How Humans Alter Dissolved Organic Matter Composition in Freshwater: Relevance for the Earth’s Biogeochemistry. Biogeochemistry 2021, 154, 323–348. [Google Scholar] [CrossRef]
- Knox, R.L.; Morrison, R.R.; Wohl, E.E. A River Ran through It: Floodplains as America’s Newest Relict Landform. ScienceAdvances 2022, 8, 6. [Google Scholar] [CrossRef]
- Hupp, C.R.; Pierce, A.R.; Noe, G.B. Floodplain Geomorphic Processes and Environmental Impacts of Human Alteration along Coastal Plain Rivers, USA. Wetlands 2009, 29, 413–429. [Google Scholar] [CrossRef]
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and Flow Regulation of the World’s Large River Systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Acreman, M.C.; Riddington, R.; Booker, D.J. Hydrological Impacts of Floodplain Restoration: A Case Study of the River Cherwell, UK. Hydrol. Earth Syst. Sci. 2003, 7, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Toth, L.A.; Arrington, D.A.; Brady, M.A.; Muszick, D.A. Conceptual Evaluation of Factors Potentially Affecting Restoration of Habitat Structure within the Channelized Kissimmee River Ecosystem. Restor. Ecol. 1995, 3, 160–180. [Google Scholar] [CrossRef]
- Anderson, D.H.; Bousquin, S.G.; William, G.E.; Colangelo, D.J. Defining Success: Expectations for Restoration of the Kissimmee River; South Florida Water Management District: West Palm Beach, FL, USA, 2005. [Google Scholar]
- Koebel, J.W.; Bousquin, S.G. The Kissimmee River Restoration Project and Evaluation Program, Florida, U.S.A. Restor. Ecol. 2014, 22, 345–352. [Google Scholar] [CrossRef]
- Toth, L.A. Restoration Response of Relict Broadleaf Marshes to Increased Water Depths. Wetlands 2010, 30, 263–274. [Google Scholar] [CrossRef]
- Koebel, J.W. An Historical Perspective on the Kissimmee River Restoration Project. Restor. Ecol. 1995, 3, 149–159. [Google Scholar] [CrossRef]
- Koebel, J.W.; Bousquin, S.G.; Anderson, D.H.; Cheek, M.D.; Carroll, C.; Chen, H.; Hanlon, C.; Welch, Z.; Anderson, B.C.; Spencer, L.; et al. Kissimmee River Restoration and Basin Initiatives. In South Florida Environmental Report; South Florida Water Management District: West Palm Beach, FL, USA, 2019; Volume 1. [Google Scholar]
- Koebel, J.W.; Bousquin, S.G.; Anderson, D.H.; Cheek, M.D.; Carroll, C.; Marois, D.; Botta, R.; Hanlon, C.; Anderson, B.C.; Beck, T.; et al. Kissimmee River Restoration and Other Kissimmee Basin Initiatives. In 2020 South Florida Environmental Report; South Florida Water Management District: West Palm Beach, FL, USA, 2022. [Google Scholar]
- Zhang, J.; Burke, P.; Iricanin, N.; Hill, S.; Gray, S.; Budell, R. Long-Term Water Quality Trends in the Lake Okeechobee Watershed, Florida. Crit. Rev. Environ. Sci. Technol. 2011, 41, 548–575. [Google Scholar] [CrossRef]
- Hiscock, J.G.; Thourot, C.S.; Zhang, J. Phosphorus Budget—Land Use Relationships for the Northern Lake Okeechobee Watershed, Florida. Ecol. Eng. 2003, 21, 63–74. [Google Scholar] [CrossRef]
- Schenk, E.R.; Hupp, C.R.; Gellis, A. Sediment Dynamics in the Restored Reach of the Kissimmee River Basin, Florida: A Vast Subtropical Riparian Wetland: Sediment Dynamics in the Restored Reach of the Kissimmee River. River Res. Applic. 2012, 28, 1753–1767. [Google Scholar] [CrossRef]
- U.S. Army Corps of Engineers Central and Southern Florida. Kissimmee River, Florida Final Feasibility Report and Environmental Impact Statement; U.S. Army Corps of Engineers: Jacksonville, FL, USA, 1985. [Google Scholar]
- Establishing a Baseline: Pre-Restoration Studies of the Channelized Kissimmee River. In Kissimmee River Restoration Studies; Bousquin, S.G.; Anderson, D.H.; Williams, G.E.; Colangelo, D.J. (Eds.) South Florida Water Management District: West Palm Beach, FL, USA, 2005; Volume 1. [Google Scholar]
- U.S. Army Corps of Engineers Central and Southern Florida. Project Final Integrated Feasibility Report and Environmental Impact Statement Environmental Restoration Kissimmee River, Florida; U.S. Army Corps of Engineers: Jacksonville, FL, USA, 1991. [Google Scholar]
- U.S. Army Corps of Engineers Central and Southern Florida. Project Kissimmee River Headwaters Revitalization Project Integrated Project Modification Report and Supplement to the Final Environmental Impact Statement; U.S. Army Corps of Engineers: Jacksonville, FL, USA, 1996. [Google Scholar]
- Koebel, J.W.; Bousquin, S.G.; Anderson, D.H.; Welch, Z.; Cheek, M.D.; Chen, H.; James, R.T.; Zhang, J.; Anderson, B.C.; Baird, R.; et al. Kissimmee River Restoration and Other Kissimmee Basin Initiatives. In 2016 South Florida Environmental Report; South Florida Water Management District: West Palm Beach, FL, USA, 2016. [Google Scholar]
- Marazzi, L.; Finlayson, C.M.; Gell, P.A.; Julian, P.; Kominoski, J.S.; Gaiser, E.E. Balancing Wetland Restoration Benefits to People and Nature. Solut. J. 2018, 1–9. [Google Scholar]
- Perry, W.B. Everglades Restoration and Water Quality Challenges in South Florida. Ecotoxicology 2008, 17, 569. [Google Scholar] [CrossRef]
- US EPA. Method 365.1: Determination of Phosphorus by Semi-Automated Colorimetry; US Environmental Protection Agency: Cincinnati, OH, USA, 1993. [Google Scholar]
- Millard, S.P. EnvStats: An R Package for Environmental Statistics, 2nd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-8455-4. [Google Scholar]
- Vito, M.; Muggeo, R. Estimating Regression Models with Unknown Break-Points. Statist. Med. 2003, 22, 3055–3071. [Google Scholar] [CrossRef]
- Benedikt, G.; Prebsema, E.; Heuvelink, G. Spatio-Temporal Interpolation Using Gstat. R J. 2016, 8, 204–218. [Google Scholar]
- Fortin, M.-J. Spatial Statistics in Landscape Ecology. In Landscape Ecological Analysis; Springer: Berlin/Heidelberg, Germany, 1999; pp. 253–279. [Google Scholar]
- Journel, A.G. The Lognormal Approach to Predicting Local Distributions of Selective Mining Unit Grades. Math. Geol. 1980, 12, 285–303. [Google Scholar] [CrossRef]
- Yamamoto, J.K. On Unbiased Backtransform of Lognormal Kriging Estimates. Comput. Geosci. 2007, 11, 219–234. [Google Scholar] [CrossRef]
- South Florida Water Management District. Kissimmee River Vegetation Floodplain, 2008 Survey; South Florida Water Management District: West Palm Beach, FL, USA, 2010. [Google Scholar]
- Gohel, D. Flextable: Functions for Tabular Reporting 2021. Available online: https://cran.r-project.org/web/packages/flextable/index.html (accessed on 15 January 2021).
- Kingsford, R.T. Ecological Impacts of Dams, Water Diversions and River Management on Floodplain Wetlands in Australia. Austral. Ecol. 2000, 25, 109–127. [Google Scholar] [CrossRef]
- Noe, G.B.; Hupp, C.R.; Rybicki, N.B. Hydrogeomorphology Influences Soil Nitrogen and Phosphorus Mineralization in Floodplain Wetlands. Ecosystems 2013, 16, 75–94. [Google Scholar] [CrossRef]
- Van der Lee, G.E.M.; Olde Venterink, H.; Asselman, N.E.M. Nutrient Retention in Floodplains of the Rhine Distributaries in The Netherlands. River Res. Applic. 2004, 20, 315–325. [Google Scholar] [CrossRef]
- Noe, G.B.; Hupp, C.R. Carbon, Nitrogen, and Phosphorus Accumulation in Floodplains of Atlantic Coastal Plain Rivers, USA. Ecol. Appl. 2005, 15, 1178–1190. [Google Scholar] [CrossRef]
- Gordon, B.A.; Dorothy, O.; Lenhart, C.F. Nutrient Retention in Ecologically Functional Floodplains: A Review. Water 2020, 12, 2762. [Google Scholar] [CrossRef]
- Ward, J.V.; Tockner, K.; Schiemer, F. Biodiversity of Floodplain River Ecosystems: Ecotones and Connectivity. Regul. Rivers Res. Manag. 1999, 15, 125–139. [Google Scholar] [CrossRef]
- Toth, L.A.; Melvin, S.L.; Arrington, D.A.; Chamberlain, J. Hydrologic Manipulations of the Channelized Kissimmee River: Implications for Restoration. BioScience 1998, 48, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Toth, L.A. The Ecological Basis of the Kissimmee River Restoration Plan. Fla. Sci. 1993, 56, 25–51. [Google Scholar]
- Koebel, J.W.; Jones, B.L.; Arrington, D.A. Restoration of the Kissimmee River, Florida: Water Quality Impacts from Canal Backfilling. Environ. Monit. Assess. 1999, 57, 85–107. [Google Scholar] [CrossRef]
- Williams, V.P. Effects of Point-Source Removal on Lake Water Quality: A Case History of Lake Tohopekaliga, Florida. Lake Reserv. Manag. 2001, 17, 315–329. [Google Scholar] [CrossRef]
- Spencer, L.J.; Bousquin, S.G. Interim Responses of Floodplain Wetland Vegetation to Phase I of the Kissimmee River Restoration Project: Comparisons of Vegetation Maps from Five Periods in the River’s History. Restor. Ecol. 2014, 22, 397–408. [Google Scholar] [CrossRef]
- Harris, W. Mineral Distribution and Weathering in the Greater Everglades: Implications for Restoration. Crit. Rev. Environ. Sci. Technol. 2011, 41, 4–27. [Google Scholar] [CrossRef]
- Osborne, T.Z.; Newman, S.; Scheidt, D.J.; Kalla, P.I.; Bruland, G.L.; Cohen, M.J.; Scinto, L.J.; Ellis, L.R. Landscape Patterns of Significant Soil Nutrients and Contaminants in the Greater Everglades Ecosystem: Past, Present, and Future. Crit. Rev. Environ. Sci. Technol. 2011, 41, 121–148. [Google Scholar] [CrossRef]
- Steiger, J.; Gurnell, A.M. Spatial Hydrogeomorphological Influences on Sediment and Nutrient Deposition in Riparian Zones: Observations from the Garonne River, France. Geomorphology 2003, 49, 1–23. [Google Scholar] [CrossRef]
- Steiger, J.; Gurnell, A.M.; Petts, G.E. Sediment Deposition along the Channel Margins of a Reach of the Middle River Severn, UK. Regul. Rivers: Res. Mgmt. 2001, 17, 443–460. [Google Scholar] [CrossRef]
- Reddy, K.R.; Osborne, T.Z.; Inglett, K.S.; Corstanje, R. Influence of Water Levels on Subsidence of Organic Soils in the Upper St. Johns River Basin; Saint Johns River Water Management District: Palatka, FL, USA, 2006. [Google Scholar]
- Bruland, G.L.; Osborne, T.Z.; Reddy, K.R.; Grunwald, S.; Newman, S.; DeBusk, W.F. Recent Changes in Soil Total Phosphorus in the Everglades: Water Conservation Area 3. Environ. Monit. Assess. 2007, 129, 379–395. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.R.; Newman, S.; Osborne, T.Z.; White, J.R.; Fitz, H.C. Phosphorous Cycling in the Greater Everglades Ecosystem: Legacy Phosphorous Implications for Management and Restoration. Crit. Rev. Environ. Sci. Technol. 2011, 41, 149–186. [Google Scholar] [CrossRef]
- Warne, A.G.; Toth, L.A.; White, W.A. Drainage-Basin–Scale Geomorphic Analysis to Determine Reference Conditions for Ecologic Restoration—Kissimmee River, Florida. GSA Bull. 2000, 112, 884–899. [Google Scholar] [CrossRef]
- Shrestha, J.; Niklaus, P.A.; Frossard, E.; Samaritani, E.; Huber, B.; Barnard, R.L.; Schleppi, P.; Tockner, K.; Luster, J. Soil Nitrogen Dynamics in a River Floodplain Mosaic. J. Environ. Qual. 2012, 41, 2033–2045. [Google Scholar] [CrossRef] [PubMed]
- Clément, J.-C.; Holmes, R.M.; Peterson, B.J.; Pinay, G. Isotopic Investigation of Denitrification in a Riparian Ecosystem in Western France. J. Appl. Ecol. 2003, 40, 1035–1048. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Period | Major Action/Event |
|---|---|
| 1954 | United States Congress authorizes the C&SF Project |
| 1962 | Construction commences of the C38 Canal and other features |
| 1971 | Construction complete |
| 1971 | The state of Florida recommends the restoration of the Kissimmee River |
| 1978–1985 | Federal feasibility study notes the potential for restoration [20] |
| 1984–1990 | Kissimmee River Demonstration Project |
| 1987–1990 | Drawdown of upstream lakes (Lake Tohopekaliga and East Lake Tohopekaliga) [21] |
| 1991 | 2nd Federal feasibility study recommends a partial backfill plan [22] |
| 1994 | Construction of test backfill and performance of high-flow tests on backfill stability |
| 1995–1997 | Drawdown of upstream lakes (Lake Jackson and Lake Kissimmee) [21] |
| 1996 | Headwaters Revitalization Feasibility Study complete [23] |
| 1999–2001 | Phase I backfilling complete (12 km canal backfilled) |
| 2001 | S-65B water control structure removed |
| 2006–2010 | Phase IV backfilling complete (9 km canal backfilled) |
| 2010–2015 | Supplemental construction to support the Kissimmee River Restoration effort [24] |
| 2015–2021 | Phases II and III backfilling complete (~14 km canal backfilled) |
| 2017 | S-65C water control structure removed |
| 2021 | Kissimmee River Restoration Construction Activities complete |
| Variable | Parameter | Statistic | S65 | S65A | S65B | S65C | S65D | S65E |
|---|---|---|---|---|---|---|---|---|
| Load | TP (kg yr−1) | Mean | 56,499 | 62,577 | 53,038 | 74,270 | 118,647 | 138,426 |
| Min | 834 | 3980 | 8 | 4040 | 8597 | 10,436 | ||
| Max | 211,233 | 165,613 | 166,524 | 203,444 | 383,061 | 348,449 | ||
| SD | 45,451 | 41,589 | 39,070 | 49,958 | 79,803 | 82,252 | ||
| N | 44 | 44 | 23 | 39 | 44 | 44 | ||
| TN (kg yr−1) | Mean | 1,238,417 | 1,277,163 | 1,289,658 | 1,375,850 | 1,625,018 | 1,685,533 | |
| Min | 26,218 | 62,151 | 138 | 111,078 | 126,163 | 112,253 | ||
| Max | 3,515,506 | 3,044,856 | 3,488,465 | 3,422,184 | 3,922,393 | 4,125,750 | ||
| SD | 789,083 | 736,539 | 912,158 | 840,922 | 968,248 | 994,486 | ||
| N | 44 | 44 | 23 | 39 | 44 | 44 | ||
| FWM | TP (μg L−1) | Mean | 58.0 | 64.5 | 55.7 | 69.5 | 94.6 | 111.9 |
| Min | 30.7 | 34.3 | 31.8 | 34.7 | 57.4 | 65.5 | ||
| Max | 94.3 | 134.8 | 92.2 | 162.9 | 201.2 | 228.0 | ||
| SD | 17.6 | 19.8 | 14.7 | 25.9 | 29.9 | 36.2 | ||
| N | 44 | 44 | 23 | 39 | 44 | 44 | ||
| TN (mg L−1) | Mean | 1.35 | 1.32 | 1.35 | 1.29 | 1.31 | 1.32 | |
| Min | 0.81 | 0.87 | 0.95 | 0.88 | 0.99 | 0.97 | ||
| Max | 2.19 | 1.90 | 1.95 | 1.97 | 1.96 | 2.02 | ||
| SD | 0.26 | 0.21 | 0.30 | 0.26 | 0.23 | 0.22 | ||
| N | 44 | 44 | 23 | 39 | 44 | 44 |
| Variable | Parameter | Statistic | S65 | S65A | S65B | S65C | S65D | S65E |
|---|---|---|---|---|---|---|---|---|
| Load | TP | Kendall τ | 0.28 | 0.29 | 0.12 | 0.30 | 0.26 | 0.16 |
| ρ-value | <0.01 | <0.01 | 0.43 | <0.01 | <0.05 | 0.12 | ||
| Thiel Sen Slope A | 1058 | 1206 | 1189 | 2027 | 2237 | 1604 | ||
| TN | Kendall τ | 0.11 | 0.11 | −0.11 | 0.10 | 0.14 | 0.06 | |
| ρ-value | 0.32 | 0.32 | 0.46 | 0.36 | 0.20 | 0.55 | ||
| Thiel Sen Slope A | 10341 | 9419 | −27066 | 11055 | 14240 | 7913 | ||
| FWM | TP | Kendall τ | 0.41 | 0.33 | 0.30 | 0.39 | 0.18 | 0.03 |
| ρ-value | <0.01 | <0.01 | 0.05 | <0.01 | 0.08 | 0.78 | ||
| Thiel Sen Slope B | 0.72 | 0.60 | 0.90 | 0.93 | 0.50 | 0.09 | ||
| TN | Kendall τ | −0.10 | −0.17 | −0.63 | −0.22 | −0.19 | −0.19 | |
| ρ-value | 0.35 | 0.11 | <0.01 | 0.05 | 0.06 | 0.07 | ||
| Thiel Sen Slope C | −0.003 | −0.004 | −0.027 1pt | −0.008 | −0.005 | −0.005 |
| Mean ± SD (N) | ||||||
|---|---|---|---|---|---|---|
| Units | Classification | Bulk Density (g cm−3) | LOI (Percent) | TP (mg kg−1;) | TN (g kg−1) | TC (g kg−1) |
| Vegetation Community | Aquatic Veg | 0.57 ± 0.36 (31) | 29.3 ± 24.0 (35) | 625 ± 448 (35) | 10.7 ± 8.7 (35) | 140.1 ± 120.2 (35) |
| Wet Prairie | 0.78 ± 0.34 (54) | 17.0 ± 18.5 (58) | 514 ± 558 (58) | 5.9 ± 6.3 (58) | 79.3 ± 93.1 (58) | |
| Broadleaf Marsh | 0.52 ± 0.31 (25) | 33.6 ± 29.0 (29) | 705 ± 531 (29) | 11.7 ± 10.1 (29) | 159.0 ± 141.5 (29) | |
| Upland Shrub | 0.93 ± 0.30 (51) | 11.3 ± 11.3 (58) | 533 ± 710 (58) | 3.7 ± 3.7 (58) | 52.1 ± 56.3 (58) | |
| Wetland Forest/Shrub | 0.52 ± 0.33 (42) | 28.6 ± 25.1 (43) | 591 ± 561 (43) | 9.4 ± 8.1 (43) | 136.2 ± 124.0 (43) | |
| Upland Forest | 0.91 ± 0.28 (28) | 10.4 ± 8.8 (32) | 480 ± 802 (32) | 2.9 ± 2.4 (32) | 49.4 ± 45.3 (32) | |
| Landscape Unit | Channel | 0.48 ± 0.31 (17) | 29.0 ± 28.7 (17) | 521 ± 449 (17) | 10.3 ± 9.8 (17) | 142.6 ± 145.3 (17) |
| Backfill | 0.90 ± 0.34 (13) | 8.4 ± 7.2 (13) | 1003 ± 496 (13) | 2.4 ± 2.5 (13) | 34.9 ± 31.8 (13) | |
| Floodplain | 0.63 ± 0.35 (102) | 26.4 ± 24.4 (113) | 470 ± 379 (113) | 9.1 ± 8.1 (113) | 124.7 ± 119.3 (113) | |
| Upland Ecotone | 0.90 ± 0.30 (38) | 10.3 ± 10.7 (42) | 187 ± 243 (42) | 3.4 ± 3.6 (42) | 51.5 ± 57.4 (42) | |
| Spoil | 0.94 ± 0.28 (34) | 10.3 ± 9.9 (37) | 1183 ± 1089 (37) | 2.9 ± 3.0 (37) | 43.1 ± 42.8 (37) | |
| Other | 0.66 ± 0.37 (27) | 24.1 ± 20.9 (33) | 521 ± 369 (33) | 8.6 ± 7.6 (33) | 116.5 ± 106.4 (33) | |
| Kriging Method | Parameter | Model | Nugget | Sill | Range (m) | Sill:Nugget | RMSE | R² |
|---|---|---|---|---|---|---|---|---|
| Ordinary | TP | Exp | 0.99 | 0.55 | 2973 | 0.55 | 0.005 | 1.00 |
| TN | Sph | 17.8 | 40 | 1035 | 2.22 | 1.8 | 0.96 | |
| TC | Sph | 4175 | 7733 | 1079 | 1.85 | 23.9 | 1.00 | |
| Residual | TP | Sph | 0.43 | 0.18 | 5306 | 0.42 | 0.001 | 1.00 |
| TN | Exp | 0.39 | 0.32 | 739 | 0.83 | 0.007 | 0.99 | |
| TC | Sph | 0.44 | 0.41 | 1292 | 0.93 | 0.030 | 0.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Julian, P., II; Osborne, T.Z.; Ellis, R. Evaluation of Biogeochemical Changes in Channelized and Restored Portions of a Subtropical Floodplain. Hydrobiology 2023, 2, 1-18. https://doi.org/10.3390/hydrobiology2010001
Julian P II, Osborne TZ, Ellis R. Evaluation of Biogeochemical Changes in Channelized and Restored Portions of a Subtropical Floodplain. Hydrobiology. 2023; 2(1):1-18. https://doi.org/10.3390/hydrobiology2010001
Chicago/Turabian StyleJulian, Paul, II, Todd Z. Osborne, and Rex Ellis. 2023. "Evaluation of Biogeochemical Changes in Channelized and Restored Portions of a Subtropical Floodplain" Hydrobiology 2, no. 1: 1-18. https://doi.org/10.3390/hydrobiology2010001