
The Dual Cardiovascular Effect of Centrally Administered Clonidine: A Comparative Study between Pentobarbital- and Ketamine/Xylazine-Anesthetized Rats

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
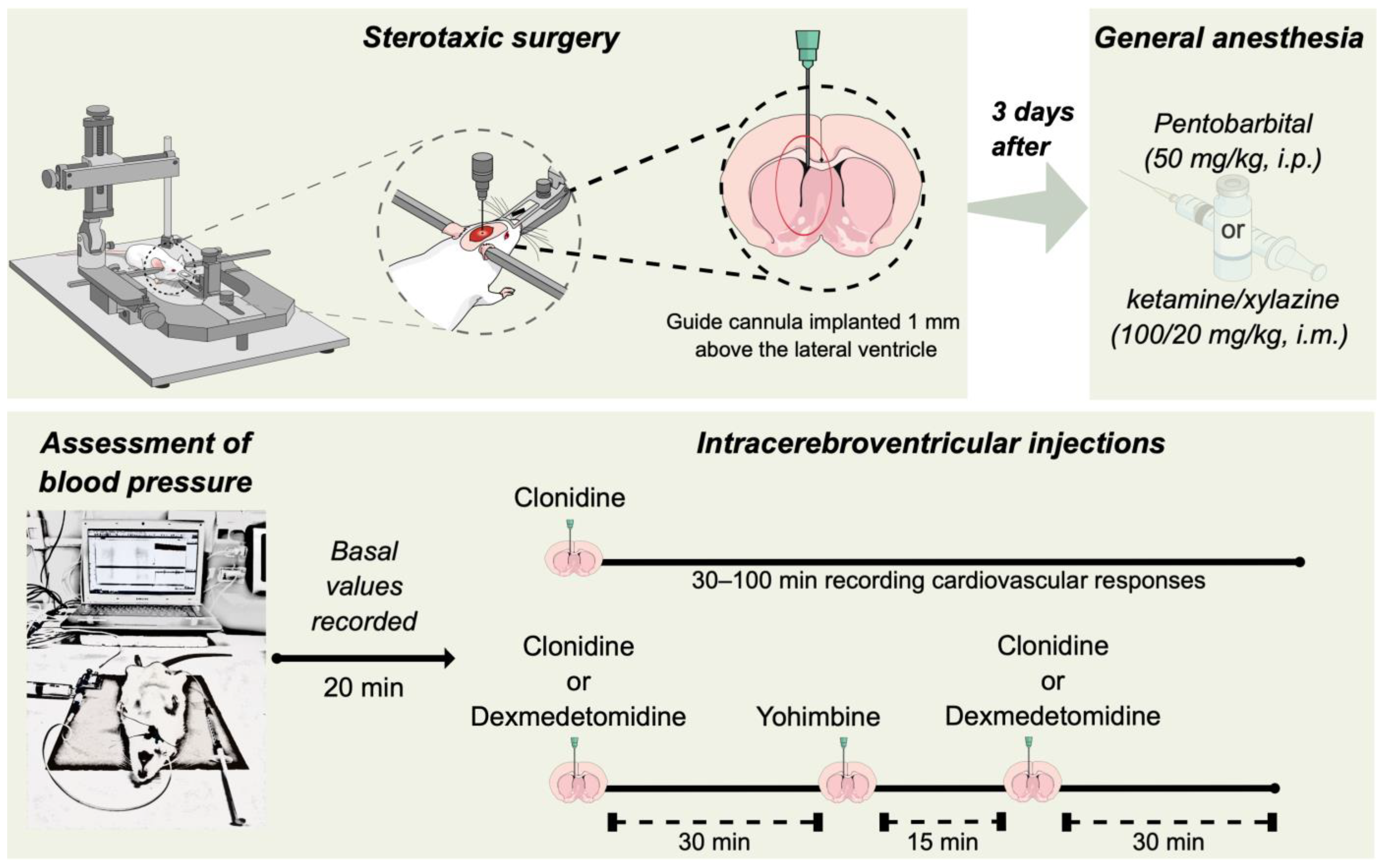
2.2. Stereotaxic Surgery
2.3. Intracerebroventricular (i.c.v.) Drug Administration
2.4. Assessment of Systemic Blood Pressure
2.5. Comparison of the Cardiovascular Effects of Centrally Administered Clonidine in Ketamine/Xylazine and Pentobarbital-Anesthetized Rats
2.6. Evaluation of the Role of α2-Adrenoceptors in the Pressor Effect of Clonidine
2.7. Drugs and Solutions
2.8. Data Presentation and Statistical Analyses
3. Results
3.1. The Cardiovascular Effects of Intracerebroventricularly Injected Clonidine Depending on the Anesthetic Choice
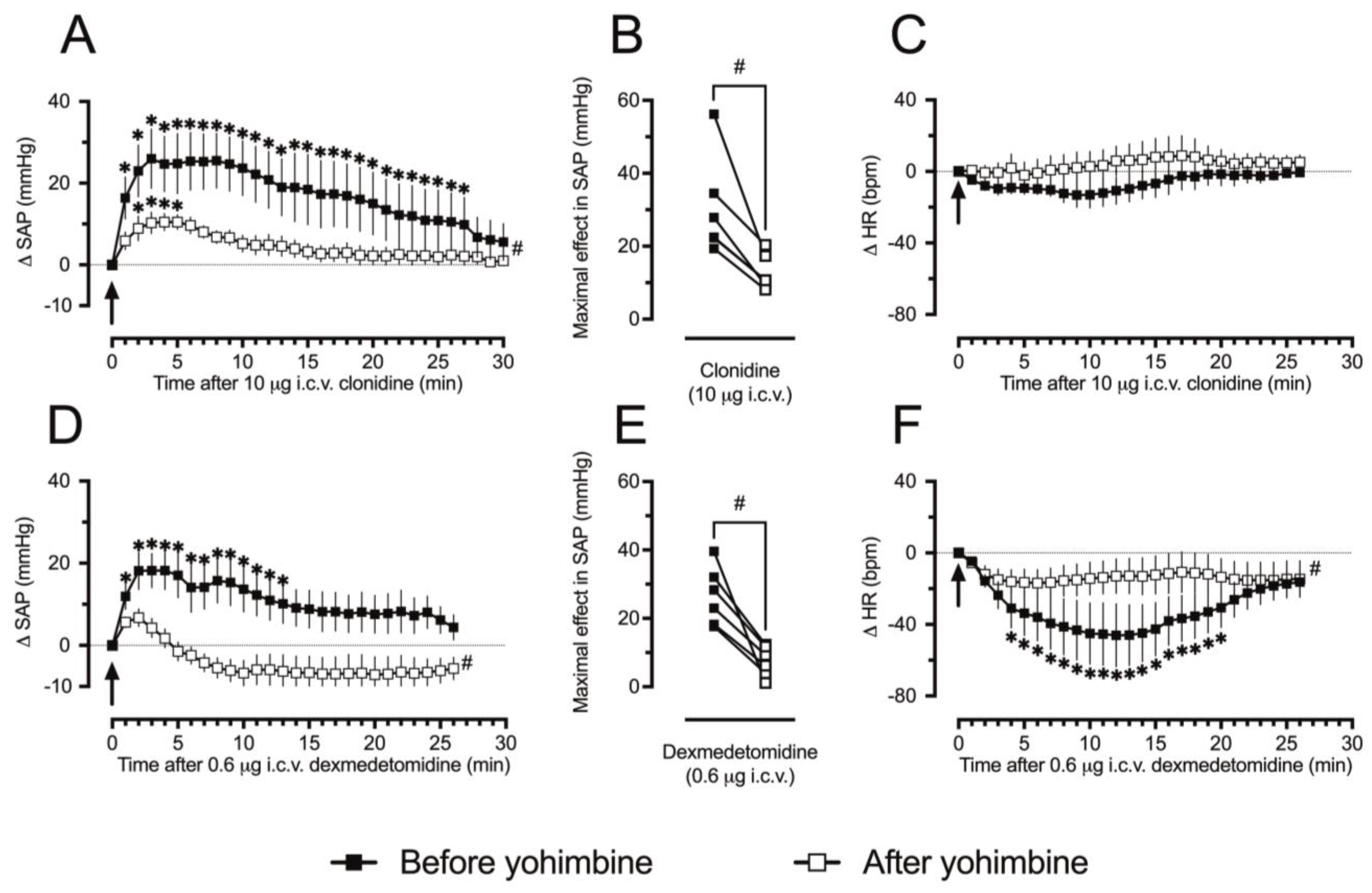
3.2. The Pressor Responses to Clonidine in Ketamine/Xylazine-Anesthetized Rats Involve α2-ARs
4. Discussion
5. Limitations of This Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bousquet, P.; Hudson, A.; Garcia-Sevilla, J.A.; Li, J.X. Imidazoline Receptor System: The Past, the Present, and the Future. Pharmacol. Rev. 2020, 72, 50–79. [Google Scholar] [CrossRef] [PubMed]
- Minneman, K.P.; Theroux, T.L.; Hollinger, S.; Han, C.; Esbenshade, T.A. Selectivity of agonists for cloned alpha 1-adrenergic receptor subtypes. Mol. Pharmacol. 1994, 46, 929–936. [Google Scholar] [PubMed]
- Maruta, T.; Nemoto, T.; Satoh, S.; Kanai, T.; Yanagita, T.; Wada, A.; Tsuneyoshi, I. Dexmedetomidine and clonidine inhibit the function of NaV1.7 independent of α2-adrenoceptor in adrenal chromaffin cells. J. Anesth. 2011, 25, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Knaus, A.; Zong, X.; Beetz, N.; Jahns, R.; Lohse, M.J.; Biel, M.; Hein, L. Direct inhibition of cardiac hyperpolarization-activated cyclic nucleotide-gated pacemaker channels by clonidine. Circulation 2007, 115, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Musgrave, I.F.; Krautwurst, D.; Hescheler, J.; Schultz, G. Clonidine and cirazoline inhibit activation of nicotinic channels in PC-12 cells. Ann. N. Y. Acad. Sci. 1995, 763, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Schaner, M.E.; Giacomini, K.M. Functional characterization of an organic cation transporter (hOCT1) in a transiently transfected human cell line (HeLa). J. Pharmacol. Exp. Ther. 1998, 286, 354–361. [Google Scholar] [PubMed]
- Egleme, C.; Godfraind, T.; Miller, R.C. Enhanced responsiveness of rat isolated aorta to clonidine after removal of the endothelial cells. Br. J. Pharmacol. 1984, 81, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Ren, L.M.; Lu, H.G.; Zhang, X. Potentiation by yohimbine of alpha-adrenoceptor-mediated vasoconstriction in response to clonidine in the rabbit ear vein. Eur. J. Pharmacol. 2008, 589, 201–205. [Google Scholar] [CrossRef]
- McRaven, D.R.; Kroetz, F.W.; Kioschos, J.M.; Kirkendall, W.M. The effect of clonidine on hemodynamics in hypertensive patients. Am. Heart J. 1971, 81, 482–489. [Google Scholar] [CrossRef]
- Kobinger, W.; Walland, A. Investigations into the mechanism of the hypotensive effect of 2-(2,6-dichlorphenylamino)-2-imidazoline-HCl. Eur. J. Pharmacol. 1967, 2, 155–162. [Google Scholar] [CrossRef]
- Aicher, S.A.; Drake, C.T. Clonidine evokes vasodepressor responses via alpha2-adrenergic receptors in gigantocellular reticular formation. J. Pharmacol. Exp. Ther. 1999, 289, 688–694. [Google Scholar] [PubMed]
- Pettinger, W.A. Drug therapy: Clonidine, a new antihypertensive drug. N. Engl. J. Med. 1975, 293, 1179–1180. [Google Scholar] [CrossRef] [PubMed]
- Unger, T.; Borghi, C.; Charchar, F.; Khan, N.A.; Poulter, N.R.; Prabhakaran, D.; Ramirez, A.; Schlaich, M.; Stergiou, G.S.; Tomaszewski, M.; et al. 2020 International Society of Hypertension Global Hypertension Practice Guidelines. Hypertension 2020, 75, 1334–1357. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Segal, S.; Kollins, S.H.; Khayrallah, M. Clonidine extended-release tablets for pediatric patients with attention-deficit/hyperactivity disorder. J. Am. Acad. Child Adolesc. Psychiatry 2011, 50, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, L.L.; Follett, K.A.; Rosenquist, R.W. Long-term outcomes during treatment of chronic pain with intrathecal clonidine or clonidine/opioid combinations. J. Pain Symptom Manag. 2003, 26, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Naguy, A. Clonidine Use in Psychiatry: Panacea or Panache. Pharmacology 2016, 98, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Luft, F.C.; Veelken, R.; Becker, H.; Ganten, D.; Lang, R.E.; Unger, T. Effect of urapidil, clonidine, and prazosin on sympathetic tone in conscious rats. Hypertension 1986, 8, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Dabire, H.; Richer, C. Implication of the central nervous system in the systemic and regional hemodynamics of two centrally acting hypotensive drugs, flesinoxan and clonidine, in the rat. J. Cardiovasc. Pharmacol. 1991, 18, 605–613. [Google Scholar] [CrossRef]
- Borkowski, K.R.; Finch, L. A comparison of the cardiovascular effects of centrally administered clonidine and adrenaline in the anaesthetized rat. J. Pharm. Pharmacol. 1979, 31, 16–19. [Google Scholar] [CrossRef]
- Dobrucki, L.W.; Cabrera, C.L.; Bohr, D.F.; Malinski, T. Central hypotensive action of clonidine requires nitric oxide. Circulation 2001, 104, 1884–1886. [Google Scholar] [CrossRef]
- Imai, Y.; Abe, K.; Sasaki, S.; Minami, N.; Nihei, M.; Munakata, M.; Sekino, H.; Yoshinaga, K. Modulation of cardiovascular depressant action of clonidine by pentobarbital anesthesia in rats. Tohoku J. Exp. Med. 1989, 157, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.G.; Zeng, J.; Yuan, W.J.; Su, D.F.; Wang, W.Z. Comparative study of NMDA and AMPA/kainate receptors involved in cardiovascular inhibition produced by imidazoline-like drugs in anaesthetized rats. Exp. Physiol. 2007, 92, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Colombari, E.; Camargo, L.A.; Saad, W.A.; Renzi, A.; De Luca Junior, L.A.; Menani, J.V. Cardiovascular effects of central clonidine in conscious rats after hypothalamic lesions. J. Auton. Nerv. Syst. 1992, 40, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, H.; Takasaki, K. Central alpha-2 adrenoceptor-mediated hypertensive response to clonidine in conscious, normotensive rats. J. Pharmacol. Exp. Ther. 1986, 236, 810–818. [Google Scholar] [PubMed]
- Nakamura, S.; Kawasaki, H.; Takasaki, K. Intracerebroventricular treatment with vasopressin V1-receptor antagonist inhibits centrally-mediated pressor response to clonidine in conscious rats. Jpn. J. Pharmacol. 1993, 63, 447–453. [Google Scholar] [CrossRef]
- Muroi, Y.; Theusch, C.M.; Czajkowski, C.; Jackson, M.B. Distinct structural changes in the GABAA receptor elicited by pentobarbital and GABA. Biophys. J. 2009, 96, 499–509. [Google Scholar] [CrossRef]
- Irnaten, M.; Walwyn, W.M.; Wang, J.; Venkatesan, P.; Evans, C.; Chang, K.S.; Andresen, M.C.; Hales, T.G.; Mendelowitz, D. Pentobarbital enhances GABAergic neurotransmission to cardiac parasympathetic neurons, which is prevented by expression of GABAAε subunit. Anesthesiology 2002, 97, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Idvall, J.; Ahlgren, I.; Aronsen, K.R.; Stenberg, P. Ketamine infusions: Pharmacokinetics and clinical effects. Br. J. Anaesth. 1979, 51, 1167–1173. [Google Scholar] [CrossRef]
- Dampney, R.A. Central neural control of the cardiovascular system: Current perspectives. Adv. Physiol. Educ. 2016, 40, 283–296. [Google Scholar] [CrossRef]
- Schreihofer, A.M.; Guyenet, P.G. Identification of C1 presympathetic neurons in rat rostral ventrolateral medulla by juxtacellular labeling in vivo. J. Comp. Neurol. 1997, 387, 524–536. [Google Scholar] [CrossRef]
- McAuley, M.A.; Macrae, I.M.; Reid, J.L. The cardiovascular actions of clonidine and neuropeptide-Y in the ventrolateral medulla of the rat. Br. J. Pharmacol. 1989, 97, 1067–1074. [Google Scholar] [CrossRef]
- Mermet, C.; Quintin, L. Effect of clonidine on catechol metabolism in the rostral ventrolateral medulla: An in vivo electrochemical study. Eur. J. Pharmacol. 1991, 204, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Z.; Wang, L.G.; Gao, L.; Wang, W. Contribution of AMPA/kainate receptors in the rostral ventrolateral medulla to the hypotensive and sympathoinhibitory effects of clonidine. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1232–R1238. [Google Scholar] [CrossRef] [PubMed]
- Yamazato, M.; Sakima, A.; Nakazato, J.; Sesoko, S.; Muratani, H.; Fukiyama, K. Hypotensive and sedative effects of clonidine injected into the rostral ventrolateral medulla of conscious rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R1868–R1876. [Google Scholar] [CrossRef] [PubMed]
- Ernsberger, P.; Giuliano, R.; Willette, R.N.; Reis, D.J. Role of imidazole receptors in the vasodepressor response to clonidine analogs in the rostral ventrolateral medulla. J. Pharmacol. Exp. Ther. 1990, 253, 408–418. [Google Scholar] [PubMed]
- Sun, M.K. Pharmacology of reticulospinal vasomotor neurons in cardiovascular regulation. Pharmacol. Rev. 1996, 48, 465–494. [Google Scholar]
- Trolin, G. Effects of pentobarbitone and decerebration on the clonidine-induced circulatory changes. Eur. J. Pharmacol. 1975, 34, 1–7. [Google Scholar] [CrossRef]
- Fehr, S.B.; Zalunardo, M.P.; Seifert, B.; Rentsch, K.M.; Rohling, R.G.; Pasch, T.; Spahn, D.R. Clonidine decreases propofol requirements during anaesthesia: Effect on bispectral index. Br. J. Anaesth. 2001, 86, 627–632. [Google Scholar] [CrossRef]
- Rajabi, M.; Razavizade, M.R.; Hamidi-Shad, M.; Tabasi, Z.; Akbari, H.; Hajian, A. Magnesium Sulfate and Clonidine; Effects on Hemodynamic Factors and Depth of General Anesthesia in Cesarean Section. Anesth. Pain Med. 2020, 10, e100563. [Google Scholar] [CrossRef]
- Sleigh, J.; Harvey, M.; Voss, L.; Denny, B. Ketamine—More mechanisms of action than just NMDA blockade. Trends Anaesth. Crit. Care 2014, 4, 76–81. [Google Scholar] [CrossRef]
- Marland, S.; Ellerton, J.; Andolfatto, G.; Strapazzon, G.; Thomassen, O.; Brandner, B.; Weatherall, A.; Paal, P. Ketamine: Use in anesthesia. CNS Neurosci. Ther. 2013, 19, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, M.D. A review of the physiological effects of alpha2-agonists related to the clinical use of medetomidine in small animal practice. Can. Vet. J. 2003, 44, 885–897. [Google Scholar] [PubMed]
- Van Pelt, L.F. Ketamine and xylazine for surgical anesthesia in rats. J. Am. Vet. Med. Assoc. 1977, 171, 842–844. [Google Scholar] [PubMed]
- Ebihara, H.; Kawasaki, H.; Nakamura, S.; Takasaki, K.; Wada, A. Pressor response to microinjection of clonidine into the hypothalamic paraventricular nucleus in conscious rats. Brain Res. 1993, 624, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Hanafusa, N.; Okamoto, K.; Takatori, S.; Kawasaki, H. Involvement of hypothalamic periventricular GABAergic nerves in the central pressor response to clonidine in freely-moving conscious rats. J. Pharmacol. Sci. 2012, 118, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Struyker Boudier, H.A.; Smeets, G.W.; Brouwer, G.M.; van Rossum, J.M. Hypothalamic alpha adrenergic receptors in cardiovascular regulation. Neuropharmacology 1974, 13, 837–846. [Google Scholar] [CrossRef]
- Philbin, K.E.; Bateman, R.J.; Mendelowitz, D. Clonidine, an alpha2-receptor agonist, diminishes GABAergic neurotransmission to cardiac vagal neurons in the nucleus ambiguus. Brain Res. 2010, 1347, 65–70. [Google Scholar] [CrossRef]
- Sharp, D.B.; Wang, X.; Mendelowitz, D. Dexmedetomidine decreases inhibitory but not excitatory neurotransmission to cardiac vagal neurons in the nucleus ambiguus. Brain Res. 2014, 1574, 1–5. [Google Scholar] [CrossRef]
- Virtanen, R.; Savola, J.M.; Saano, V.; Nyman, L. Characterization of the selectivity, specificity and potency of medetomidine as an alpha 2-adrenoceptor agonist. Eur. J. Pharmacol. 1988, 150, 9–14. [Google Scholar] [CrossRef]
- Ernsberger, P.; Meeley, M.P.; Mann, J.J.; Reis, D.J. Clonidine binds to imidazole binding sites as well as alpha 2-adrenoceptors in the ventrolateral medulla. Eur. J. Pharmacol. 1987, 134, 1–13. [Google Scholar] [CrossRef]
- Khan, Z.P.; Ferguson, C.N.; Jones, R.M. Alpha-2 and imidazoline receptor agonists. Their pharmacology and therapeutic role. Anaesthesia 1999, 54, 146–165. [Google Scholar] [CrossRef] [PubMed]
- Freedman, R.R.; Moten, M. Gender differences in modulation of peripheral vascular adrenoceptors. Ann. Behav. Med. 1995, 17, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, J.; Rosano, G.; Walther, T.; Duarte, J.; Niessner, A.; Kaski, J.C.; Ceconi, C.; Drexel, H.; Kjeldsen, K.; Savarese, G.; et al. Gender differences in the effects of cardiovascular drugs. Eur. Heart J. Cardiovasc. Pharmacother. 2017, 3, 163–182. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsubara, N.K.; da Silva-Santos, J.E. The Dual Cardiovascular Effect of Centrally Administered Clonidine: A Comparative Study between Pentobarbital- and Ketamine/Xylazine-Anesthetized Rats. Future Pharmacol. 2024, 4, 17-29. https://doi.org/10.3390/futurepharmacol4010003
Matsubara NK, da Silva-Santos JE. The Dual Cardiovascular Effect of Centrally Administered Clonidine: A Comparative Study between Pentobarbital- and Ketamine/Xylazine-Anesthetized Rats. Future Pharmacology. 2024; 4(1):17-29. https://doi.org/10.3390/futurepharmacol4010003
Chicago/Turabian StyleMatsubara, Natália Kimie, and José Eduardo da Silva-Santos. 2024. "The Dual Cardiovascular Effect of Centrally Administered Clonidine: A Comparative Study between Pentobarbital- and Ketamine/Xylazine-Anesthetized Rats" Future Pharmacology 4, no. 1: 17-29. https://doi.org/10.3390/futurepharmacol4010003