1. Introduction
The phylum Cnidaria includes sessile animals such as sea anemones and corals in the subphylum Anthozoa, unusual parasitic forms known as myxozoans [
1], and taxa including free-swimming jellyfish in the subphylum Medusozoa [
2]. Medusozoans are strikingly diverse, ranging from colonial siphonophores and hydroids to solitary polyps and medusae, and often have complex life cycles linking divergent forms, and thus have undergone a long history of taxonomic revision [
3,
4,
5,
6]. Within Medusozoa is the lesser-known class Staurozoa, or stalked jellyfish, a group of benthic cnidarians that includes about 50 known species [
7]. Oddly, it was not until the discovery of the gastric cirri in stauromedusae (the sexually reproductive life stage of staurozoans) that scientists first associated these organisms with jellyfishes (reviewed in [
6]. Early phylogenetic studies revealed the monophyly of the class Staurozoa within Medusozoa [
2,
8], but an insufficient grasp of suitable morphological characters for delineating these taxa rendered the systematic relationships among the species poorly understood. More recently, a comprehensive morphological and molecular analysis of a large number of staurozoans provided a phylogenetic revision with two suborders comprising the class: Myostaurida, taxa possessing interradial longitudinal muscles in the peduncle, and Amyostaurida, those lacking such muscles [
6,
9].
Like all cnidarians, staurozoans have nematocysts (stinging capsules used for prey-capture, adhering to substrates, and protection), are radially symmetrical, can reproduce sexually and asexually, and have two body layers (endoderm and ectoderm) with a central gelatinous layer (mesoglea). Staurozoans typically inhabit temperate and boreal waters; however, some reports exist from the tropics (reviewed in Miranda et al. 2018). Stauromedusae are usually found attached to substrates such as macroalgae, seagrass, and rocks in shallow waters, but occur even in deep-sea hydrothermal vents where they are found in dense monospecific populations (reviewed in Mills and Hirano 2007; Miranda et al. 2018). However, little is known about staurozoan substrate preference during early development. Furthermore, their small size (several millimeters to centimeters in length), color that often matches the substrate, and frilly calyx, make them difficult to detect on sea algae and seagrass [
7], hence the challenging nature of field studies on stalked jellyfish (reviewed in [
7,
10]). Major knowledge gaps exist surrounding staurozoan sexual reproduction and early life history. The absence of a pelagic medusa in the life cycle (i.e., maturation in the absence of fission or budding) makes staurozoans unique among non-hydrozoan medusozoans.
A review [
11] showed that, of nine species comprising five genera (including three
Haliclystus species), peak abundance varies in the wild, but an annual life cycle is thought to be a common trait, with some exceptions. Curiously, frequent reports of disappearance of stauromedusae for several months prior to discovery of juvenile stages in the field suggests that larvae enter a dormant stage (cysts) until conditions are right for completing the life cycle (reviewed in [
11]). For example, it was previously perceived that the gradual disappearance of stauromedusae coincided with seasonal death of
Sargassum (brown algae) at different regions across Japan [
12], and that the life cycle of
Haliclystus inabai (as
Stenoscyphus inabai) was concurrent with the life cycle of the brown alga
Cystophora expansa at Black Rock, Australia [
13]. Currently, environmental factors contributing to synchronized spermcasting events or triggering early development are not understood, although multiple studies have indicated light may be an important cue, e.g., [
14,
15]. Individual abiotic requirements (e.g., temperature) of each species, or relationship to the life cycle of their preferred substrate (e.g., algae or bivalves), are not understood either.
An early work provides an extensive account of the life cycle of the staurozoan
Haliclystus octoradiatus based on live rearing in the lab [
15]. Ripe
H. octoradiatus individuals were collected in Roscoff (France) during all seasons, with the fewest present in the spring and the most in the summer. These life cycle studies on
H. octoradiatus mention a “settled larva” form with a hemispherical body covered in nematocysts that gives rise laterally to motile frustules (frustulation), which then settle and, like the settled larva from which they are derived, go through several development stages towards becoming stauropolyps. In recent years, the “settled larva” stage has been dubbed the “microhydrula” stage after the hemispherically shaped
Microhydrula limopsicola, originally described as a species of Hydrozoa, was determined genetically to be the settled larvae of
Haliclystus antarcticus [
16]. Through asexual reproduction, the microhydrula stage would permit proliferation in favorable environments; however, thus far, the microhydrula stage has been found in only two species of
Haliclystus (the most species-rich staurozoan genus). While speculation exists that this early development stage may be a synapomorphy of the genus
Haliclystus [
7], for most of the 11 Staurozoa genera currently recognized, little is known of their life cycles.
In this study, we focus on two staurozoan morphotypes discovered in Kitsunezaki (Ishinomaki City, Miyagi, Japan)—an area that was devastated by the 2011 Great East Japan Earthquake and Tsunami [
17]. We analyzed the respective morphologies, cnidomes (repertoire of nematocyst stinging capsules), and degree of gonad conspicuousness of individuals collected over a non-continuous 18-month period. Phylogenetic analyses using two mitochondrial molecular markers (16S rRNA gene and COI) clarified the Kitsunezaki specimens as representing three co-occurring species, two conforming to the description of
Haliclystus inabai (Myostaurida) and one to
Calvadosia nagatensis (Amyostaurida). Analysis of our additional substantial sequence data revealed that
H. inabai comprises two distinct species—one including only samples from Japanese localities (
H. inabai lineage 1), and the other including samples from Japan and several continents (
H. inabai lineage 2), either from nearshore waters or found in aquariums. This study underlines the importance of collaboration between academia and aquariums. Scientists in both settings can produce research findings of reciprocal relevance, and collaborations can take advantage of complementary expertise, e.g., systematics and live jellyfish husbandry, to make more rapid progress in advancing our understanding of marine biodiversity.
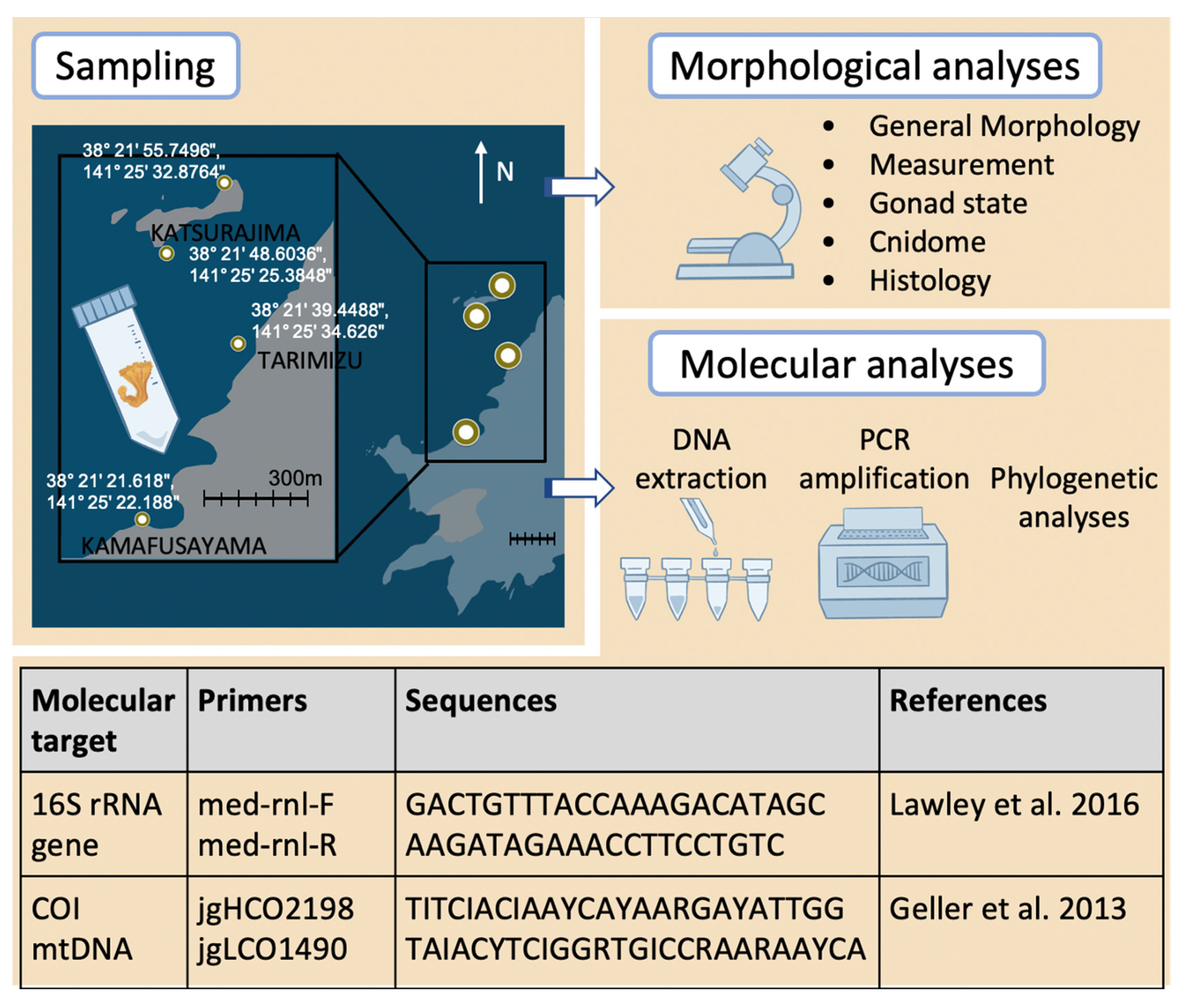
2. Materials and Methods
Stauromedusae of two distinct morphotypes were collected (
Figure 1) at the study site in Kitsunezaki, Ishinomaki, Oshika Peninsula, Miyagi Prefecture (38°21′ N, 141°25′ E, and vicinity—details in
Figure 2). This location on Honshu was hit directly by the Great East Japan Earthquake on 11 March 2011. Following recovery efforts, an active survey site for research on seaweeds and seagrasses was established in July of the same year (for sampling site details see [
18,
19]).
Eisenia bicyclis kelp (
Figure 1A) and
Gelidium (
Figure 1B) grow abundantly on inshore (0–30 m) rocky bottoms (1–4 m depth), while the seagrass
Zostera caespitosa dominates the offshore (30–70 m) sandy bottoms (4–6 m depth).
Prior to this study, the two distinct stauromedusae (
Figure 1B–F) were first collected from the location in 2018, by one of the co-authors (M.A.) and, subsequently, this study began one year later in October 2019. Seawater temperature was measured using HOBO data logger (
Supplementary File S1) and dive computer (ScubaPro M2). The stauromedusae were collected by hand while SCUBA diving, then placed into 50 mL Falcon tubes filled with seawater (
Figure 2) along with a small piece of the macroalgal substrate, before being transported live in an insulated cooler to the laboratory of Marine Plant Ecology, Graduate School of Agricultural Science, Faculty of Agriculture, Tohoku University.
Out of the total specimens collected at Kitsunezaki (ca. 100), a portion was preserved immediately after collection in pure ethanol (99.5%), and then stored at −30 °C for future molecular analysis; some were fixed in 70% ethanol or 5–8% formalin solution for morphological study. Several stauromedusae (ca. 25) were kept alive for observations and fed three times a week on Artemia nauplii. However, rearing stauromedusae collected in the wild using artificial seawater proved to be challenging, and most died within a few days, with only a handful living for up to 79 days in simple aquariums.
In addition to the Tohoku specimens obtained at the survey site, additional H. inabai specimens that had unexpectedly appeared in aquariums in Japan (Kamo Aquarium and Kagoshima Aquarium) and Canada (Aquarium du Québec) were obtained for morphological and molecular analyses. Life history observations were made in the Aquarium du Québec.
2.1. H. inabai Stauromedusae Appearing in Aquariums
In the Aquarium du Québec, several
Haliclystus inabai stauromedusae were first discovered on 16 April 2020 in an exhibit tank of 1016 L in volume (totaling 2600 L including life support and sump). Within the exhibition tank, with a sand substrate, were potbelly seahorses and other decor including dead coral rocks or plastic ones. Both adult and juvenile
H. inabai stauromedusae as well as early life stages (discussed below) were found on artificial plants that were frequently cleaned, including Eelgrass Zostera (bio models, Aguanga, CA, USA) as well as some uncategorized types, and Green Cardamine (resin ornament, PetSave Direct, Woodbridge, ON, Canada), the latter of which hosted mainly early life stages on one particularly clean leaf. Following the discovery, additional stauromedusae were noted in a smaller quarantine tank (182 L) also containing potbelly seahorses and plastic plant decor (
Figure 3A). The temperature in both artificial seawater tanks (Instant Ocean, 32 g/L, potassium iodide 0.06 to 0.08 ppm), was kept at about 16 °C. Over a two-month period (28 November 2020–1 February 2021) a total of 31 stauromedusae (approximately 12.5 mm; range 11–15 mm in length) were counted in the smaller tank attached to coral rocks, shells, air stone weights, and plastic plants (
Figure 3B,C), and one individual was found attached to a seahorse. Though easily removed from the latter with seawater forced using a pipette, it was difficult to detach the stauromedusae from harder surfaces, such as aquarium walls and coral rocks. Plastic leaves bearing staurozoans were transferred to a new tank in a sea-table (6 L plastic tank HerpHeaven, Brunswick, ME, USA), with a small pump for water circulation and air tubing for aeration (gentle water flow; fluctuating 16–19 °C), in order to conduct observations on feeding and life history. Staurozoans were fed on nauplii (48 h after hatching, 550 μm in length) of
Artemia brine shrimp (SELCO enriched, Artemia International, Fairview, TX, USA). Juvenile stauromedusae were typically beige-brown with a hint of pink (possibly due to ingested brine shrimp), while adults were brown with a white “snowflake” pattern along the length, or took on a green hue (
Figure 3D–F). Photo and video documentation was made via Samsung Galaxy Smartphone. Six of these individuals were preserved in 95% ethanol and shipped to Tohoku University on 20 January 2021 for molecular analysis.
Similarly, Haliclystus stauromedusae were also discovered at Kamo Aquarium (Tsuruoka, Japan), appearing in reserve aquarium tanks (144 L, with a 100 L filtration tank—mesh sizes of 10 μm, then 5 μm, 15 °C) and Kagoshima Aquarium (Kagoshima, Japan). Individuals were sent to Tohoku University, both living and preserved in 99.5% ethanol, for further analyses.
2.2. Morphology, Microscopy
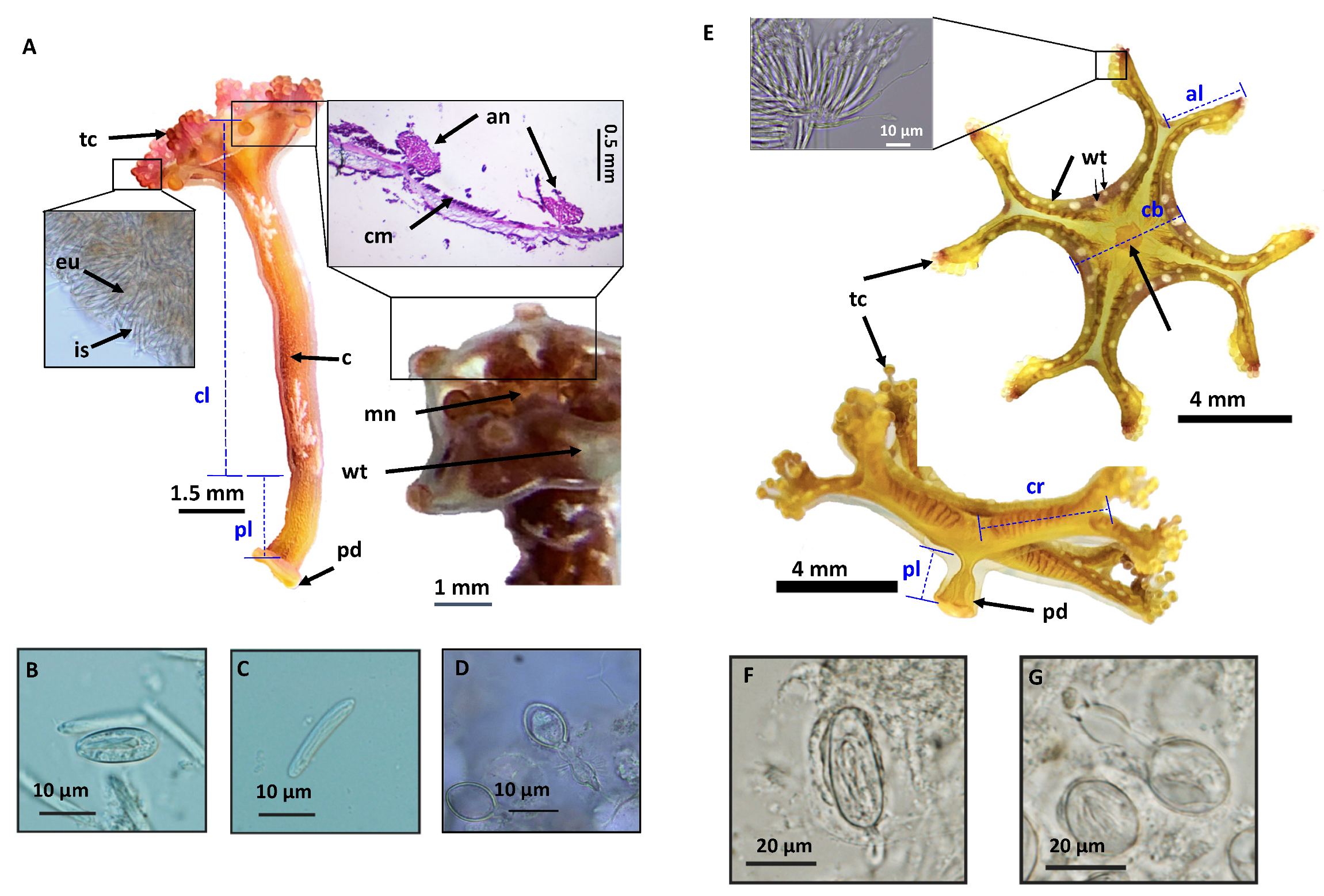
Specimens collected from Kitsunezaki and preserved in 70% ethanol or 5–8% formalin were observed under a dissecting microscope (Olympus SZX12), imaged with a digital camera (Olympus DP72), and measured (CellsSens software version 1.8, Olympus, Tokyo, Japan). The presence or absence of gonads was confirmed, and the cnidome (the repertoire of stinging capsules, called nematocysts) was characterized using compound microscopy (Olympus BX61 or Nikon Eclipse Ni-U and camera) (
Figure 2).
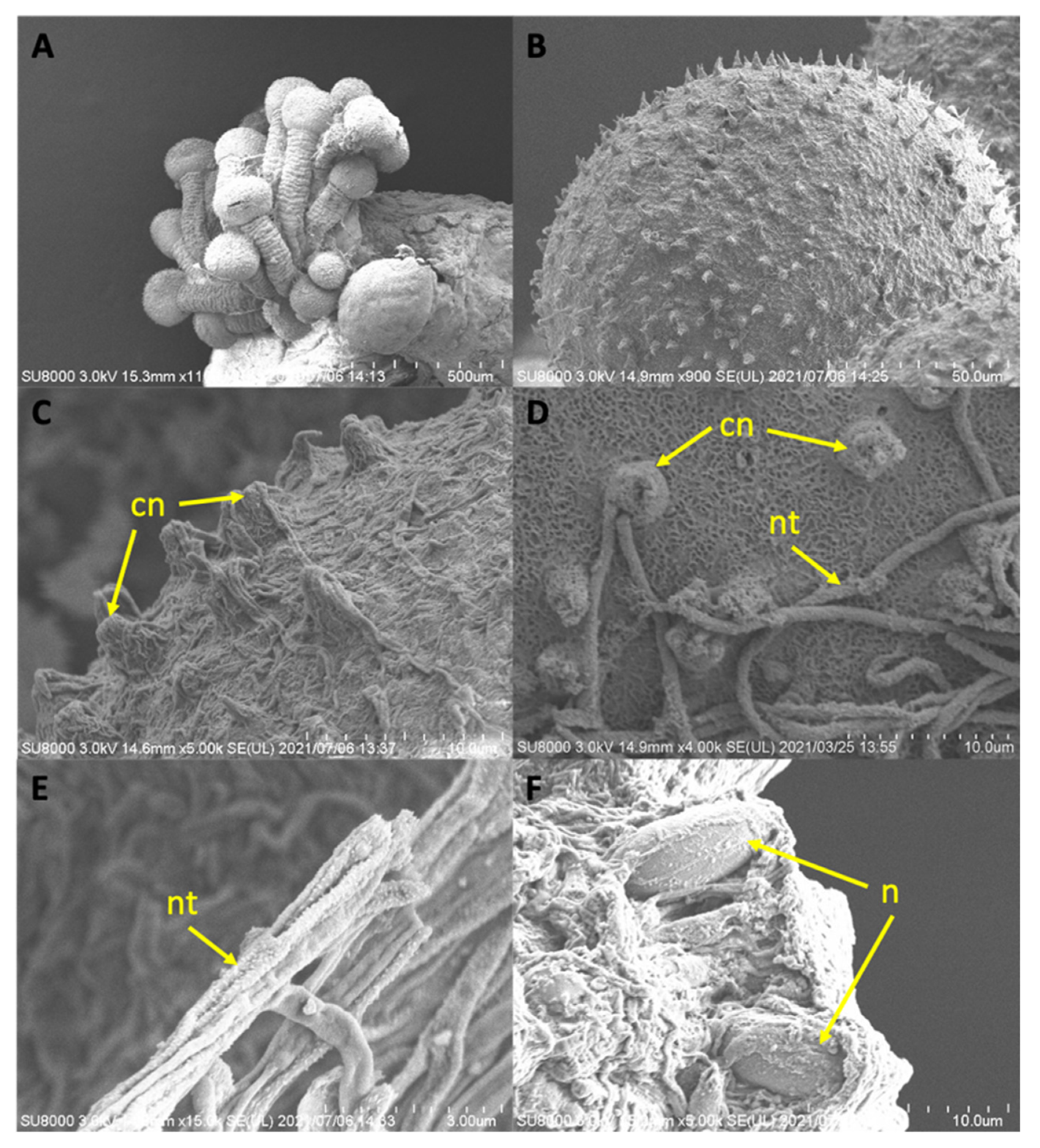
The ultrastructure of finer features was documented with Scanning Electron Microscopy (SEM) (Hitachi SU8000 (Type II)) at the Electron Microscopy Section in Tohoku University Graduate School of Agricultural Science. For SEM preparation, samples previously preserved in 70% ethanol were subjected to a graded rehydrated series (70–25%, 5–10 minutes each) of 1X PBS (Phosphate Buffered Saline, pH 7.4); fixed in isosmotically adjusted PBS buffer glutaraldehyde solution (2.5%) overnight at 4 °C; washed with isosmotic 1X PBS buffer and milli-Q water twice to remove salts; and subjected to a graded dehydration series of ethanol (25–100%, 5–10 minutes each). Dried samples were mounted onto an SEM stub (13 mm) using an adhesive carbon strip, and subjected to critical point dry, and coated in gold/palladium (MSP-1S sputtering device) before observation with SEM.
General histological procedures [
22] were carried out on staurozoans preserved in 8% formalin. Specimens were dehydrated in a series of graded ethanol (70–100%); cleared in Lemosol (2 steps), infiltrated and embedded in paraffin; serially sectioned transversely (8.0–10.0 µm thick); washed with distilled water, and stained using hematoxylin-eosin. Prepared slides were photographed with light microscopy.
2.3. Molecular Barcoding
DNA was extracted from Staurozoa samples using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, NW, Germany), and total DNA yield was quantified (ng/μL) using a fluorometer (QuantiFluor, Promega, Tokyo, Japan). The targets for DNA amplification were two mitochondrial gene fragments: 16S rRNA gene (565 bp) and cytochrome oxidase subunit 1 (COI) gene (720 bp). Each reaction (50 μL) contained 25 μL
Taq PCR Master Mix (Qiagen), template DNA (~200 ng), nuclease free water, and 2 μL each of forward and reverse primers (10 μM;
Figure 2).
PCR (polymerase chain reaction) was used to amplify the two molecular targets in a thermal cycler (BentoLab, Bioworks, London, UK or Takara Bio, Kusatsu, Shiga, Japan). Cycling profile for 16S primers included initial denaturation at 94 °C for 3 minutes, followed by 38 cycles of denaturation, annealing and extension (94 °C for 30 s, 54 °C for 30 s, and 72 °C for 45 s), and a final extension at 72 °C for 10 minutes [
23]. Cycling profile for COI primer included denaturation at 94 °C for 5 minutes, followed by 38 cycles of denaturation, annealing, and extension (94 °C for 30 s, 46 °C for 30 s, and 72 °C for 1 min), and a final extension at 72 °C for 10 minutes. PCR products were visualized on a 1% agarose gel using gel electrophoresis and visualized with a gel illuminator and nucleic acid gel stains (GelGreen or GelRed). Amplicons were purified with ExoSap-IT (Applied Biosystems, Tokyo, Japan) PCR Product Cleanup toolkit prior to Sanger sequencing (Eurofins or Genewiz, Tokyo, Japan).
Generated 16S rRNA and COI gene sequences were edited in Geneious Prime (v.2022.0.1,
www.geneious.com: accessed on 11 November 2021). Additional Staurozoa sequences were acquired from GenBank (
https://www.ncbi.nlm.nih.gov/genbank/: accessed on 11 November 2021).
Depastromorpha africana and
Craterolophus convolvulus were used as outgroups in the analyses of
Haliclystus and
Calvadosia species, respectively. Sequences were aligned in MAFFT using the E-INS-i and G-INS-i algorithms, respectively, for 16S and COI. Maximum likelihood trees, with nodes assessed by 400 bootstrap replicates, were constructed using PHYML as implemented in Geneious Prime (v.2022.0.1) and assuming the GTR model of nucleotide substitution. Sequences generated in this study were accessioned into GenBank, and morphological vouchers were deposited as museum specimens (
Table 1). Alignments and resulting topologies are provided as
Supplementary Files S2–S9.
4. Discussion
4.1. Staurozoan Identification and Cnidome
This study employed molecular barcoding, morphological analyses, light microscopy and SEM, and collaborations with aquariums in Japan and Canada, to identify three species of Staurozoa reported herein for the first time in Kitsunezaki—two as Haliclystus inabai (Kishinouye, 1893) and one as Calvadosia nagatensis (Oka, 1897), of the respective suborders Myostaurida and Amyostaurida.
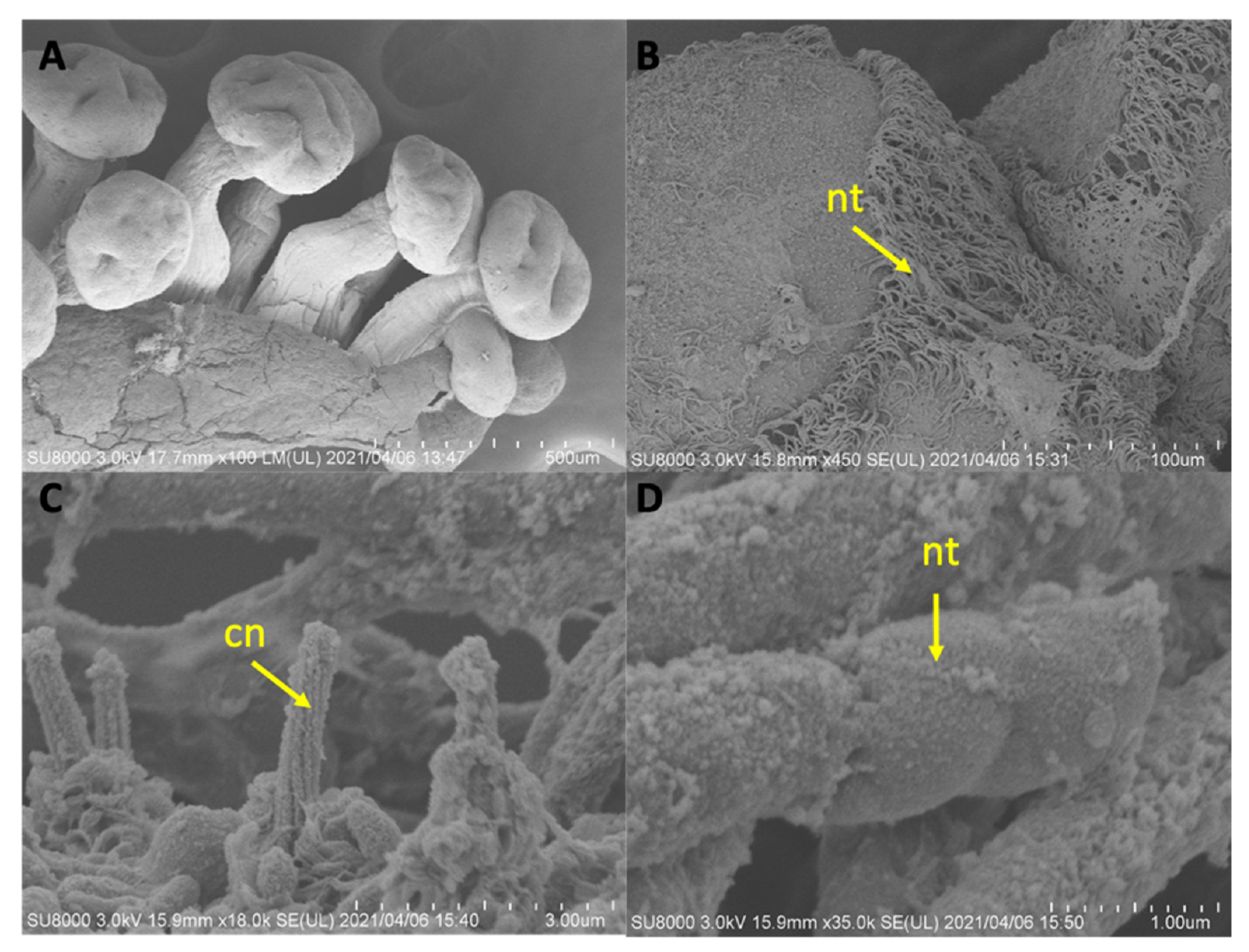
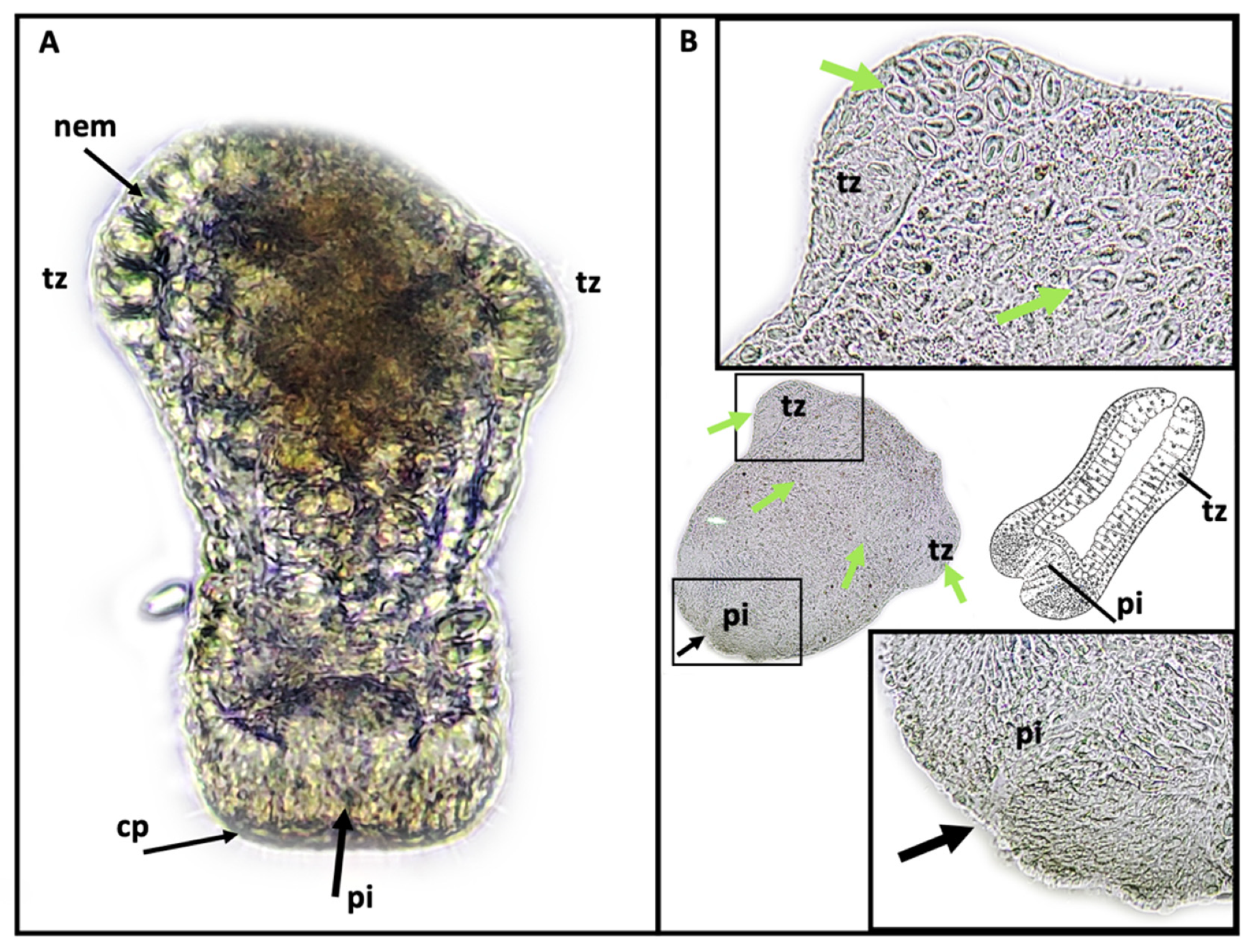
The use of SEM in this study to conduct ultrastructural observations of secondary tentacles of both staurozoan species provided a clear picture of the structure of the bilaterally symmetrical mechanosensory cnidocil complex (apparatus) at the apex of each nematocyst capsule, which consists of central cilium surrounded by a crescent of stereocilia. This sensory component of the nematocyst controls the firing of the nematocysts, and is vital for nematocyst discharge. Though well-studied in several hydrozoan species, e.g.,
Hydra [
36] and
Craspedacusta sowerbii [
29], only a single mention exists of the cnidocil complex in a Staurozoa (James-Clark 1878); the structure is documented herein for the first time in the class with SEM. More detailed observations using TEM techniques are required to verify whether the ciliary pattern of the cnidocil apparatus follows the same as that seen in hydrozoans.
4.2. Molecular Phylogenetics
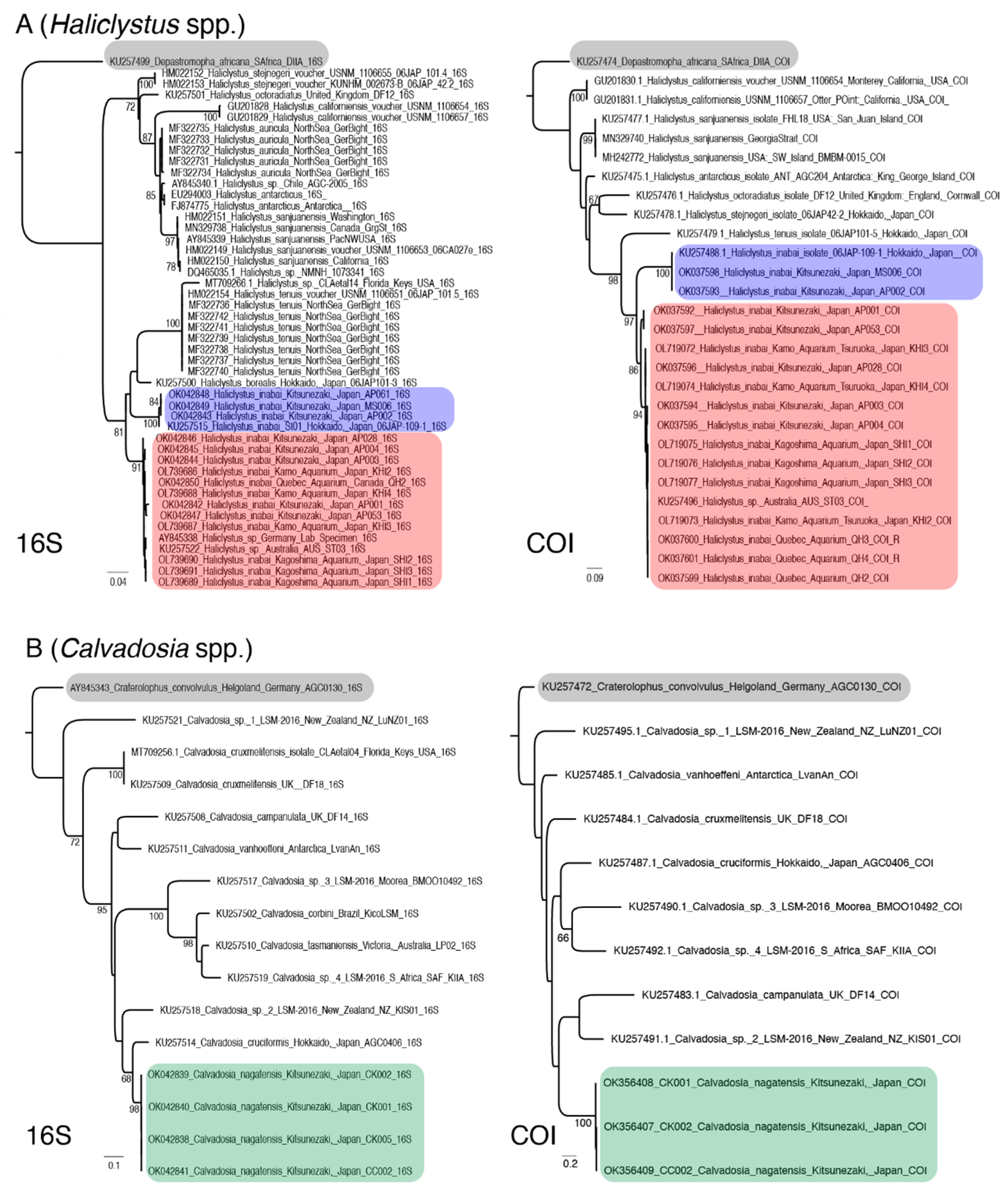
Our molecular analyses revealed that H. inabai consists of two distinct lineages: Haliclystus inabai lineage 1 is so far known only from Japan, comprised exclusively of specimens collected in the wild (Kitsunezaki and Hokkaido); H. inabai lineage 2 is more broadly distributed, with specimens collected from Kitsunezaki (Japan), Victoria (Australia), aquariums in Québec (Canada), Tsuruoka (Japan), Kagoshima (Japan), and a university lab (Germany).
Despite their broad average genetic differences, no apparent morphological distinction was detected, suggesting that the two clades represent two distinct but cryptic species. More extensive sampling and study of both lineages could permit morphological distinction of the two species, determination of which lineage represents the true H. inabai, and ultimately a formal naming of the new species.
Our molecular analysis identified a sequence labeled as
Haliclystus sp. from a lab in Germany (AY845338.1) clustering with the
H. inabai sequences, revealing a case of clarified identity, as this sequence was originally deposited as
Haliclystus sp. (
Figure 8A). Furthermore,
Haliclystus sp. (KU257522, KU257496) from Victoria (Australia, in the Southern Hemisphere) was previously hypothesized to be a separate species from the Hokkaido one (Northern Hemisphere) based on molecular data for an individual from each population (Miranda et al. 2016b). However, increased sample analyses in this study, for both gene markers, revealed that the Australian
Haliclystus resembling
H. inabai is also present in Japan, and appearing spontaneously in aquariums in other parts of the world where the samples’ origins are unknown (
Figure 8A,B).
The exchange of live aquatic samples, which is part-and-parcel to scientific collaboration between academic institutions and aquariums, may have played a role in the distribution of
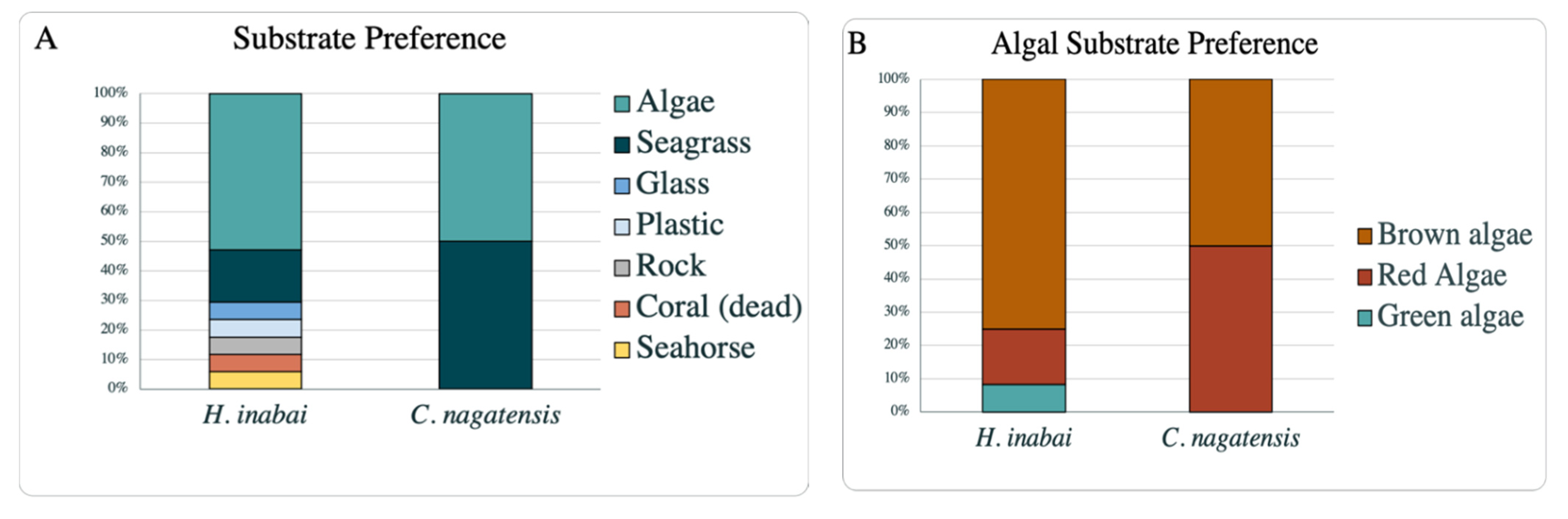
H. inabai beyond the walls of aquariums on different continents. In addition to observations on substrate preference in this study, the survey of literature revealed
H. inabai to have a broad substrate preference, consistent with the records of this species from multiple countries. Furthermore, in all cases in this study where
H. inabai appeared in aquariums, no stauromedusae had been placed in those aquarium tanks, suggesting
H. inabai was likely introduced in the form of one of the microscopic early life cycle stages attached to a substrate (i.e., synthetic plants, coral heads, or even a live seahorse). How staurozoans are distributed across oceans is still a mystery, as no studies on staurozoan mobility or recruitment exist. Although speculation exists about the potential for rafting and ocean currents to aid in distribution of staurozoans across oceans, an inferred dispersal mode for over 100 non-staurozoan medusozoans [
37] to date, no reports exist that test that hypothesis. Recently, eDNA (environmental DNA) signatures of two staurozoan species,
Calvadosia cruxmelitensis and
Haliclystus cf.
tenuis, were detected in the Florida Keys (Key Largo and Fleming Key, FL, USA) [
23], where they had not been reported previously, suggesting that these cryptic animals may have more widespread distributions than previously appreciated. Novel approaches to eDNA detection (e.g., [
38]) should aid our understanding of global distribution paths of staurozoans and their underlying mechanisms of dispersal.
This study is the first to accession the molecular markers 16S rRNA and COI of
C. nagatensis to GenBank, showing the limited resources available for this species, whose more narrow distribution range is consistent with the dearth of records on substrate preference of this species (as discussed in the previous section). While
C. nagatensis appears to have less adaptive potential to different environments and substrates compared to
H. inabai, population declines or extirpations are purported to be connected to pollution or loss of algal/seagrass substrates [
7], but additional studies are needed to better understand potential threats to staurozoan species stemming from human activities.
4.3. Distribution and Seasonal Variation
During our 18-month field and lab study, we gained a better understanding of the two Staurozoa species documented for the first time at the Kitsunezaki survey site. However, due to unforeseen circumstances related to weather and the impact of the COVID-19 pandemic countermeasures (reviewed in Abramo et al. 2022 [
39]), sporadic sampling impeded diligent sampling efforts and our ability to document sexual reproduction or early life stages of either species in the field.
In this study, we collected stauromedusae of both
H. inabai (both lineages) and
C. nagatensis on
Eisenia bicyclis kelp and
Gelidium seaweed, but no early life stages were witnessed in the field. Differential use of substrates during ontogeny has been documented in staturozans. Wietrzykowski (1912) reported that
H. octoradiatus adult stauromedusae were mainly found on
Zostera eelgrass, while early life stages were exclusively found on filamentous
Ceramium red algae; the interstitial staurozoan
Stylocoronella riedli, on the other hand, settles on coarse sand mixed with shell during early development [
35], while its stauromedusa stage has been found on a sea cucumber [
40]. Additionally, the microhydrula stage of
Haliclystus antarcticus was discovered upon the shell of lamellibranch bivalve (
Limopsis hirtella), which burrows in deep sediment (30 m) in the Southern Ocean, by Jarms and Tiemann (1996) (as the hydrozoan
Microhydrula limopsicola), while the stauromedusa stage is witnessed mostly on rocks and macroalgae (0–8 m deep) [
7,
16]. In light of these reports, a more thorough survey of the benthos at the Kitsunezaki survey site could lead to the discovery of early life stages of these staurozoans, on yet unexamined substrates, such as other algae species or even bivalve shells.
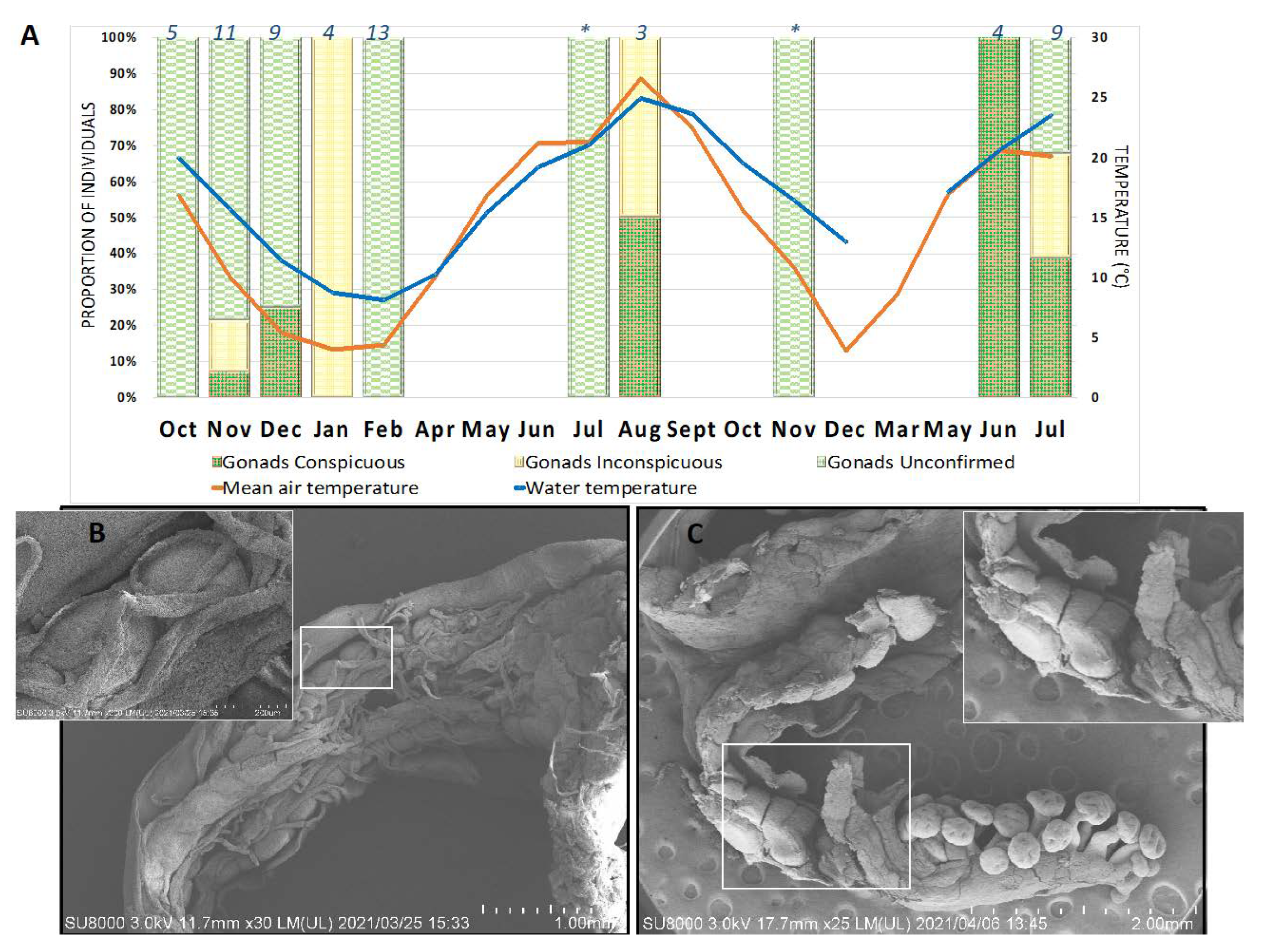
Our stauromedusa distribution data are inconclusive regarding the putative seasonality of reproduction in H. inabai, as individuals with gonads were collected both in cold (October–May) and warm months (June–September). This is in part due to insufficient sampling at the Kitsunezaki collection site during all months of the year, but also due to our inability to differentiate the two lineages of H. inabai, prior to molecular analysis. However, as spawning was not observed in this study, systematic long-term observations are needed to test the hypothesis of whether, as with H. octoradiatus (see Wietrzykowski 1912), one or both lineages of H. inabai are capable of spawning throughout the year.
The absence of Calvadosia nagatensis samples during colder months during this study—September through June—suggests this species has a seasonal appearance during the summer months in the Tohoku region. However, Oka (1897) described C. nagatensis (as Lucernaria nagatensis) based on a specimen collected in April, while Kishinouye (1902) subsequently reported this species in Japan during winter months as well (e.g., in Misaki) and Uchida (1929) in both summer and winter months. Therefore, long-term quantitative studies involving surveys of the algal beds at the Kitsunezaki site are needed to understand the seasonality and early life history of C. nagatensis in the Tohoku region.
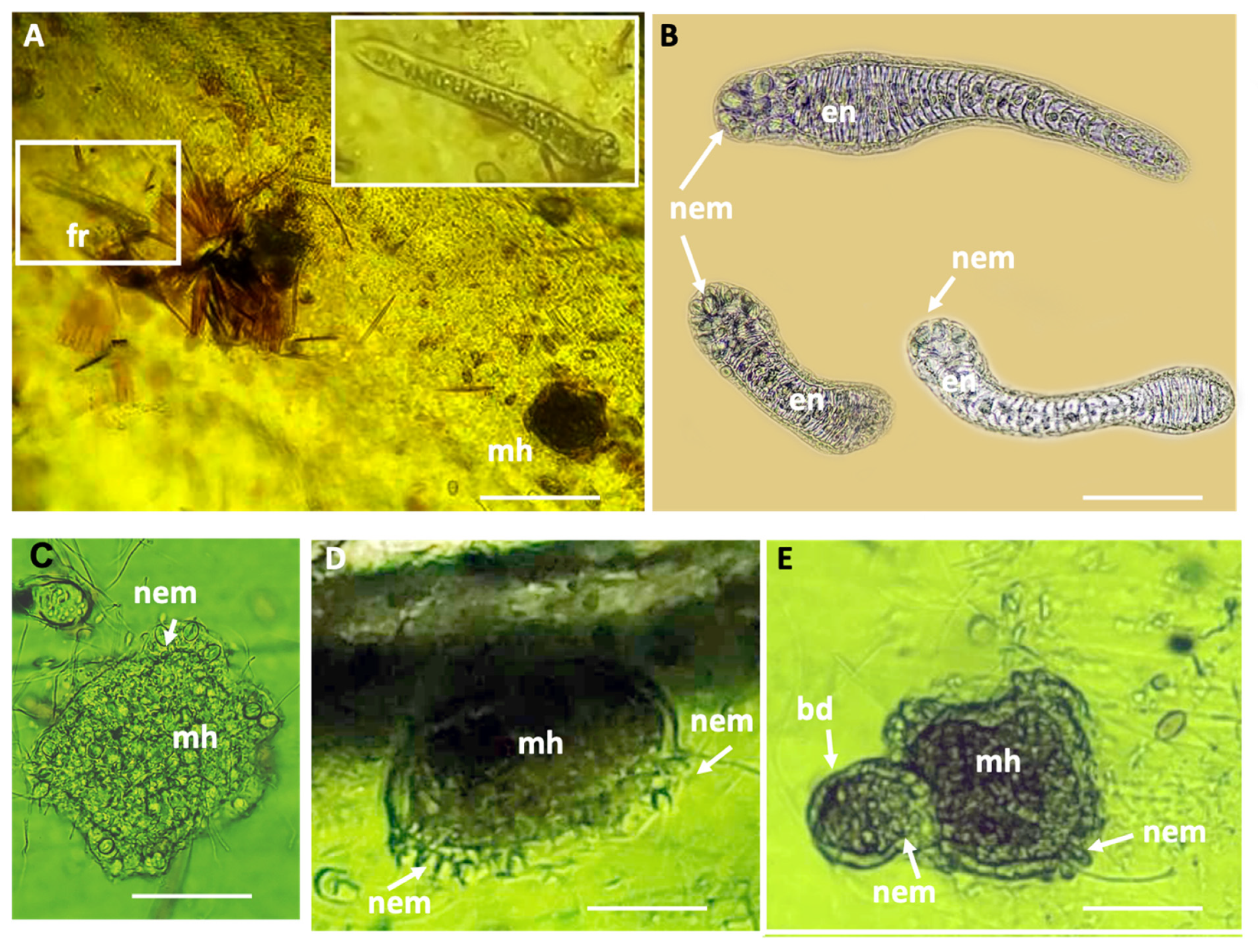
4.4. Haliclystus inabai Life Cycle
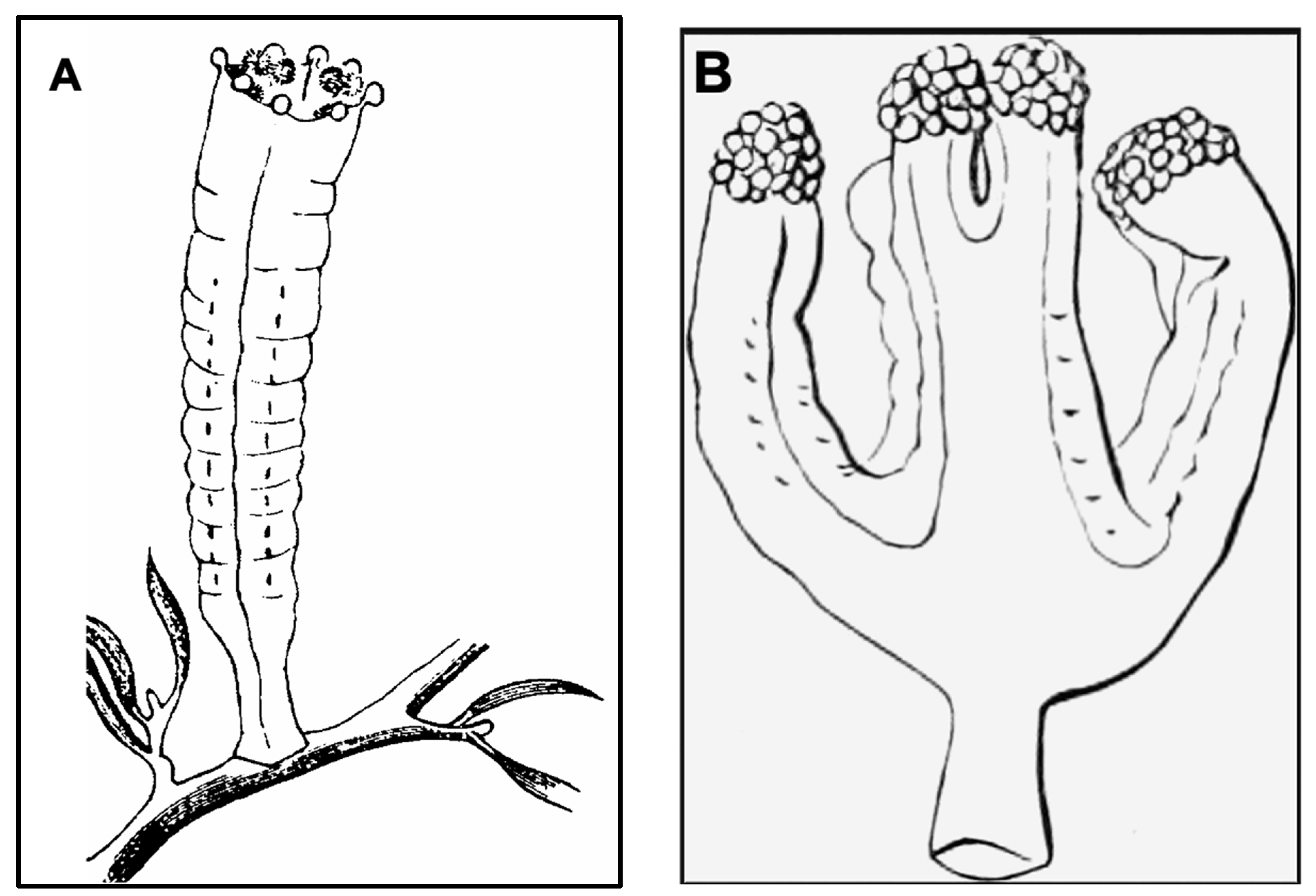
The life cycles of most Staurozoa species remain enigmas, and despite having reared the stauromedusae specimens collected from Kitsunezaki for more than two months in the lab, it was impossible to further investigate their life history. However, thanks to our collaboration with aquariums specializing in rearing jellyfish for exhibition, we were able to gain an initial understanding of the early life history of H. inabai lineage 2, based on photo and video documentation from the Aquarium du Québec and comparisons with earlier works on other Haliclystus species (Miranda et al. 2010; Wietrzykowski 1912).
In life cycle studies on
H. octoradiatus [
15], the hemispherical settled larval stage, whose ectoderm is replete with nematocysts, gives rise laterally to motile frustules (undergoing frustulation), which then settle and go through several development stages before becoming stauropolyps. In this study, we documented the settled larval stages for
H. inabai, which were previously only known in two other species (
H. octoradiatus and
H. antarcticus) for which it was dubbed the “microhydrula” stage [
16]. Similar to those reports, the microhydrula stage in
H. inabai has numerous nematocysts interspersed in the ectoderm. We also witnessed what appeared to be a lateral bud producing a motile frustule, the latter of which we documented in abundance. While frutules are on the substrate, the large size and number of penetrant rhopaloids nematocysts around the distal mouth permit them to capture relatively large prey (such as
Artemia, seen captured in this study). Though only a single putative asexual bud was witnessed in this study, reportedly, asexual buds can be rapidly produced and number up to four in
H. octoradiatus [
15].
In this study, the cnidome of
H. inabai at both frustule and microhydrula stages appear to contain only one type of rhopaloid, in contrast to stauromedusae, which have at least four types of nematocysts—type I and type II isorhizas, euryteles, and birhopaloids. It is not uncommon for cnidomes to vary at different development stages in a single species, and the putative ontogenetic changes in cnidome witnessed here are similar to those for
H. antarcticus [
16]. Further examination of the cnidome of
H. inabai is needed to delineate nematocyst types present at different life stages.
4.5. Ecological Response and Conservation
The rocky coastal environment of Kitsunezaki has historically been dominated by several species of algae including
Sargassum spp,
Gelidium elegans, and
Eisenia bicyclis (Suzuki et al. 2017), the latter two being macroalgae species associated with the two staurozoan species described in the present study. However, on 11 March 2011, a megathrust earthquake (of magnitude 9.0), known as the Great East Japan Earthquake, occurred off the Pacific coast of northeastern Japan, which caused a tsunami with an estimated height of 15 m in the Oshika Peninsula and devastation to the Tohoku region (Geospatial Information Authority of Japan 2011). Since then, studies have examined correlations between the earthquake and resulting subsidence and the decline of macroalgae epibionts such as abalone, sea urchin, and other fishery resources (reviewed in [
41]). In the Kitsunezaki site, where subsidence of 0.9 m occurred, a shift of
Eisenia bicyclis distribution towards the shallower area and a decline in juvenile recruitment of the kelp was observed at the offshore margin of the study site [
19].
While our study is the first report of staurozoans from Kitsunezaki, staurozoan specimens corresponding to
Haliclystus tenuis and
Calvadosia cruciformis from Ayukawa, Miyagi Prefecture (Tohoku Region, Japan), a little south on the Oshika Peninsula from our study site, were previously accessioned into the Saito Ho-on Kai Museum of Natural History [
42]. In addition to the above two species, a single specimen of
Calvadosia tsingtaoensis was reported (as
Sasakiella tsingtaoensis) from Onagawa Bay, north of Kitsunezaki on the opposite side of the peninsula [
43], but it lacked both interradial and perradial primary tentacles, suggesting it may instead corresponded to
C. nagatensis. No further report exists for
C. tsingtaoensis in Japan. The absence of biodiversity census data for the Kitsunezaki study site just prior to the Great East Japan Earthquake and Tsunami in 2011 precludes pinpointing of the origin or introduction timeframe of
C. nagatensis and
H. inabai (either lineage) in this geographic locality. Some studies have reported a decline or complete loss of staurozoan biodiversity in areas with strong anthropogenic effects (e.g., pollution), while others suggest that substrate damage, even from natural disasters, could also result in the disappearance of staurozoans (reviewed in Miranda et al. 2018).
Breakwater restoration work commenced at Kitsunezaki in July 2014, and a subsequent study suggested that the breakwater restoration work had a negative effect on the density and age structure of
Eisenia bicyclis population in the Kisunezaki survey site [
44]. The ratio of young plants decreased while the whole population became dominated by older plants; inevitable senescence of the adult population could lead to a decline in the
E. bicyclis population in the future [
44], and consequently reduce staurozoan recruits on
E. bicyclis. Studies are needed to understand linkage between the population dynamics of the macroalgae and stauromedusa presence, reproduction and early life cycle, or even seasonality, as this study did not find any obvious pattern with respect to gonad state.
5. Conclusions
We present the first genetic barcode data for C. nagatensis in Japan, and compare these data with that of global species of Calvadosia for which sequence data are generally lacking. Likewise, prior to this study, publicly available molecular sequence data existed only for one H. inabai lineage 1 individual from Japan (Hokkaido), as well as one individual of H. inabai lineage 2 from Australia (Victoria). Analysis of our additional substantial sequence data reveals that H. inabai comprises two distinct species—one including only samples from Japanese localities (H. inabai lineage 1), and the other including samples from locations in several continents (H. inabai lineage 2), either from nearshore waters or found in aquariums. Increased sampling efforts and further comparative analyses are needed before a formal new species description can be made for the unnamed lineage. A consolidation of 100 years of literature documenting substrate variation for these two species revealed a clear propensity by the two named species of stauromedusae for settlement on macroalgae and seagrass. While C. nagatensis is limited to several species of seaweed (algae or kelp) and seagrass, H. inabai (lineage not considered) inhabits a much more diverse suite of substrates.
In addition to analyzing
H. inabai specimens collected from the wild from both lineages 1 and 2, we also documented individuals of
H. inabai lineage 2 thriving in captivity, in two major aquariums in Japan (Kamo Aquarium in Tsuruoka and Kagoshima Aquarium in Kagoshima) and in Canada (Aquarium du Québec). Photo and video documentation of early life stages in the latter permitted confirmation that the life cycle of
H. inabai lineage 2 includes “microhydrula” and frustule stages, similar to those described in two other species of
Haliclystus:
H. octoradiatus and
H. antarcticus [
15,
16]. In light of these new findings, we discuss whether the broad, global distribution of
H. inabai lineage 2 is linked to its multiphasic life cycle, which includes the microscopic sessile microhydrula stage that buds motile frustules during its life cycle, and surmise that its non-specific substrate preference facilitates global distribution of
H. inabai lineage 2, potentially through the aquarium trade. This study underlines the importance of collaboration between academia and aquariums. Scientists in both settings can produce research findings of reciprocal relevance, and collaborations can take advantage of complementary expertise, e.g., systematics and live jellyfish husbandry, to make more rapid progress in advancing our understanding of marine biodiversity.
The discovery of these staurozoan species in the Tohoku Region of Japan—an area that was devastated by the 2011 Great East Japan Earthquake—underlines the importance of documenting marine species in both healthy and recovering ecosystems. The crucial relationship between the condition of the macroalgae and its epibionts and their respective responses to environmental disaster—both natural and anthropogenic—are being increasingly highlighted. Although this study focused on species of stalked jellyfish, many other species—including important commercial fisheries species—require macroalgae substrates to complete their life cycles. The discoveries herein were made possible by scientific collaboration with aquarists, and would be very difficult to observe in nature. This work is proof that multi-disciplinary research facilitates a synergistic approach to addressing issues related to ocean ecosystems, biodiversity, and conservation.



 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}