
Use of Plant Extracts, Bee-Derived Products, and Probiotic-Related Applications to Fight Multidrug-Resistant Pathogens in the Post-Antibiotic Era

 , , and
, , and 
Abstract
:1. Introduction
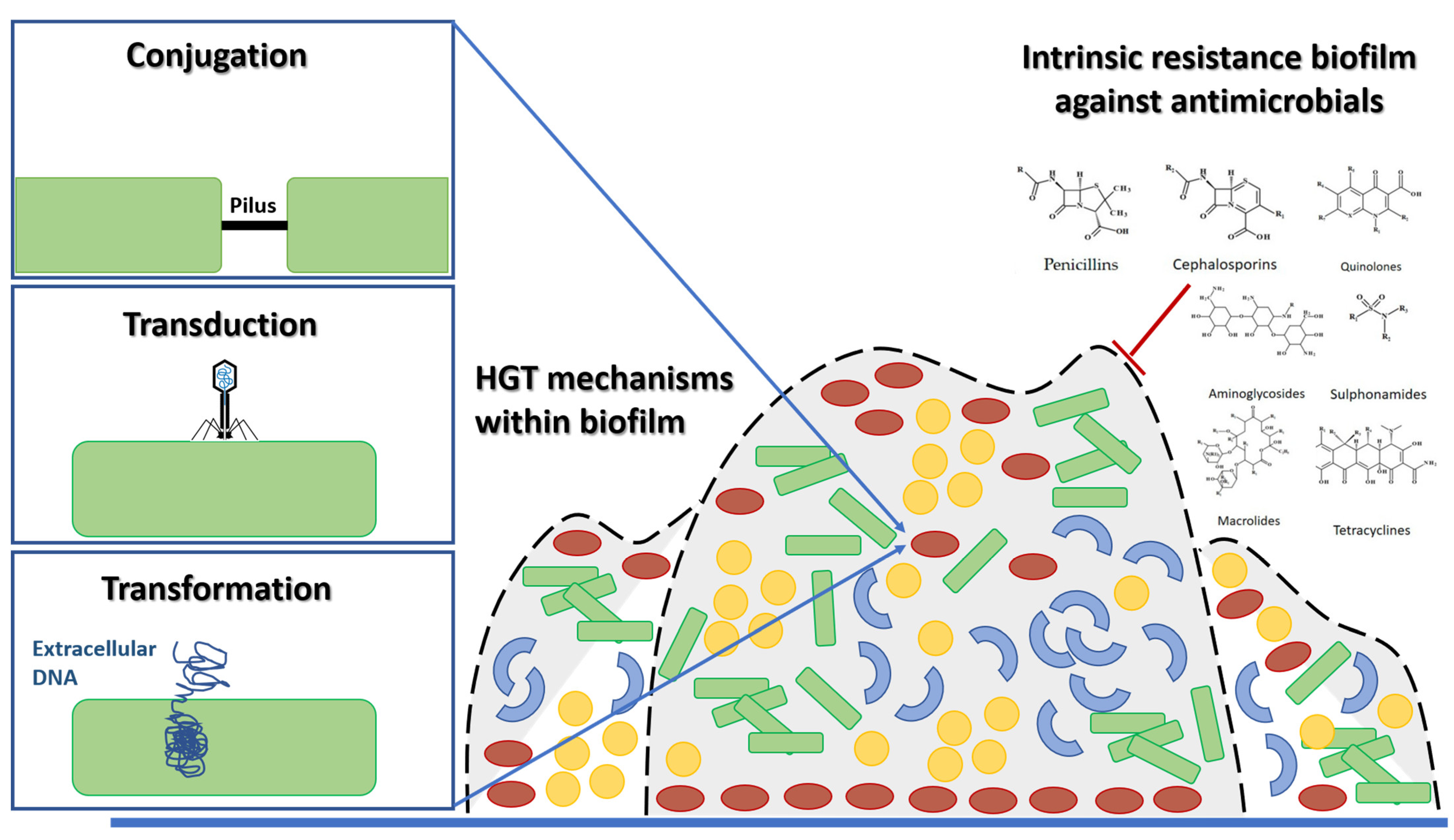
2. Rise of MDR Pathogens and Future Trends concerning the Global Infectious Disease Crisis
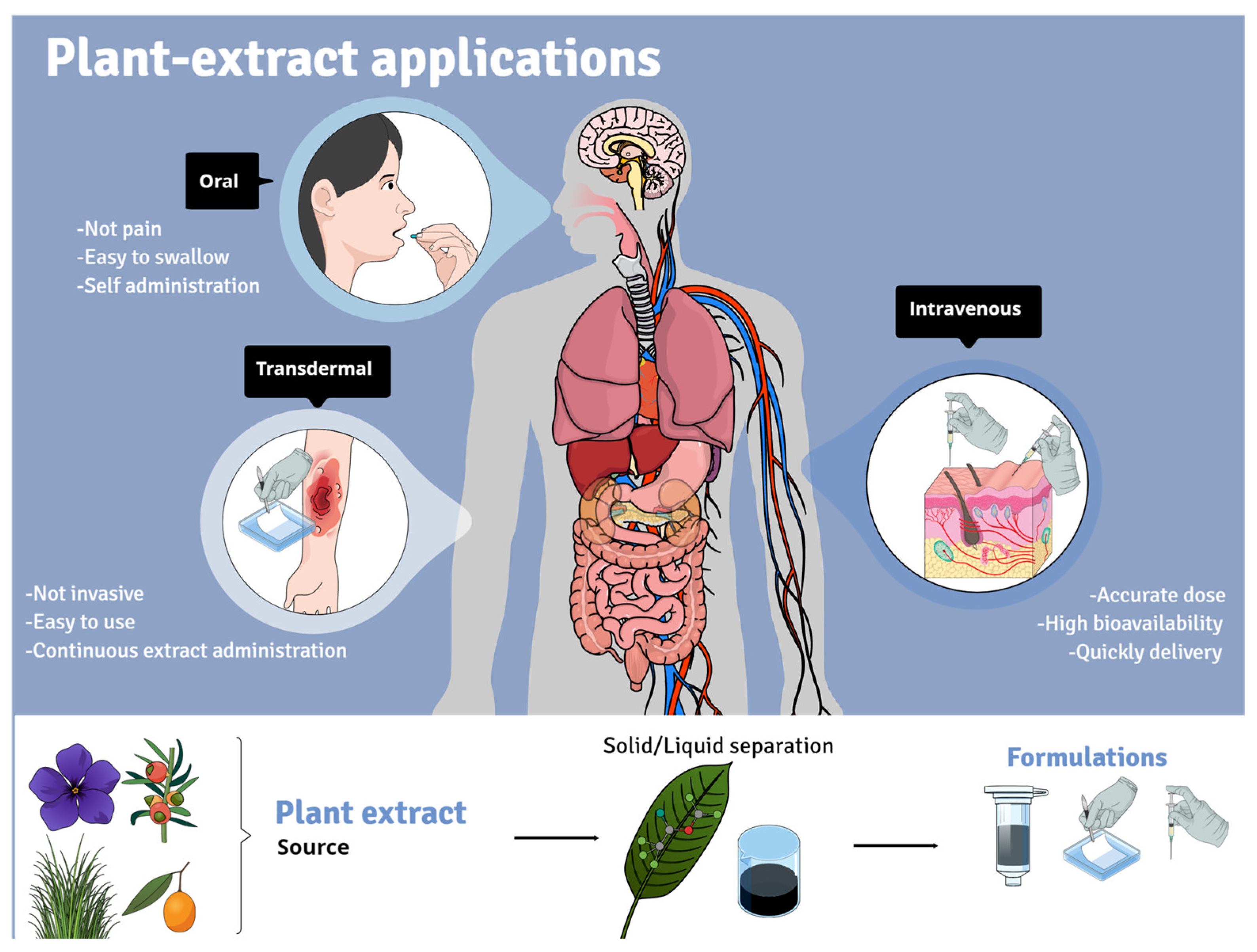
3. Plant Extracts
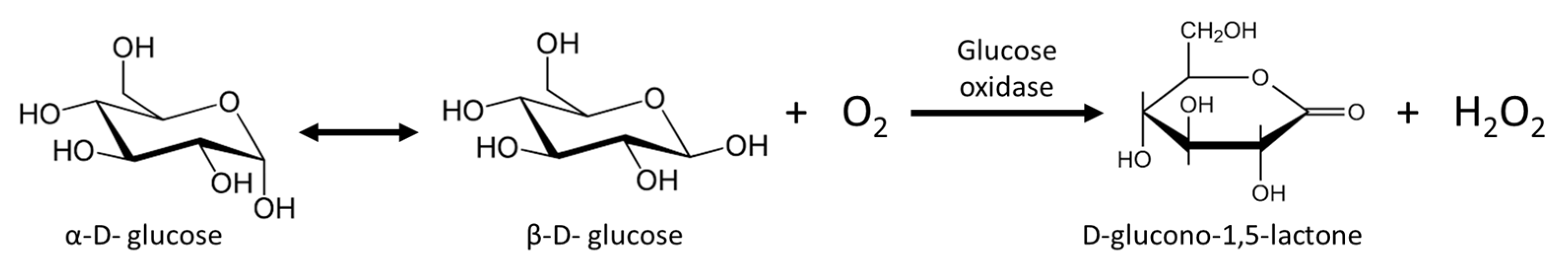
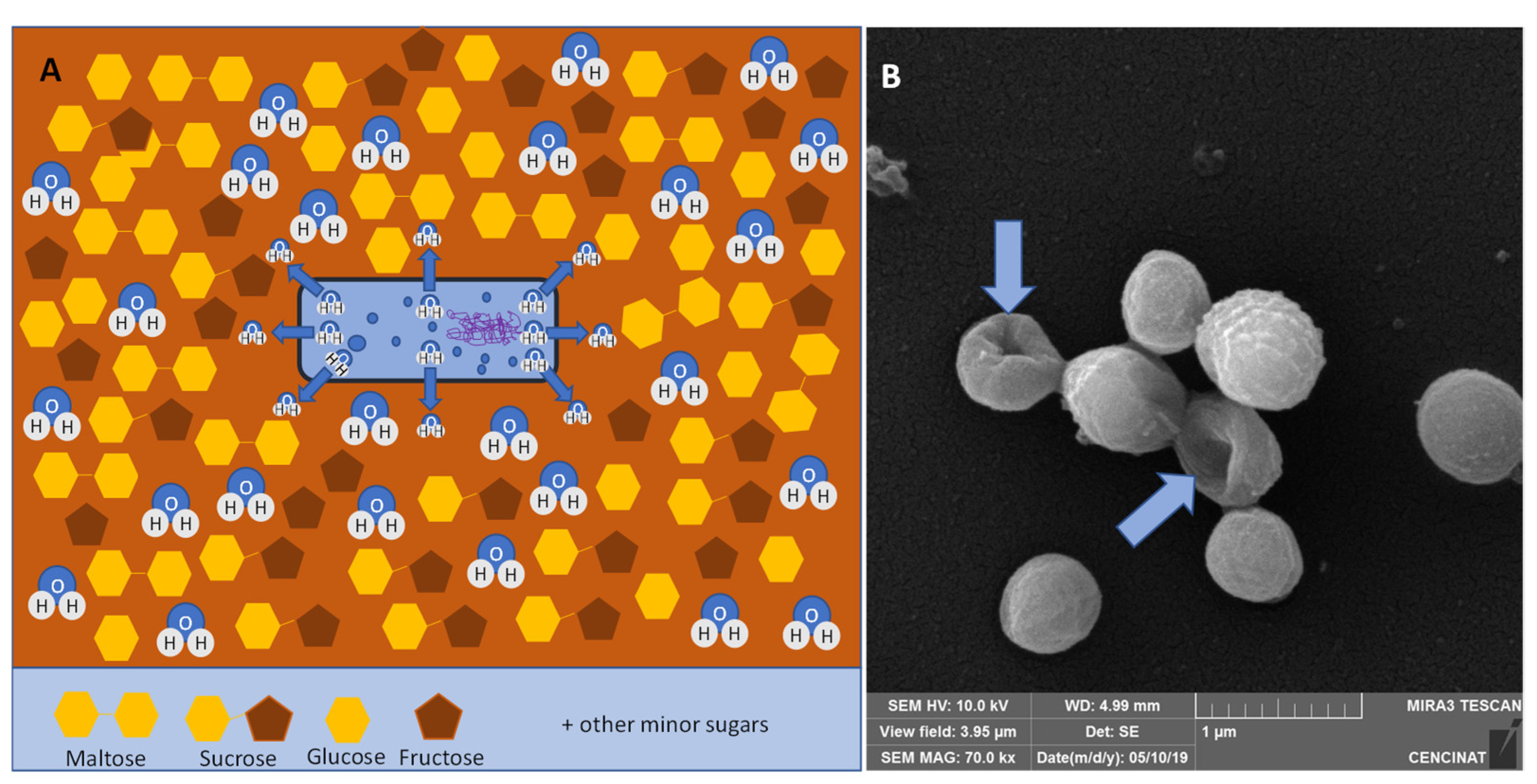
4. Honey and Propolis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographical Origin | Bacterial Strain (Gram-Positive) | References | Bacterial Strain (Gram-Negative) | References |
|---|---|---|---|---|
| Australia | S. aureus ATCC 25923 | [109] | K. pneumoniae ATCC 13883 | [109] |
| Brazil | B. subtilis ATCC 6633, Enterococcus spp., E. faecalis ATTC 29212, ATCC 43300 and ESA 553, Micrococcus luteus ATCC 10240, S. aureus ATCC 6538, ATCC 43300, ATCC 25923, SA 10 and ESA 654, S. epidermidis ATCC 12228 and ESA 675, S. mutans, and S. pyogenes | [110,111,112,113,114,115] | E. coli ATCC 8739, ATCC 25922 and EC06, K. pneumoniae ATCC 4352 and ESA 154, P. mirabilis ATCC 43300 and ESA 37, P. aeruginosa ATCC 25853, ATCC 15442, PA 24 and ESA 22, and Salmonella spp. | [111,113,114,115] |
| Bulgaria | S. aureus ATCC 209 | [116] | E. coli WF | [116] |
| Chile | S. aureus ATCC 25923 and S. pyogenes ISP 364-00 | [117] | E. coli ATCC 25922 and P. aeruginosa ATCC 27853 | [117] |
| Czech Republic | S. aureus ATCC 29213, ATCC 25923 and ATCC 977, S. epidermidis ATCC 14990, S. aureus MRSA/NCTC, S. saprophyticus ATCC 15305, S. oralis ATCC 35037, B. subtilis ATCC 6051, Enterococcus spp., S. agalactiae ATTC 27956, S. pneumoniae ATCC 49619, and S. pyogenes ATCC 12344 | [118] | A. baumani, Burkholderia cepacia, E. cloacae ATCC 700323, E. coli O157:H7, H. influenzae ATCC 49747, K. pneumoniae ATCC 700603., P. aeruginosa ATCC 27853, Salmonella spp., Shigella flexneri, and Y. enterocolitica ATCC 9610 | [118] |
| Germany | S. aureus ATCC 29213, ATCC 25923 and ATCC 977, S. epidermidis ATCC 14990, S. aureus MRSA/NCTC, S. saprophyticus ATCC 15305, S. oralis ATCC 35037, B. subtilis ATCC 6051, Enterococcus spp., S. agalactiae ATTC 27956, S. pneumoniae ATCC 49619, and S. pyogenes ATCC 12344 | [118] | A. baumani, B. cepacia, E. cloacae ATCC 700323, E. coli O157:H7, H. influenzae ATCC 49747, K. pneumoniae ATCC 700603., P. aeruginosa ATCC 27853, Salmonella spp., S. flexneri, and Y. enterocolitica ATCC 9610 | [118] |
| Greece | S. aureus ATCC 25923 and S. epidermidis ATCC 12228 | [119] | E. cloacae ATCC 13047, E. Coli ATCC 25922, P. aeruginosa ATCC 227853, and K. pneumoniae ATCC 13883 | [119] |
| India | S. aureus ATCC 6538P | [120] | – | – |
| Ireland | S. aureus ATCC 29213, ATCC 25923 and ATCC 977, S. epidermidis ATCC 14990, S. aureus MRSA/NCTC, S. saprophyticus ATCC 15305, S. oralis ATCC 35037, B. subtilis ATCC 6051, Enterococcus spp., S. agalactiae ATTC 27956, S. oralis, S. pneumoniae ATCC 49619, and S. pyogenes ATCC 12344 | [118] | A. baumani, B. cepacia, E. cloacae ATCC 700323, E. coli O157:H7, H. influenzae ATCC 49747, K. pneumoniae ATCC 700603, P. aerugino-sa ATCC 27853, Salmonella spp., S. flexneri, and Y. enterocolitica ATCC 9610 | [118] |
| Italy | – | – | Campylobacter jejuni (clinical isolate) and P. aeruginosa P1242 | [121,122] |
| Korea | S. mutans ATCC 25175, S. sobrinus ATCC33478, S. mutans KCOM 1088, KCOM 1091, KCOM 1092, KCOM 1095, KCOM 1097, KCOM 1111, KCOM 1112, KCOM 1113, KCOM 1116, KCOM 1117, KCOM 1118, KCOM 1123, KCOM 1124, KCOM 1126, KCOM 1127, KCOM 1128, KCOM 2762, KCOM 1136, KCOM 1137, KCOM 1139, KCOM 1142, KCOM 1143, KCOM 1145, KCOM 1146, KCOM 1197, KCOM 1200, KCOM 1201, KCOM 1202, KCOM 1203, KCOM 1207, KCOM 1208, KCOM 1209, KCOM 1212, KCOM 1214, KCOM 1217, KCOM 1219, KCOM 1226 (clinical isolates), and S. sobrinus KCOM 1061, KCOM 1150, KCOM 1151, KCOM 1152, KCOM 1153, KCOM 1157, KCOM 1158, KCOM 1159, KCOM 1185, KCOM 1191, KCOM 1193, KCOM 1196, KCOM 1221, KCOM 1228, and KCOM 1218 (clinical isolates) | [123] | – | – |
| Morocco | S. aureus ATCC 6538 and MRSA 2, 15, and 16 (clinical isolates) | [124] | – | – |
| Oman | S. aureus ATCC 209 | [116] | E. coli WF | [116] |
| Poland | S. aureus ATCC 25923 and S. aureus (clinical isolates) | [125] | – | – |
| Slovakia | B. cereus WSBC 10530, S. aureus ATCC 25923, S. aureus Z MJ346, S. pyogenes Z M494, E. faecalis Z MJ90, L. monocytogenes Z M58, and L. monocytogenes Z M70 | [126] | E. coli ATCC 11229, E. coli O157:H7 Z MJ128, S. typhimurium ATCC 14028, S. enteritidis Z M138, C. coli ATCC 33559, C. coli 2235, C. coli 3341-05, C. jejuni ATCC 33560, C. jejuni NCTC 11168, C. jejuni 375-06, and C. jejuni 3552 | [126] |
| Turkey | S. mutans ATCC 25175, S. aureus 6538-P, S.sobrinus ATCC 33478, S. epidermidis ATCC 12228, E. faecalis ATCC 29212, and M. luteus ATCC 9341 | [127] | P. aeruginosa ATCC 27853, E. coli ATCC 11230, S. Typhimurium CCM 5445, and K. aerogenes ATCC 13048 | [127] |
5. Bacteria as a Source of Alternative or Complementary Treatments (Prebiotics, Probiotics, Symbiotics, and Postbiotics)
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reardon, S. WHO Warns against “post-Antibiotic” Era. Nature 2014, 15, 135–138. [Google Scholar] [CrossRef]
- Kwon, J.H.; Powderly, W.G. The Post-Antibiotic Era Is Here. Science 2021, 373, 471. [Google Scholar] [CrossRef] [PubMed]
- Vinueza, D.; Ochoa-Herrera, V.; Maurice, L.; Tamayo, E.; Mejía, L.; Tejera, E.; Machado, A. Determining the Microbial and Chemical Contamination in Ecuador’s Main Rivers. Sci. Rep. 2021, 11, 17640. [Google Scholar] [CrossRef] [PubMed]
- Montero, L.; Irazabal, J.; Cardenas, P.; Graham, J.P.; Trueba, G. Extended-Spectrum Beta-Lactamase Producing-Escherichia Coli Isolated from Irrigation Waters and Produce in Ecuador. Front. Microbiol. 2021, 12, 709418. [Google Scholar] [CrossRef]
- Cangui-Panchi, S.P.; Ñacato-Toapanta, A.L.; Enríquez-Martínez, L.J.; Reyes, J.; Garzon-Chavez, D.; Machado, A. Biofilm-Forming Microorganisms Causing Hospital-Acquired Infections from Intravenous Catheter: A Systematic Review. Curr. Res. Microb. Sci. 2022, 3, 100175. [Google Scholar] [CrossRef]
- Abe, K.; Nomura, N.; Suzuki, S. Biofilms: Hot Spots of Horizontal Gene Transfer (HGT) in Aquatic Environments, with a Focus on a New HGT Mechanism. FEMS Microbiol. Ecol. 2020, 96, fiaa031. [Google Scholar] [CrossRef]
- Cangui-Panchi, S.P.; Ñacato-Toapanta, A.L.; Enríquez-Martínez, L.J.; Salinas-Delgado, G.A.; Reyes, J.; Garzon-Chavez, D.; Machado, A. Battle Royale: Immune Response on Biofilms—Host-Pathogen Interactions. Curr. Res. Immunol. 2023, 4, 100057. [Google Scholar] [CrossRef]
- Chandra, P.; Unnikrishnan, M.K.; Vandana, K.E.; Mukhopadhyay, C.; Dinesh Acharya, U.; Surulivel Rajan, M.; Rajesh, V. Antimicrobial Resistance and the Post Antibiotic Era: Better Late than Never Effort. Expert. Opin. Drug. Saf. 2021, 20, 1375–1390. [Google Scholar] [CrossRef]
- Hansson, K.; Brenthel, A. Imagining a Post-Antibiotic Era: A Cultural Analysis of Crisis and Antibiotic Resistance. Med. Hum. 2022, 48, 381–388. [Google Scholar] [CrossRef]
- Avershina, E.; Shapovalova, V.; Shipulin, G. Fighting Antibiotic Resistance in Hospital-Acquired Infections: Current State and Emerging Technologies in Disease Prevention, Diagnostics and Therapy. Front. Microbiol. 2021, 12, 2044. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Atiencia-Carrera, M.B.; Cabezas-Mera, F.S.; Tejera, E.; Machado, A. Prevalence of Biofilms in Candida Spp. Bloodstream Infections: A Meta-Analysis. PLoS ONE 2022, 17, e0263522. [Google Scholar] [CrossRef]
- Khan, H.A.; Baig, F.K.; Mehboob, R. Nosocomial Infections: Epidemiology, Prevention, Control and Surveillance. Asian Pac. J. Trop. Biomed. 2017, 7, 478–482. [Google Scholar] [CrossRef]
- Wang, F.; Fu, Y.H.; Sheng, H.J.; Topp, E.; Jiang, X.; Zhu, Y.G.; Tiedje, J.M. Antibiotic Resistance in the Soil Ecosystem: A One Health Perspective. Curr. Opin. Env. Sci. Health 2021, 20, 100230. [Google Scholar] [CrossRef]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic Pollution in the Environment: From Microbial Ecology to Public Policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef] [Green Version]
- Baker, S.; Thomson, N.; Weill, F.X.; Holt, K.E. Genomic Insights into the Emergence and Spread of Antimicrobial-Resistant Bacterial Pathogens. Science 2018, 360, 733–738. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.A.; Ponder, J.B.; Willette, M.; Johnson, T.J.; VanderWaal, K.L. Merging Metagenomics and Spatial Epidemiology To Understand the Distribution of Antimicrobial Resistance Genes from Enterobacteriaceae in Wild Owls. Appl. Environ. Microbiol. 2020, 86, e00571-20. [Google Scholar] [CrossRef]
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Perspective. Microbiol. Spectr. 2018, 6, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Rybak, J.M.; Barker, K.S.; Muñoz, J.F.; Parker, J.E.; Ahmad, S.; Mokaddas, E.; Abdullah, A.; Elhagracy, R.S.; Kelly, S.L.; Cuomo, C.A.; et al. In Vivo Emergence of High-Level Resistance during Treatment Reveals the First Identified Mechanism of Amphotericin B Resistance in Candida Auris. Clin. Microbiol. Infect. 2022, 28, 838–843. [Google Scholar] [CrossRef]
- Muñoz-Barreno, A.; Cabezas-Mera, F.; Tejera, E.; Machado, A. Comparative Effectiveness of Treatments for Bacterial Vaginosis: A Network Meta-Analysis. Antibiotics 2021, 10, 978. [Google Scholar] [CrossRef]
- Dale-Skinner, J.W.; Bonev, B.B. Molecular Mechanisms of Antibiotic Resistance: The Need for Novel Antimicrobial Therapies. In New Strategies Combating Bacterial Infection; Blackwell Publishing Ltd.: Oxford, UK, 2009; pp. 1–46. ISBN 9783527322060. [Google Scholar]
- Chawla, M.; Verma, J.; Gupta, R.; Das, B. Antibiotic Potentiators against Multidrug-Resistant Bacteria: Discovery, Development, and Clinical Relevance. Front. Microbiol. 2022, 13, 887251. [Google Scholar] [CrossRef] [PubMed]
- Klemm, E.J.; Wong, V.K.; Dougan, G. Emergence of Dominant Multidrug-Resistant Bacterial Clades: Lessons from History and Whole-Genome Sequencing. Proc. Natl. Acad. Sci. USA 2018, 115, 12872–12877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial Biofilm and Associated Infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Atiencia-Carrera, M.B.; Cabezas-Mera, F.S.; Vizuete, K.; Debut, A.; Tejera, E.; Machado, A. Evaluation of the Biofilm Life Cycle between Candida Albicans and Candida Tropicalis. Front. Cell. Infect. Microbiol. 2022, 12, 953168. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Bjarnsholt, T. Introduction to Biofilms. In Biofilm Infections; Bjarnsholt, T., Moser, C., Jensen, P.Ø., Høiby, N., Eds.; Springer: New York, NY, USA, 2011; pp. 1–9. ISBN 9781441960832. [Google Scholar]
- Machado, A.; Cerca, N. Influence of Biofilm Formation by Gardnerella Vaginalis and Other Anaerobes on Bacterial Vaginosis. J. Infect. Dis. 2015, 212, 1856–1861. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K. Persister Cells, Dormancy and Infectious Disease. Nat. Rev. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef]
- Rubini, D.; Banu, S.F.; Nisha, P.; Murugan, R.; Thamotharan, S.; Percino, M.J.; Subramani, P.; Nithyanand, P. Essential Oils from Unexplored Aromatic Plants Quench Biofilm Formation and Virulence of Methicillin Resistant Staphylococcus Aureus. Microb. Pathog. 2018, 122, 162–173. [Google Scholar] [CrossRef]
- Simões, M.; Bennett, R.N.; Rosa, E.A.S. Understanding Antimicrobial Activities of Phytochemicals against Multidrug Resistant Bacteria and Biofilms. Nat. Prod. Rep. 2009, 26, 746–757. [Google Scholar] [CrossRef]
- Jubair, N.; Rajagopal, M.; Chinnappan, S.; Abdullah, N.B.; Fatima, A. Review on the Antibacterial Mechanism of Plant-Derived Compounds against Multidrug-Resistant Bacteria (MDR). Evid.-Based Complement. Altern. Med. 2021, 2021, 3663315. [Google Scholar] [CrossRef]
- van Duin, D.; Paterson, D.L. Multidrug-Resistant Bacteria in the Community: An Update. Infect. Dis. Clin. N. Am. 2020, 34, 709–722. [Google Scholar] [CrossRef]
- Serra-Burriel, M.; Keys, M.; Campillo-Artero, C.; Agodi, A.; Barchitta, M.; Gikas, A.; Palos, C.; López-Casasnovas, G. Impact of Multi-Drug Resistant Bacteria on Economic and Clinical Outcomes of Healthcare-Associated Infections in Adults: Systematic Review and Meta-Analysis. PLoS ONE 2020, 15, e0227139. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2023, 12, 116. [Google Scholar] [CrossRef]
- Harbarth, S.; Kahlmeter, G.; Kluytmans, J.; Mendelson, M.; Hospital, G.S.; Town, C.; Africa, S.; Pulcini, C.; Singh, N.; Theuretzbacher, U.; et al. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef]
- Pandey, R.; Mishra, S.K.; Shrestha, A. Characterisation of Eskape Pathogens with Special Reference to Multidrug Resistance and Biofilm Production in a Nepalese Hospital. Infect. Drug. Resist. 2021, 14, 2201–2212. [Google Scholar] [CrossRef]
- Peto, L.; Fawcett, N.J.; Crook, D.W.; Peto, T.E.A.; Llewelyn, M.J.; Walker, A.S. Selective Culture Enrichment and Sequencing of Feces to Enhance Detection of Antimicrobial Resistance Genes in Third-Generation Cephalosporin Resistant Enterobacteriaceae. PLoS ONE 2019, 14, e0222831. [Google Scholar] [CrossRef]
- Cerezales, M.; Ocampo-Sosa, A.A.; Álvarez Montes, L.; Díaz Ríos, C.; Bustamante, Z.; Santos, J.; Martínez-Martínez, L.; Higgins, P.G.; Gallego, L. High Prevalence of Extensively Drug-Resistant Acinetobacter Baumannii at a Children Hospital in Bolivia. Pediatr. Infect. Dis. J. 2018, 37, 1118–1123. [Google Scholar] [CrossRef]
- Santos, A.C.C.; Malta, S.M.; Dantas, R.C.C.; Coelho Rocha, N.D.; Ariston de Carvalho Azevedo, V.; Ueira-Vieira, C. Antimicrobial Activity of Supernatants Produced by Bacteria Isolated from Brazilian Stingless Bee’s Larval Food. BMC Microbiol. 2022, 22, 127. [Google Scholar] [CrossRef]
- Fernandez-Soto, P.; Celi, D.; Tejera, E.; Alvarez-Suarez, J.M.; Machado, A. Cinnamomum Sp. and Pelargonium Odoratissimum as the Main Contributors to the Antibacterial Activity of the Medicinal Drink Horchata: A Study Based on the Antibacterial and Chemical Analysis of 21 Plants. Molecules 2023, 28, 693. [Google Scholar] [CrossRef]
- Qassadi, F.I.; Zhu, Z.; Monaghan, T.M. Plant-Derived Products with Therapeutic Potential against Gastrointestinal Bacteria. Pathogens 2023, 12, 333. [Google Scholar] [CrossRef]
- Pancu, D.F.; Scurtu, A.; Macasoi, I.G.; Marti, D.; Mioc, M.; Soica, C.; Coricovac, D.; Horhat, D.; Poenaru, M.; Dehelean, C. Antibiotics: Conventional Therapy and Natural Compounds with Antibacterial Activity-a Pharmaco-Toxicological Screening. Antibiotics 2021, 10, 401. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Mendoza, L.; Vispo, S.N.; De Lima, L.; Mora, J.R.; Machado, A.; Alexis, F. Hydrogel for the Controlled Delivery of Bioactive Components from Extracts of Eupatorium Glutinosum Lam. Leaves. Molecules 2023, 28, 1591. [Google Scholar] [CrossRef] [PubMed]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically Useful Plant Terpenoids: Biosynthesis, Occurrence, and Mechanism of Action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daglia, M. Polyphenols as Antimicrobial Agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Tatsimo, S.J.N.; de Dieu Tamokou, J.; Havyarimana, L.; Csupor, D.; Forgo, P.; Hohmann, J.; Kuiate, J.-R.; Tane, P. Antimicrobial and Antioxidant Activity of Kaempferol Rhamnoside Derivatives from Bryophyllum Pinnatum. BMC Res. Notes 2012, 5, 158. [Google Scholar] [CrossRef] [Green Version]
- Holler, J.G.; Christensen, S.B.; Slotved, H.-C.; Rasmussen, H.B.; Guzman, A.; Olsen, C.-E.; Petersen, B.; Molgaard, P. Novel Inhibitory Activity of the Staphylococcus Aureus NorA Efflux Pump by a Kaempferol Rhamnoside Isolated from Persea Lingue Nees. J. Antimicrob. Chemother. 2012, 67, 1138–1144. [Google Scholar] [CrossRef] [Green Version]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Fazly Bazzaz, B.S. Review on Plant Antimicrobials: A Mechanistic Viewpoint. Antimicrob. Resist. Infect. Control. 2019, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Duda-Madej, A.; Kozłowska, J.; Krzyżek, P.; Anioł, M.; Seniuk, A.; Jermakow, K.; Dworniczek, E. Antimicrobial O-Alkyl Derivatives of Naringenin and Their Oximes against Multidrug-Resistant Bacteria. Molecules 2020, 25, 3642. [Google Scholar] [CrossRef]
- Chassagne, F.; Samarakoon, T.; Porras, G.; Lyles, J.T.; Dettweiler, M.; Marquez, L.; Salam, A.M.; Shabih, S.; Farrokhi, D.R.; Quave, C.L. A Systematic Review of Plants with Antibacterial Activities: A Taxonomic and Phylogenetic Perspective. Front. Pharm. 2021, 11, 2069. [Google Scholar] [CrossRef]
- CN115708794A; Antibacterial Essential Oil. Tongfu Group China Co. Ltd.: Nantong, China, 2023.
- CN115844777A; Itching-Relieving, Antibacterial and Anti-Inflammatory Composition Containing Plant Extract and Application Thereof. Anhui Chuntang Pharmaceutical Co. Ltd.: Hefei, China, 2022.
- CN115399343B; Plant Composite Antibacterial Agent Containing Peony Extract and Preparation Method and Application Thereof. Shandong Ruiying Pharmaceutical Group Co. Ltd.: Shandong, China, 2022.
- KR102424044B1; Antibacterial Hand Sanitizer Composition Containing Plant Extract. Gusta Co. Ltd.: Seoul, Republic of Korea, 2021.
- CN112868678A; Plant Antibacterial Mite-Killing Agent, Preparation Method Thereof and Daily Necessities Containing Plant Antibacterial Mite-Killing Agent. Guangdong Demay New Materials Technology Co. Ltd.: Shanghai, China, 2021.
- WO2021182661A1; Composition for Improving Antibacterial, Anti-Inflammatory, Antiviral, and Immune Functions, Comprising Extract of Ligularia Stenocephala as Active Ingredient. Dongguk University Gyeongju Campus Industry-Academy Cooperation Foundation: Gyeongju-si, Republic of Korea, 2020.
- US20210386074A1; Plant Extract Compositions and Methods of Making and Using the Same. Greenology Products Inc.: Raleigh, NC, USA, 2020.
- US9138451B2; Plant Extract Hydrolysates and Antibacterial Product Containing the Same. Bionorica SE: Ghent, NY, USA, 2009.
- WO2003035093A1; Antibacterial Composition Comprising Plant Extract. Naturobiotech Co. Ltd.: Suwon, Republic of Korea, 2002.
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsakris, Z.; Rozos, G.; Tsigalou, C.; Bezirtzoglou, E. Interactions between Medical Plant-Derived Bioactive Compounds: Focus on Antimicrobial Combination Effects. Antibiotics 2022, 11, 1014. [Google Scholar] [CrossRef]
- Grecka, K.; Kuś, P.M.; Okińczyc, P.; Worobo, R.W.; Walkusz, J.; Szweda, P. The Anti-Staphylococcal Potential of Ethanolic Polish Propolis Extracts. Molecules 2019, 24, 1732. [Google Scholar] [CrossRef] [Green Version]
- Rybczyńska-Tkaczyk, K.; Grenda, A.; Jakubczyk, A.; Kiersnowska, K.; Bik-Małodzińska, M. Natural Compounds with Antimicrobial Properties in Cosmetics. Pathogens 2023, 12, 320. [Google Scholar] [CrossRef]
- Ilieva, Y.; Marinov, T.; Trayanov, I.; Kaleva, M.; Zaharieva, M.M.; Yocheva, L.; Kokanova-Nedialkova, Z.; Najdenski, H.; Nedialkov, P. Outstanding Antibacterial Activity of Hypericum Rochelii—Comparison of the Antimicrobial Effects of Extracts and Fractions from Four Hypericum Species Growing in Bulgaria with a Focus on Prenylated Phloroglucinols. Life 2023, 13, 274. [Google Scholar] [CrossRef]
- Häsler Gunnarsdottir, S.; Sommerauer, L.; Schnabel, T.; Oostingh, G.J.; Schuster, A. Antioxidative and Antimicrobial Evaluation of Bark Extracts from Common European Trees in Light of Dermal Applications. Antibiotics 2023, 12, 130. [Google Scholar] [CrossRef]
- Aspé, E.; Fernández, K. The Effect of Different Extraction Techniques on Extraction Yield, Total Phenolic, and Anti-Radical Capacity of Extracts from Pinus Radiata Bark. Ind. Crops Prod. 2011, 34, 838–844. [Google Scholar] [CrossRef]
- Raju, D.; Jose, J. Development and Evaluation of Novel Topical Gel of Neem Extract for the Treatment of Bacterial Infections. J. Cosmet. Derm. 2019, 18, 1776–1783. [Google Scholar] [CrossRef]
- Iraqui, P.; Chakraborty, T.; Das, M.K.; Yadav, R.N.S. Herbal Antimicrobial Gel with Leaf Extract of Cassia Alata L. J. Drug Deliv. Ther. 2019, 9, 82–94. [Google Scholar] [CrossRef]
- Popova, T.P.; Ignatov, I.; Petrova, T.E.; Kaleva, M.D.; Huether, F.; Karadzhov, S.D. Antimicrobial Activity In Vitro of Cream from Plant Extracts and Nanosilver, and Clinical Research In Vivo on Veterinary Clinical Cases. Cosmetics 2022, 9, 122. [Google Scholar] [CrossRef]
- Nowak, A.; Zagórska-Dziok, M.; Perużyńska, M.; Cybulska, K.; Kucharska, E.; Ossowicz-Rupniewska, P.; Piotrowska, K.; Duchnik, W.; Kucharski, Ł.; Sulikowski, T.; et al. Assessment of the Anti-Inflammatory, Antibacterial and Anti-Aging Properties and Possible Use on the Skin of Hydrogels Containing Epilobium Angustifolium L. Extracts. Front. Pharm. 2022, 13, 896706. [Google Scholar] [CrossRef]
- Owusu, F.W.A.; Asare, C.O.; Enstie, P.; Adi-Dako, O.; Yeboah, G.N.; Kumadoh, D.; Tetteh-Annor, A.; Amenuke, E.M.; Karen, M. Formulation and in Vitro Evaluation of Oral Capsules and Suspension from the Ethanolic Extract of Cola Nitida Seeds for the Treatment of Diarrhea. Biomed. Res. Int. 2021, 2021, 6630449. [Google Scholar] [CrossRef]
- Chaerunisaa, A.Y.; Susilawati, Y.; Muhaimin, M.; Milanda, T.; Hendriani, R.; Subarnas, A. Antibacterial Activity and Subchronic Toxicity of Cassia Fistula L. Barks in Rats. Toxicol. Rep. 2020, 7, 649–657. [Google Scholar] [CrossRef] [PubMed]
- González-Renteria, M.; del Carmen Monroy-Dosta, M.; Guzmán-García, X.; Hernández-Calderas, I.; Ramos-Lopez, Y.M.A. Antibacterial Activity of Lemna Minor Extracts against Pseudomonas Fluorescens and Safety Evaluation in a Zebrafish Model. Saudi J. Biol. Sci. 2020, 27, 3465–3473. [Google Scholar] [CrossRef] [PubMed]
- Eteraf-Oskouei, T.; Najafi, M. Traditional and Modern Uses of Natural Honey in Human Diseases: A Review. Iran. J. Basic Med. Sci. 2013, 16, 731. [Google Scholar] [PubMed]
- Giampieri, F.; Quiles, J.L.; Cianciosi, D.; Forbes-Hernández, T.Y.; Orantes-Bermejo, F.J.; Alvarez-Suarez, J.M.; Battino, M. Bee Products: An Emblematic Example of Underutilized Sources of Bioactive Compounds. J. Agric. Food Chem. 2022, 70, 6833–6848. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Suarez, J.M. Bee Products—Chemical and Biological Properties; Springer International Publishing: Cham, Switzerland, 2017; ISBN 9783319596891. [Google Scholar]
- Scepankova, H.; Saraiva, J.A.; Estevinho, L.M. Honey Health Benefits and Uses in Medicine. In Bee Products—Chemical and Biological Properties; Springer International Publishing: Cham, Switzerland, 2017; pp. 83–96. [Google Scholar]
- Proaño, A.; Coello, D.; Villacrés-Granda, I.; Ballesteros, I.; Debut, A.; Vizuete, K.; Brenciani, A.; Alvarez-Suarez, J.M. The Osmotic Action of Sugar Combined with Hydrogen Peroxide and Bee-Derived Antibacterial Peptide Defensin-1 Is Crucial for the Antibiofilm Activity of Eucalyptus Honey. LWT 2021, 136, 110379. [Google Scholar] [CrossRef]
- García-Tenesaca, M.; Navarrete, E.S.; Iturralde, G.A.; Villacrés Granda, I.M.; Tejera, E.; Beltrán-Ayala, P.; Giampieri, F.; Battino, M.; Alvarez-Suarez, J.M. Influence of Botanical Origin and Chemical Composition on the Protective Effect against Oxidative Damage and the Capacity to Reduce in Vitro Bacterial Biofilms of Monofloral Honeys from the Andean Region of Ecuador. Int. J. Mol. Sci. 2018, 19, 45. [Google Scholar] [CrossRef] [Green Version]
- Brudzynski, K. Honey as an Ecological Reservoir of Antibacterial Compounds Produced by Antagonistic Microbial Interactions in Plant Nectars, Honey and Honey Bee. Antibiotics 2021, 10, 551. [Google Scholar] [CrossRef]
- Jaktaji, R.P.; Ghalamfarsa, F. Antibacterial Activity of Honeys and Potential Synergism of Honeys with Antibiotics and Alkaloid Extract of Sophora Alopecuroides Plant against Antibiotic-Resistant Escherichia Coli Mutant. Iran. J. Basic. Med. Sci. 2021, 24, 623–628. [Google Scholar] [CrossRef]
- Jaktaji, R.P.; Koochaki, S. In Vitro Activity of Honey, Total Alkaloids of Sophora Alopecuroides and Matrine Alone and in Combination with Antibiotics against Multidrug-Resistant Pseudomonas Aeruginosa Isolates. Lett. Appl. Microbiol. 2022, 75, 70–80. [Google Scholar] [CrossRef]
- Stefanis, C.; Stavropoulou, E.; Giorgi, E.; Voidarou, C.; Constantinidis, T.C.; Vrioni, G.; Tsakris, A. Honey’s Antioxidant and Antimicrobial Properties: A Bibliometric Study. Antioxidants 2023, 12, 414. [Google Scholar] [CrossRef]
- Roshan, N.; Rippers, T.; Locher, C.; Hammer, K.A. Antibacterial Activity and Chemical Characteristics of Several Western Australian Honeys Compared to Manuka Honey and Pasture Honey. Arch. Microbiol. 2017, 199, 347–355. [Google Scholar] [CrossRef]
- Lusby, P.E.; Coombes, A.L.; Wilkinson, J.M. Bactericidal Activity of Different Honeys against Pathogenic Bacteria. Arch. Med. Res. 2005, 36, 464–467. [Google Scholar] [CrossRef]
- Zapata-Vahos, I.C.; Henao-Rojas, J.C.; Yepes-Betancur, D.P.; Marín-Henao, D.; Giraldo Sánchez, C.E.; Calvo-Cardona, S.J.; David, D.; Quijano-Abril, M. Physicochemical Parameters, Antioxidant Capacity, and Antimicrobial Activity of Honeys from Tropical Forests of Colombia: Apis Mellifera and Melipona Eburnea. Foods 2023, 12, 1001. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Tulipani, S.; Díaz, D.; Estevez, Y.; Romandini, S.; Giampieri, F.; Damiani, E.; Astolfi, P.; Bompadre, S.; Battino, M. Antioxidant and Antimicrobial Capacity of Several Monofloral Cuban Honeys and Their Correlation with Color, Polyphenol Content and Other Chemical Compounds. Food Chem. Toxicol. 2010, 48, 2490–2499. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Giampieri, F.; Brenciani, A.; Mazzoni, L.; Gasparrini, M.; González-Paramás, A.M.; Santos-Buelga, C.; Morroni, G.; Simoni, S.; Forbes-Hernández, T.Y.; et al. Apis Mellifera vs Melipona Beecheii Cuban Polifloral Honeys: A Comparison Based on Their Physicochemical Parameters, Chemical Composition and Biological Properties. LWT 2018, 87, 272–279. [Google Scholar] [CrossRef]
- Morroni, G.; Alvarez-Suarez, J.M.; Brenciani, A.; Simoni, S.; Fioriti, S.; Pugnaloni, A.; Giampieri, F.; Mazzoni, L.; Gasparrini, M.; Marini, E.; et al. Comparison of the Antimicrobial Activities of Four Honeys from Three Countries (New Zealand, Cuba, and Kenya). Front. Microbiol. 2018, 9, 1378. [Google Scholar] [CrossRef] [Green Version]
- Valdés-Silverio, L.A.; Iturralde, G.; García-Tenesaca, M.; Paredes-Moreta, J.; Narváez-Narváez, D.A.; Rojas-Carrillo, M.; Tejera, E.; Beltrán-Ayala, P.; Giampieri, F.; Alvarez-Suarez, J.M. Physicochemical Parameters, Chemical Composition, Antioxidant Capacity, Microbial Contamination and Antimicrobial Activity of Eucalyptus Honey from the Andean Region of Ecuador. J. Apic. Res. 2018, 57, 382–394. [Google Scholar] [CrossRef]
- Villacrés-Granda, I.; Proaño, A.; Coello, D.; Debut, A.; Vizuete, K.; Ballesteros, I.; Granda-Albuja, G.; Rosero-Mayanquer, H.; Battino, M.; Giampieri, F.; et al. Effect of Thermal Liquefaction on Quality, Chemical Composition and Antibiofilm Activity against Multiresistant Human Pathogens of Crystallized Eucalyptus Honey. Food Chem. 2021, 365, 130519. [Google Scholar] [CrossRef]
- Leyva-Jimenez, F.J.; Lozano-Sanchez, J.; Borras-Linares, I.; de la Luz Cadiz-Gurrea, M.; Mahmoodi-Khaledi, E. Potential Antimicrobial Activity of Honey Phenolic Compounds against Gram Positive and Gram Negative Bacteria. LWT 2019, 101, 236–245. [Google Scholar] [CrossRef]
- Coniglio, M.A.; Laganà, P.; Faro, G.; Marranzano, M. Antimicrobial Potential of Sicilian Honeys against Staphylococcus Aureus and Pseudomonas Aeruginosa. J. AOAC Int. 2018, 101, 956–959. [Google Scholar] [CrossRef]
- Aumeeruddy, M.Z.; Aumeeruddy-Elalfi, Z.; Neetoo, H.; Zengin, G.; Blom van Staden, A.; Fibrich, B.; Lambrechts, I.A.; Rademan, S.; Szuman, K.M.; Lall, N.; et al. Pharmacological Activities, Chemical Profile, and Physicochemical Properties of Raw and Commercial Honey. Biocatal. Agric. Biotechnol. 2019, 18, 101005. [Google Scholar] [CrossRef]
- Lu, J.; Turnbull, L.; Burke, C.M.; Liu, M.; Carter, D.A.; Schlothauer, R.C.; Whitchurch, C.B.; Harry, E.J. Manuka-Type Honeys Can Eradicate Biofilms Produced by Staphylococcus Aureus Strains with Different Biofilm-Forming Abilities. PeerJ 2014, 2, e326. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, H.; Sultan, M.; Qamar, M.U.; Junaid, K.; Rasool, N.; Alanazi, A.; Alruways, M.W.; Mazhari, B.B.Z.; Alruwaili, Y.; Bukhari, S.N.A.; et al. Antibacterial Efficacy of Indigenous Pakistani Honey against Extensively Drug-Resistant Clinical Isolates of Salmonella Enterica Serovar Typhi: An Alternative Option to Combat Antimicrobial Resistance. BMC Complement. Med. 2023, 23, 42. [Google Scholar] [CrossRef] [PubMed]
- Kunat-Budzyńska, M.; Rysiak, A.; Wiater, A.; Grąz, M.; Andrejko, M.; Budzyński, M.; Bryś, M.S.; Sudziński, M.; Tomczyk, M.; Gancarz, M.; et al. Chemical Composition and Antimicrobial Activity of New Honey Varietals. Int. J. Env. Res. Public Health 2023, 20, 2458. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J.; Bohova, J.; Horniackova, M.; Klaudiny, J.; Majtan, V. Anti-Biofilm Effects of Honey against Wound Pathogens Proteus mirabilis and Enterobacter cloacae. Phytother. Res. 2014, 28, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Bucekova, M.; Jardekova, L.; Juricova, V.; Bugarova, V.; Di Marco, G.; Gismondi, A.; Leonardi, D.; Farkasovska, J.; Godocikova, J.; Laho, M.; et al. Antibacterial Activity of Different Blossom Honeys: New Findings. Molecules 2019, 24, 1573. [Google Scholar] [CrossRef] [Green Version]
- Combarros-Fuertes, P.; Estevinho, L.M.; Dias, L.G.; Castro, J.M.; Tomás-Barberán, F.A.; Tornadijo, M.E.; Fresno-Baro, J.M. Bioactive Components and Antioxidant and Antibacterial Activities of Different Varieties of Honey: A Screening Prior to Clinical Application. J. Agric. Food Chem. 2019, 67, 688–698. [Google Scholar] [CrossRef] [Green Version]
- Kolayli, S.; Kazaz, G.; Özkök, A.; Keskin, M.; Kara, Y.; Demir Kanbur, E.; Ertürk, Ö. The Phenolic Composition, Aroma Compounds, Physicochemical and Antimicrobial Properties of Nigella sativa L. (Black Cumin) Honey. Eur. Food Res. Technol. 2023, 249, 653–664. [Google Scholar] [CrossRef]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [Green Version]
- Sforcin, J.M.; Bankova, V. Propolis: Is There a Potential for the Development of New Drugs? J. Ethnopharmacol. 2011, 133, 253–260. [Google Scholar] [CrossRef]
- Sforcin, J.M. Biological Properties and Therapeutic Applications of Propolis. Phytother. Res. 2016, 30, 894–905. [Google Scholar] [CrossRef]
- Pimenta, H.C.; Violante, I.M.P.; de Musis, C.R.; Borges, Á.H.; Aranha, A.M.F. In Vitro Effectiveness of Brazilian Brown Propolis against Enterococcus Faecalis. Braz. Oral. Res. 2015, 29, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Viuda-Martos, M.; Ruiz-Navajas, Y.; Fernández-López, J.; Pérez-Álvarez, J.A. Functional Properties of Honey, Propolis, and Royal Jelly. J. Food Sci. 2008, 73, R117–R124. [Google Scholar] [CrossRef]
- Seibert, J.B.; Bautista-Silva, J.P.; Amparo, T.R.; Petit, A.; Pervier, P.; dos Santos Almeida, J.C.; Azevedo, M.C.; Silveira, B.M.; Brandão, G.C.; de Souza, G.H.B.; et al. Development of Propolis Nanoemulsion with Antioxidant and Antimicrobial Activity for Use as a Potential Natural Preservative. Food Chem. 2019, 287, 61–67. [Google Scholar] [CrossRef]
- Massaro, C.F.; Simpson, J.B.; Powell, D.; Brooks, P. Chemical Composition and Antimicrobial Activity of Honeybee (Apis Mellifera Ligustica) Propolis from Subtropical Eastern Australia. Sci. Nat. 2015, 102, 68. [Google Scholar] [CrossRef]
- Bittencourt, M.L.F.; Ribeiro, P.R.; Franco, R.L.P.; Hilhorst, H.W.M.; de Castro, R.D.; Fernandez, L.G. Metabolite Profiling, Antioxidant and Antibacterial Activities of Brazilian Propolis: Use of Correlation and Multivariate Analyses to Identify Potential Bioactive Compounds. Food Res. Int. 2015, 76, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Moncla, B.J.; Guevara, P.W.; Wallace, J.A.; Marcucci, M.C.; Nor, J.E.; Bretz, W.A. The Inhibitory Activity of Typified Propolis against. Enterococcus Species. Z. Nat. C 2012, 67, 249–256. [Google Scholar] [CrossRef]
- Schmidt, E.M.; Stock, D.; Chada, F.J.G.; Finger, D.; Christine Helena Frankland Sawaya, A.; Eberlin, M.N.; Felsner, M.L.; Quináia, S.P.; Monteiro, M.C.; Torres, Y.R. A Comparison between Characterization and Biological Properties of Brazilian Fresh and Aged Propolis. Biomed. Res. Int. 2014, 2014, 257617. [Google Scholar] [CrossRef] [Green Version]
- Campos, J.F.; dos Santos, U.P.; dos Santos da Rocha, P.; Damião, M.J.; Balestieri, J.B.P.; Cardoso, C.A.L.; Paredes-Gamero, E.J.; Estevinho, L.M.; de Picoli Souza, K.; dos Santos, E.L. Antimicrobial, Antioxidant, Anti-Inflammatory, and Cytotoxic Activities of Propolis from the Stingless Bee Tetragonisca fiebrigi (Jataí). Evid.-Based Complement. Altern. Med. 2015, 2015, 296186. [Google Scholar] [CrossRef] [Green Version]
- Regueira, M.S.; Tintino, S.R.; da Silva, A.R.P.; do Socorro Costa, M.; Boligon, A.A.; Matias, E.F.F.; de Queiroz Balbino, V.; Menezes, I.R.A.; Melo Coutinho, H.D. Seasonal Variation of Brazilian Red Propolis: Antibacterial Activity, Synergistic Effect and Phytochemical Screening. Food Chem. Toxicol. 2017, 107, 572–580. [Google Scholar] [CrossRef]
- Dantas Silva, R.P.; Machado, B.A.S.; de Abreu Barreto, G.; Costa, S.S.; Andrade, L.N.; Amaral, R.G.; Carvalho, A.A.; Padilha, F.F.; Barbosa, J.D.V.; Umsza-Guez, M.A. Antioxidant, Antimicrobial, Antiparasitic, and Cytotoxic Properties of Various Brazilian Propolis Extracts. PLoS ONE 2017, 12, e0172585. [Google Scholar] [CrossRef] [PubMed]
- Popova, M.; Dimitrova, R.; Al-Lawati, H.T.; Tsvetkova, I.; Najdenski, H.; Bankova, V. Omani Propolis: Chemical Profiling, Antibacterial Activity and New Propolis Plant Sources. Chem. Cent. J. 2013, 7, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridi, R.; Montenegro, G.; Nuñez-Quijada, G.; Giordano, A.; Fernanda Morán-Romero, M.; Jara-Pezoa, I.; Speisky, H.; Atala, E.; López-Alarcón, C. International Regulations of Propolis Quality: Required Assays Do Not Necessarily Reflect Their Polyphenolic-Related In Vitro Activities. J. Food Sci. 2015, 80, C1188–C1195. [Google Scholar] [CrossRef] [PubMed]
- AL-Ani, I.; Zimmermann, S.; Reichling, J.; Wink, M. Antimicrobial Activities of European Propolis Collected from Various Geographic Origins Alone and in Combination with Antibiotics. Medicines 2018, 5, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, M.P.; Chinou, I.B.; Marekov, I.N.; Bankova, V.S. Terpenes with Antimicrobial Activity from Cretan Propolis. Phytochemistry 2009, 70, 1262–1271. [Google Scholar] [CrossRef]
- Patel, J.; Ketkar, S.; Patil, S.; Fearnley, J.; Mahadik, K.R.; Paradkar, A.R. Potentiating Antimicrobial Efficacy of Propolis through Niosomal-Based System for Administration. Integr. Med. Res. 2015, 4, 94–101. [Google Scholar] [CrossRef] [Green Version]
- De Marco, S.; Piccioni, M.; Pagiotti, R.; Pietrella, D. Antibiofilm and Antioxidant Activity of Propolis and Bud Poplar Resins versus Pseudomonas Aeruginosa. Evid.-Based Complement. Altern. Med. 2017, 2017, 5163575. [Google Scholar] [CrossRef] [Green Version]
- Campana, R.; Patrone, V.; Franzini, I.T.M.; Diamantini, G.; Vittoria, E.; Baffone, W. Antimicrobial Activity of Two Propolis Samples against Human Campylobacter jejuni. J. Med. Food 2009, 12, 1050–1056. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, C.S.; Kim, B.-H.; Ro, S.-B.; Lim, Y.K.; Park, S.-N.; Cho, E.; Ko, J.-H.; Kwon, S.-S.; Ko, Y.-M.; et al. Antimicrobial Effect of Korean Propolis against the Mutans Streptococci Isolated from Korean. J. Microbiol. 2011, 49, 161–164. [Google Scholar] [CrossRef]
- El-Guendouz, S.; Aazza, S.; Lyoussi, B.; Bankova, V.; Popova, M.; Neto, L.; Faleiro, M.L.; da Graça Miguel, M. Moroccan Propolis: A Natural Antioxidant, Antibacterial, and Antibiofilm against Staphylococcus aureus with No Induction of Resistance after Continuous Exposure. Evid.-Based Complement. Altern. Med. 2018, 2018, 9759240. [Google Scholar] [CrossRef] [Green Version]
- Wojtyczka, R.; Dziedzic, A.; Idzik, D.; Kępa, M.; Kubina, R.; Kabała-Dzik, A.; Smoleń-Dzirba, J.; Stojko, J.; Sajewicz, M.; Wąsik, T. Susceptibility of Staphylococcus Aureus Clinical Isolates to Propolis Extract Alone or in Combination with Antimicrobial Drugs. Molecules 2013, 18, 9623–9640. [Google Scholar] [CrossRef] [Green Version]
- Mavri, A.; Abramovič, H.; Polak, T.; Bertoncelj, J.; Jamnik, P.; Smole Možina, S.; Jeršek, B. Chemical Properties and Antioxidant and Antimicrobial Activities of Slovenian Propolis. Chem. Biodivers. 2012, 9, 1545–1558. [Google Scholar] [CrossRef]
- Uzel, A.; Sorkun, K.; Önçağ, Ö.; Çoğulu, D.; Gençay, Ö.; Salïh, B. Chemical Compositions and Antimicrobial Activities of Four Different Anatolian Propolis Samples. Microbiol. Res. 2005, 160, 189–195. [Google Scholar] [CrossRef]
- Ötleş, S. Probiotics and Prebiotics in Food, Nutrition and Health; Taylor & Francis Group: Boca Raton, FL, USA, 2014; ISBN 9781466586246. [Google Scholar]
- de Melo, F.H.C.; Menezes, F.N.D.D.; de Sousa, J.M.B.; dos Santos Lima, M.; da Silva Campelo Borges, G.; de Souza, E.L.; Magnani, M. Prebiotic Activity of Monofloral Honeys Produced by Stingless Bees in the Semi-Arid Region of Brazilian Northeastern toward Lactobacillus Acidophilus LA-05 and Bifidobacterium Lactis BB-12. Food Res. Int. 2020, 128, 108809. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.H. The Microbiota of the Vagina and Its Influence on Women’s Health and Disease. Am. J. Med. Sci. 2012, 343, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Suez, J.; Zmora, N.; Elinav, E. Probiotics in the Next-Generation Sequencing Era. Gut Microbes 2020, 11, 77–93. [Google Scholar] [CrossRef]
- Wieërs, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; de Foy, J.-M.P.; Dequenne, I.; de Timary, P.; Cani, P.D. How Probiotics Affect the Microbiota. Front. Cell. Infect. Microbiol. 2020, 9, 454. [Google Scholar] [CrossRef] [Green Version]
- Abdou, A.M.; Hedia, R.H.; Omara, S.T.; Mahmoud, M.A.E.F.; Kandil, M.M.; Bakry, M.A. Interspecies Comparison of Probiotics Isolated from Different Animals. Vet. World 2018, 11, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Zangl, I.; Pap, I.J.; Aspöck, C.; Schüller, C. The Role of Lactobacillus Species in the Control of Candida via Biotrophic Interactions. Microb. Cell 2020, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pacha-Herrera, D.; Erazo-Garcia, M.P.; Cueva, D.F.; Orellana, M.; Borja-Serrano, P.; Arboleda, C.; Tejera, E.; Machado, A. Clustering Analysis of the Multi-Microbial Consortium by Lactobacillus Species against Vaginal Dysbiosis Among Ecuadorian Women. Front. Cell. Infect. Microbiol. 2022, 12, 863208. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Arias, R.J.; Guachi-Álvarez, B.O.; Montalvo-Vivero, D.E.; Machado, A. Lactobacilli Displacement and Candida Albicans Inhibition on Initial Adhesion Assays: A Probiotic Analysis. BMC Res. Notes 2022, 15, 239. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.Y.; Cheah, Y.K.; Seow, H.F.; Sandai, D.; Than, L.T.L. Probiotic Lactobacillus Rhamnosus GR-1 and Lactobacillus Reuteri RC-14 Exhibit Strong Antifungal Effects against Vulvovaginal Candidiasis-Causing Candida Glabrata Isolates. J. Appl. Microbiol. 2015, 118, 1180–1190. [Google Scholar] [CrossRef] [Green Version]
- Chelliah, R.; Kim, E.J.; Daliri, E.B.M.; Antony, U.; Oh, D.H. In Vitro Probiotic Evaluation of Saccharomyces boulardii with Antimicrobial Spectrum in a Caenorhabditis elegans Model. Foods 2021, 10, 1428. [Google Scholar] [CrossRef]
- Zulkhairi Amin, F.A.; Sabri, S.; Ismail, M.; Chan, K.W.; Ismail, N.; Mohd Esa, N.; Mohd Lila, M.A.; Zawawi, N. Probiotic Properties of Bacillus Strains Isolated from Stingless Bee (Heterotrigona itama) Honey Collected across Malaysia. Int. J. Environ. Res. Public Health 2019, 17, 278. [Google Scholar] [CrossRef] [Green Version]
- Dyshlyuk, L.S.; Milentyeva, I.S.; Asyakina, L.K.; Ostroumov, L.A.; Osintsev, A.M.; Pozdnyakova, A.V. Using Bifidobacterium and Propionibacterium Strains in Probiotic Consortia to Normalize the Gastrointestinal Tract. Braz. J. Biol. 2022, 84, 1–17. [Google Scholar] [CrossRef]
- Luiz, F.; Do Carmo, R.; Rabah, H.; Fernandes Cordeiro, B.; Da Silva, H.S.; Pessoa, R.M.; Odília, S.; Fernandes, A.; Cardoso, V.N.; Gagnaire, V.; et al. Probiotic Propionibacterium Freudenreichii Requires SlpB Protein to Mitigate Mucositis Induced by Chemotherapy. Oncotarget 2020, 10, 7198–7219. [Google Scholar]
- Di Cerbo, A.; Palmieri, B.; Aponte, M.; Morales-Medina, J.C.; Iannitti, T. Mechanisms and Therapeutic Effectiveness of Lactobacilli. J. Clin. Pathol. 2016, 69, 187–203. [Google Scholar] [CrossRef] [Green Version]
- Hefzy, E.M.; Khalil, M.A.F.; Ibrahim Amin, A.A.; Ashour, H.M.; Abdelaliem, Y.F. Bacteriocin-like Inhibitory Substances from Probiotics as Therapeutic Agents for Candida Vulvovaginitis. Antibiotics 2021, 10, 306. [Google Scholar] [CrossRef]
- Dos Santos, C.I.; França, Y.R.; Campos, C.D.L.; Bomfim, M.R.Q.; Melo, B.O.; Holanda, R.A.; Santos, V.L.; Monteiro, S.G.; Moffa, E.B.; Monteiro, A.S.; et al. Antifungal and Antivirulence Activity of Vaginal Lactobacillus Spp. Products against Candida Vaginal Isolates. Pathogens 2019, 8, 150. [Google Scholar] [CrossRef] [Green Version]
- Anjana; Tiwari, S.K. Bacteriocin-Producing Probiotic Lactic Acid Bacteria in Controlling Dysbiosis of the Gut Microbiota. Front. Cell. Infect. Microbiol. 2022, 12, 415. [Google Scholar] [CrossRef]
- Allonsius, C.N.; van den Broek, M.F.L.; De Boeck, I.; Kiekens, S.; Oerlemans, E.F.M.; Kiekens, F.; Foubert, K.; Vandenheuvel, D.; Cos, P.; Delputte, P.; et al. Interplay between Lactobacillus Rhamnosus GG and Candida and the Involvement of Exopolysaccharides. Microb. Biotechnol. 2017, 10, 1753–1763. [Google Scholar] [CrossRef]
- Matsubara, V.H.; Wang, Y.; Bandara, H.M.H.N.; Mayer, M.P.A.; Samaranayake, L.P. Probiotic Lactobacilli Inhibit Early Stages of Candida Albicans Biofilm Development by Reducing Their Growth, Cell Adhesion, and Filamentation. Appl. Microbiol. Biotechnol. 2016, 100, 6415–6426. [Google Scholar] [CrossRef] [Green Version]
- Mendling, W. Microbiota of the Human Body. Adv. Exp. Med. Biol. 2016, 902, 83–93. [Google Scholar] [CrossRef]
- Carlson, J.L.; Erickson, J.M.; Lloyd, B.B.; Slavin, J.L. Health Effects and Sources of Prebiotic Dietary Fiber. Curr. Dev. Nutr. 2018, 2, nzy005. [Google Scholar] [CrossRef] [Green Version]
- Alexander, C.; Swanson, K.S.; Fahey, G.C.; Garleb, K.A. Perspective: Physiologic Importance of Short-Chain Fatty Acids from Nondigestible Carbohydrate Fermentation. Adv. Nutr. 2019, 10, 576–589. [Google Scholar] [CrossRef] [Green Version]
- Abouloifa, H.; Khodaei, N.; Rokni, Y.; Karboune, S.; Brasca, M.; D’Hallewin, G.; Salah, R.B.; Saalaoui, E.; Asehraou, A. The Prebiotics (Fructo-Oligosaccharides and Xylo-Oligosaccharides) Modulate the Probiotic Properties of Lactiplantibacillus and Levilactobacillus Strains Isolated from Traditional Fermented Olive. World J. Microbiol. Biotechnol. 2020, 36, 185. [Google Scholar] [CrossRef]
- Roupar, D.; Coelho, M.C.; Gonçalves, D.A.; Silva, S.P.; Coelho, E.; Silva, S.; Coimbra, M.A.; Pintado, M.; Teixeira, J.A.; Nobre, C. Evaluation of Microbial-Fructo-Oligosaccharides Metabolism by Human Gut Microbiota Fermentation as Compared to Commercial Inulin-Derived Oligosaccharides. Foods 2022, 11, 954. [Google Scholar] [CrossRef]
- Mounir, M.; Ibijbijen, A.; Farih, K.; Rabetafika, H.N.; Razafindralambo, H.L. Synbiotics and Their Antioxidant Properties, Mechanisms, and Benefits on Human and Animal Health: A Narrative Review. Biomolecules 2022, 12, 1443. [Google Scholar] [CrossRef]
- Gaucher, F.; Bonnassie, S.; Rabah, H.; Marchand, P. Review: Adaptation of Beneficial Propionibacteria, Lactobacilli, and Bifidobacteria Improves Tolerance Toward Technological and Digestive Stresses. Front. Microbiol. 2019, 10, 841. [Google Scholar] [CrossRef] [PubMed]
- Megur, A.; Daliri, E.B.-M.; Baltriukienė, D.; Burokas, A. Prebiotics as a Tool for the Prevention and Treatment of Obesity and Diabetes: Classification and Ability to Modulate the Gut Microbiota. Int. J. Mol. Sci. 2022, 23, 6097. [Google Scholar] [CrossRef] [PubMed]
- Herbst, T.; Sichelstiel, A.; Schär, C.; Yadava, K.; Bürki, K.; Cahenzli, J.; McCoy, K.; Marsland, B.J.; Harris, N.L. Dysregulation of Allergic Airway Inflammation in the Absence of Microbial Colonization. Am. J. Respir. Crit. Care Med. 2011, 184, 198–205. [Google Scholar] [CrossRef]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Björkstén, B.; Engstrand, L.; Jenmalm, M.C. Low Diversity of the Gut Microbiota in Infants with Atopic Eczema. J. Allergy Clin. Immunol. 2012, 129, 434–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylund, L.; Satokari, R.; Nikkilä, J.; Rajilić-Stojanović, M.; Kalliomäki, M.; Isolauri, E.; Salminen, S.; de Vos, W.M. Microarray Analysis Reveals Marked Intestinal Microbiota Aberrancy in Infants Having Eczema Compared to Healthy Children in At-Risk for Atopic Disease. BMC Microbiol. 2013, 13, 12. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, M.; Oomah, B.D.; Oliveira, W.P.; Burgos-díaz, C.; Rubilar, M.; Shene, C. Probiotics and Prebiotics Potential for the Care of Skin, Female Urogenital Tract, and Respiratory Tract. Folia Microbiol. 2020, 65, 245–264. [Google Scholar] [CrossRef] [Green Version]
- Nambiar, R.B.; Perumal, A.B.; Shittu, T.; Sadiku, E.R.; Sellamuthu, P.S. Editorial: Probiotics, Prebiotics, Synbiotics, Postbiotics, & Paraprobiotics—New Perspective for Functional Foods and Nutraceuticals. Front. Nutr. 2023, 10, 1164676. [Google Scholar] [CrossRef]
- Pu, J.; Hang, S.; Liu, M.; Chen, Z.; Xiong, J.; Li, Y.; Wu, H.; Zhao, X.; Liu, S.; Gu, Q.; et al. A Class IIb Bacteriocin Plantaricin NC8 Modulates Gut Microbiota of Different Enterotypes In Vitro. Front. Nutr. 2022, 9, 877948. [Google Scholar] [CrossRef]
- Sornsenee, P.; Chatatikun, M.; Mitsuwan, W.; Kongpol, K.; Kooltheat, N.; Sohbenalee, S.; Pruksaphanrat, S.; Mudpan, A.; Romyasamit, C. Lyophilized Cell-Free Supernatants of Lactobacillus Isolates Exhibited Antibiofilm, Antioxidant, and Reduces Nitric Oxide Activity in Lipopolysaccharide- Stimulated RAW 264.7 Cells. PeerJ 2021, 9, e12586. [Google Scholar] [CrossRef]
- Scarpellini, E.; Rinninella, E.; Basilico, M.; Colomier, E.; Rasetti, C.; Larussa, T.; Santori, P.; Abenavoli, L. From Pre-and Probiotics to Post-Biotics: A Narrative Review. Int. J. Env. Res. Public. Health 2022, 19, 37. [Google Scholar] [CrossRef]
- Raman, M.; Ambalam, P.; Kondepudi, K.K.; Pithva, S.; Kothari, C.; Patel, A.T.; Purama, R.K.; Dave, J.M.; Vyas, B.R.M. Potential of Probiotics, Prebiotics and Synbiotics for Management of Colorectal Cancer. Gut Microbes 2013, 4, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Hong, L.; Lee, S.M.; Kim, W.S.; Choi, Y.J.; Oh, S.H.; Li, Y.L.; Choi, S.H.; Chung, D.H.; Jung, E.; Kang, S.K.; et al. Synbiotics Containing Nanoprebiotics: A Novel Therapeutic Strategy to Restore Gut Dysbiosis. Front. Microbiol. 2021, 12, 715241. [Google Scholar] [CrossRef]
- Bandyopadhyay, B.; Das, S.; Mitra, P.K.; Kundu, A.; Mandal, V.; Adhikary, R.; Mandal, V.; Mandal, N.C. Characterization of Two New Strains of Lactococcus Lactis for Their Probiotic Efficacy over Commercial Synbiotics Consortia. Braz. J. Microbiol. 2022, 53, 903–920. [Google Scholar] [CrossRef]
- Polakowski, C.B.; Kato, M.; Preti, V.B.; Schieferdecker, M.E.M.; Ligocki Campos, A.C. Impact of the Preoperative Use of Synbiotics in Colorectal Cancer Patients: A Prospective, Randomized, Double-Blind, Placebo-Controlled Study. Nutrition 2019, 58, 40–46. [Google Scholar] [CrossRef]
- Sergeev, I.N.; Aljutaily, T.; Walton, G.; Huarte, E. Effects of Synbiotic Supplement on Human Gut Microbiota, Body Composition and Weight Loss in Obesity. Nutrients 2020, 12, 222. [Google Scholar] [CrossRef] [Green Version]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Thorakkattu, P.; Khanashyam, A.C.; Shah, K.; Babu, K.S.; Mundanat, A.S.; Deliephan, A.; Deokar, G.S.; Santivarangkna, C.; Nirmal, N.P. Postbiotics: Current Trends in Food and Pharmaceutical Industry. Foods 2022, 11, 3094. [Google Scholar] [CrossRef]
- Cuevas-González, P.F.; Liceaga, A.M.; Aguilar-Toalá, J.E. Postbiotics and Paraprobiotics: From Concepts to Applications. Food Res. Int. 2020, 136, 109502. [Google Scholar] [CrossRef]
- Overmyer, K.A.; Rhoads, T.W.; Merrill, A.E.; Ye, Z.; Westphall, M.S.; Acharya, A.; Shukla, S.K.; Coon, J.J. Proteomics, Lipidomics, Metabolomics, and 16S DNA Sequencing of Dental Plaque from Patients with Diabetes and Periodontal Disease. Mol. Cell. Proteom. 2021, 20, 100126. [Google Scholar] [CrossRef]
- Mayorgas, A.; Dotti, I.; Salas, A. Microbial Metabolites, Postbiotics, and Intestinal Epithelial Function. Mol. Nutr. Food Res. 2021, 65, 2000188. [Google Scholar] [CrossRef]
- Joshi, C.; Patel, P.; Kothari, V. Anti-Infective Potential of Hydroalcoholic Extract of Punica Granatum Peel against Gram-Negative Bacterial Pathogens. F1000Research 2019, 8, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandri, G.; Milani, C.; Duranti, S.; Mancabelli, L.; Ranjanoro, T.; Modica, S.; Carnevali, L.; Statello, R.; Bottacini, F.; Turroni, F.; et al. Ability of Bifidobacteria to Metabolize Chitin-Glucan and Its Impact on the Gut Microbiota. Sci. Rep. 2019, 9, 5755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.; Ma, Y.; Yang, S.; Zhang, S.; Liu, S.; Xiao, J.; Wang, Y.; Wang, W.; Yang, H.; Li, S.; et al. Gut Microbiota-Derived Ursodeoxycholic Acid from Neonatal Dairy Calves Improves Intestinal Homeostasis and Colitis to Attenuate Extended-Spectrum β-Lactamase-Producing Enteroaggregative Escherichia Coli Infection. Microbiome 2022, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Kao, H.J.; Balasubramaniam, A.; Chen, C.C.; Huang, C.M. Extracellular Electrons Transferred from Honey Probiotic Bacillus Circulans Inhibits Inflammatory Acne Vulgaris. Sci. Rep. 2022, 12, 19217. [Google Scholar] [CrossRef]
- Islam, M.I.; Seo, H.; Redwan, A.; Kim, S.; Lee, S.; Siddiquee, M.; Song, H.Y. In Vitro and In Vivo Anti-Clostridioides Difficile Effect of a Probiotic Bacillus Amyloliquefaciens Strain. J. Microbiol. Biotechnol. 2022, 32, 46–55. [Google Scholar] [CrossRef]
- Ishnaiwer, M.; Bezabih, Y.; Javaudin, F.; Sassi, M.; Bemer, P.; Batard, E.; Dion, M. In Vitro and In Vivo Activity of New Strains of Bacillus Subtilis against ESBL-Producing Escherichia Coli: An Experimental Study. J. Appl. Microbiol. 2022, 132, 2270–2279. [Google Scholar] [CrossRef]
- Cui, L.H.; Yan, C.G.; Li, H.S.; Kim, W.S.; Hong, L.; Kang, S.K.; Choi, Y.J.; Cho, C.S. A New Method of Producing a Natural Antibacterial Peptide by Encapsulated Probiotics Internalized with Inulin Nanoparticles as Prebiotics. J. Microbiol. Biotechnol. 2018, 28, 510–519. [Google Scholar] [CrossRef]
- Hashem, N.M.; Hosny, N.S.; El-Desoky, N.I.; Shehata, M.G. Effect of Nanoencapsulated Alginate-Synbiotic on Gut Microflora Balance, Immunity, and Growth Performance of Growing Rabbits. Polymers 2021, 13, 4191. [Google Scholar] [CrossRef]
- Jung, Y.J.; Kim, H.S.; Jaygal, G.; Cho, H.R.; Lee, K.B.; Song, I.B.; Kim, J.H.; Kwak, M.S.; Han, K.H.; Bae, M.J.; et al. Postbiotics Enhance NK Cell Activation in Stress-Induced Mice through Gut Microbiome Regulation. J. Microbiol. Biotechnol. 2022, 32, 612–620. [Google Scholar] [CrossRef]
- Golkar, N.; Ashoori, Y.; Heidari, R.; Omidifar, N.; Abootalebi, S.N.; Mohkam, M.; Gholami, A. A Novel Effective Formulation of Bioactive Compounds for Wound Healing: Preparation, In Vivo Characterization, and Comparison of Various Postbiotics Cold Creams in a Rat Model. Evid.-Based Complement. Altern. Med. 2021, 2021, 8577116. [Google Scholar] [CrossRef]
- Puccetti, M.; Gomes dos Reis, L.; Pariano, M.; Costantini, C.; Renga, G.; Ricci, M.; Traini, D.; Giovagnoli, S. Development and In Vitro-In Vivo Performances of an Inhalable Indole-3-Carboxaldehyde Dry Powder to Target Pulmonary Inflammation and Infection. Int. J. Pharm. 2021, 607, 121004. [Google Scholar] [CrossRef]




| Title | Date | Patents | Country | Plants | Bacteria | Application | References |
|---|---|---|---|---|---|---|---|
| Antibacterial essential oil | 2023 | CN115708794A | China | Grape, Zedoariae rhizoma, Radix angelicae pubescentis, myrrh, Ligusticum wallichii, Eucalyptus globulus, Boswellia carterii, clove, peppermint, and coriander | Streptococcus pyogenes, S. aureus, and K. pneumoniae | It inhibits the formation of a biofilm on the surface of a biological material. | [53] |
| An antibacterial and anti-inflammatory composition containing plant extracts that provides itching relief, to be applied accordingly | 2022 | CN115844777A | China | Basil, bergamot, Salvia miltiorrhiza, witch hazel, aloe, mint, juniper berry, camellia seed, calendula, Polygonum multiflorum, honeysuckle, camphor tree, pseudo-ginseng, honeysuckle, olive, camellia, tea, daphne, Gentiana rigescens, Polygonatum kingianum, licorice, and Chrysanthemum | Escherichia coli, S. aureus, and Candida albicans | It can be used in oral care products, medicines, and skin care products, it provides relief from itching, and it has antibacterial and anti-inflammatory effects. | [54] |
| Plant composite antibacterial agent containing peony extracts, to be prepared and applied accordingly | 2022 | CN115399343B | China | Scutellaria baicalensis, aloe, selfheal, honeysuckle, Pogostemon cablin, oregano, clove, lavender, and Folium artemisiae argyi or Forsythia suspensa | S. aureus, C. albicans, E. coli, and P. Aeruginosa | It is highly sanitary and safe, and it has low metal corrosion and low skin irritation.It has fast-acting, highly efficient, and long-lasting properties. | [55] |
| Antibacterial hand sanitizer composition containing plant extracts | 2021 | KR102424044B1 | South Korea | Leek, green onion, purslane, water parsley, and perilla leaves | E. coli, S. aureus, and S. epidermidis | It has excellent moisturizing abilities to help maintain skin health. Moreover, it is possible to formulate a hand sanitizer with excellent sterilization power. | [56] |
| Plant antibacterial mite-killing agent, to be prepared accordingly, and it can be used as a daily essential | 2021 | CN112868678A | China | Thymus vulgaris, rosemary, Sophora flavescens, Folium artemisiae argyi, licorice, and dandelion | E. coli, S. aureus, and C. albicans | The active molecules of the plant extract can act on the brain nerve cells of the mites, thus stimulating the brain neurons, and enabling the mites to enter a deep sleep; this will achieve the effect of efficiently killing the mites. | [57] |
| Composition for improving antibacterial, anti-inflammatory, antiviral, and immune functions, comprising the extract of ligularia stenocephala as an active ingredient | 2020 | WO2021182661A1 | South Korea | Ligularia stenocephala | E. coli, P. aeruginosa, Aspergillus niger, Staphylococcus hominis, Bacillus subtilis, and Streptococcus pneumoniae | It may be offered as a health-functional food composition or a pharmaceutical composition that enhances antibacterial, anti-inflammatory, and immune functions. | [58] |
| Plant extract compositions and methods to make and use plant extract compositions | 2020 | US20210386074A1 | United States | Ginger, green coffee, rosemary, and honeysuckle | Klebsiella aerogenes, and S. aureus | The extract composition of the present invention may have general or broad-spectrum disinfectant efficacy. | [59] |
| Plant extract hydrolysates and an antibacterial product containing plant extract hydrolysates | 2009 | US9138451B2 | United States | Equiseti, Juglandis, Millefolii, Quercus, Taraxaci, Althaeae, Matricariae, Centaurium, Levisticum, Rosmarinus, Angelica(e), Artemisia, Astragalus, Leonurus, Salvia, Saposhnikovia, Scutellaria, Siegesbeckia, Armoracia, Capsicum, Cistus, Echinacea, Echinacea, Galphimia, and Hedera | S. aureus, S. epidermidis, S. pyogenes, S. pneumoniae, Streptococcus mutans, and Haemophilus influenzae | It can be used to produce agents with antibacterial effects against severe infections. | [60] |
| Antibacterial composition comprising plant extracts | 2002 | WO2003035093A1 | South Korea | Foeniculum vulgare, Illicium verum, Asarum heterotropoides, Cinnamomum | Candida and Trichophyton sp. | An antifungal composition that is safe for skin and has superior antifungal activity. It can be applied as a cleaner, a treating agent for dermatomycosis, such as athlete’s foot, a disinfectant, among other uses. | [61] |
| Geographical Origin | Floral Origin | Bacterial Strain | Analytical Method | References |
|---|---|---|---|---|
| Australia | E. marginata, E. patens, E. platypus, E. wandoo Banksia spp., Callistemon spp., Corymbia calophylla, Leptospermum subtenue, and Leptospermum scoparium | S. aureus ATCC 25923, ATCC 29213 and NCTC 10442, S. epidermidis ATCC 11047, E. faecalis ATCC 29212 and ATCC 51299, A. baumannii ATCC 7844, E. coli ATTC 25922, P. aeruginosa ATCC, 27853, and Salmonella enterica subsp. enterica serovar Typhimurium ATCC 13311 | Kirby–Bauer Test Minimal inhibitory concentration (MIC) and minimal bactericidal concentration (MBC) | [85] |
| E. macrorrhyncha | Alcaligenes faecalis, Citrobacter freundii, E. coli, K. aerogenes, K. pneumoniae, Mycobacterium phlei, Salmonella enterica subsp. enterica serovar California, Salmonella enterica subsp. enterica serovar Enteritidis, S. Typhimurium, Shigella sonnei, S. aureus and S. epidermidis, Serratia marcescens, and C. albicans | MIC (agar dilution method) | [86] | |
| Colombia | Polyfloral honey | E. coli ATCC 25922, S. Typhimurium ATCC 14028, and Listeria monocytogenes ATCC 19118 | Agar diffusion technique | [87] |
| Cuba | Turbina corymbosa (L.) Raf, Ipomoea triloba L., Avicennia germinans Jacq., Govania polygama (Jack) Urb, and Lysiloma latisiquum (L.) Benth | B. subtilis ATCC 6633 and S. aureus ATCC 25923 | Minimum active dilution (MAD) via the agar incorporation technique | [88] |
| Polyfloral honey | Clinical isolates: S. aureus 13, S. epidermidis 35, S. pneumoniae 9, S. pyogenes 12, S. pyogenes C-105, S. pyogenes m46, Streptococcus agalactiae 1357, Streptococcus mitis 22, Streptococcus oralis 1235, Streptococcus anginosus 2513, Streptococcus parasanguinis 2761, Streptococcus salivarius 14, Streptococcus gordonii 143, E. faecalis 212, E. faecium 17, L. monocytogenes 49, Enterobacter cloacae 19902, C. freundii 55, Salmonella enterica subsp. enterica serovar Fyris 3813, S. marcescens 28315, A. baumanii 8, K. pneumoniae 15, P. aeruginosa 24. E. coli 23, Proteus mirabilis 112, and C. albicans 18 | MAD via the agar incorporation technique | [89] | |
| Polyfloral honey | Clinical isolates: S. aureus 13, S. epidermidis 35, S. pneumoniae 9, S. pyogenes 12, S. pyogenes C-105, S. pyogenes m46, S. agalactiae 1357, S. mitis 22, S. oralis 1235, S. anginosus 2513, S. parasanguinis 2761, S. salivarius 14, S. gordonii 143, E. faecalis 212, E. faecium 17, L. monocytogenes 49, E. cloacae 19902, C. freundii 55, S. Fyris 3813, S. marcescens 28315, A. baumanii 8, K. pneumoniae 15, P. aeruginosa 24. E. coli 23, P. mirabilis 112, and C. albicans 18 | MAD via the agar incorporation technique Inhibition of biofilm formation and removal of preformed biofilm assay Transmission Electron Microscopy (TEM) for morphology analysis | [90] | |
| Ecuador | Eucalyptus spp. | S. aureus ATCC 25923, S. pyogenes ATCC 19615, P. aeruginosa ATCC 27853, E. coli ATCC 25922, and C. albicans ATTC 90028 | MAD via the agar incorporation technique | [91] |
| S. aureus CAMP and K. pneumoniae KPC 609803 | Inhibition of biofilm formation and removal of preformed biofilm assays | [80] | ||
| Eucalyptus spp. | S. aureus MRSA ATCC 2592, P. aeruginosa ATCC 2785, and S. aureus MRSA S21 (clinical isolate) | Inhibition of biofilm formation and removal of preformed biofilm assay TEM for morphology analysis | [92] | |
| Eucalyptus spp. | S. aureus, MRSA S21 (clinical isolate), and P. aeruginosa P28 clinical isolate) | Inhibition of biofilm formation and removal of preformed biofilm assay TEM for morphology analysis | [79] | |
| Persea americana | S. aureus CAMP and K. pneumoniae KPC 609803 | Inhibition of biofilm formation and removal of preformed biofilm assay | [80] | |
| Brassica napus | S. aureus CAMP and K. pneumoniae KPC 609803 | Inhibition of biofilm formation and removal of preformed biofilm assay | [80] | |
| Iran | Eucalyptus spp. | E. coli ATCC 25922, P. aeruginosa ATCC 27853, S. aureus ATCC 25923, and E. faecalis ATCC 11700 | Kirby–Bauer Test | [93] |
| Italy | Eucalyptus spp. | S. aureus subsp. aureus ATCC 9144, and P. aeruginosa ATCC 27853 | Kirby–Bauer Test | [94] |
| Kenya | Polyfloral honey | Clinical isolates: S. aureus 13, S. epidermidis 35, S. pneumoniae 9, S. pyogenes 12, S. pyogenes C-105, S. pyogenes m46, S. agalactiae 1357, S. mitis 22, S. oralis 1235, S. anginosus 2513, S. parasanguinis 2761, S. salivarius 14, S. gordonii 143, E. faecalis 212, E. faecium 17, L. monocytogenes 49, E. cloacae 19902, C. freundii 55, S. Fyris 3813, S. marcescens 28315, A. baumanii 8, K. pneumoniae 15, P. aeruginosa 24, E. coli 23, P. mirabilis 112, and C. albicans 18 | MAD via the agar incorporation technique Inhibition of biofilm formation and removal of preformed biofilm assay TEM for morphology analysis | [90] |
| Mauritius | Eucalyptus spp. | E. coli (clinical isolate), E. coli ATCC 25922, Proteus spp. (clinical isolate), P. mirabilis ATCC 12453, Pseudomonas spp. (clinical isolate), P. aeruginosa ATCC 27853, Klebsiella spp. (clinical isolate), Streptococcus spp. (clinical isolate), and S. epidermidis ATCC 35984 and ATCC 14990 | Kirby–Bauer Test | [95] |
| New Zealand | Leptospermum scoparium | Clinical isolates: S. aureus 13, S. epidermidis 35, S. pneumoniae 9, S. pyogenes 12, S. pyogenes C-105, S. pyogenes m46, S. agalactiae 1357, S. mitis 22, S. oralis 1235, S. anginosus 2513, S. parasanguinis 2761, S. salivarius 14, S. gordonii 143, E. faecalis 212, E. faecium 17, L. monocytogenes 49, E. cloacae 19902, C. freundii 55, S. Fyris 3813, S. marcescens 28315, A. baumanii 8, K. pneumoniae 15, P. aeruginosa 24. E. coli 23, P. mirabilis 112, and C. albicans 18 | MAD via the agar incorporation technique | [89] |
| Leptospermum scoparium var. Incanum, Leptospermum scoparium var. incanum + Kunzea ericoides Leptospermum scoparium var. incanum + Kunzea ericoides, and Trifolium spp. | S. aureus NCTC 8325 and ATCC 25923, S. aureus HA-MRSA, and S. aureus CA-MRSA | Inhibition of biofilm formation and removal of preformed biofilm assays | [96] | |
| Pakistan | Ziziphus mauritiana, Azadirachta indica, Ziziphus spina-christi, Citrus sinensis, and Brassica nigra | Salmonella enterica subsp. enterica serovar Typhi | MBC, MIC, and agar well diffusion assays | [97] |
| Poland | Prunus spinosa L., Polyfloral honey, Salix spp., Brassica napus L., Phacelia tanacetifolia Benth., Solidago vigaurea L., and Helianthus spp. | E. coli D31 (CGSC 5165), Bacillus circulans ATCC 61; S. aureus, 1-KI (clinical isolate), P. aeruginosa (ATCC 27853), P. aeruginosa 02/18 (clinical isolate), A. niger 71, Saccharomyces cerevisiae, and C. albicans | MAD via the agar incorporation technique | [98] |
| Slovakia | Crataegus laevigata, Abies alba Mill, and Robinia pseudoacacia | P. mirabilis and E. cloacae | MAD via the agar incorporation technique Removal of preformed biofilm assay | [99] |
| Robinia pseudoacacia, Rubus spp., Brassica napus, Rubus idaeus, and Phacelia spp. | P. aeruginosa CCM1960 and S. aureus CCM4223 | MIC and MBC assays | [100] | |
| Spain | Eucalyptus spp. | S. aureus and MRSA (clinical isolate), S. pyogenes, E. coli, and P. aeruginosa (clinical isolate) | Disk–plate diffusion method | [101] |
| Turkey | Nigella sativa L. | E. coli ATCC 25,922, E. faecalis ATCC 29,121, S. aureus ATCC 6538, S. enteric subsp. enterica ATCC 14,028/363–154, B. subtilis B209, Bacillus cereus, and L. monocytogenes ATCC 7677 | Disc diffusion and MIC assays | [102] |
| Type of Biotics | Compounds and/or Species | Antimicrobial Activity | References |
|---|---|---|---|
| Prebiotics | Punica granatum peel extract | In vitro and in vivo Caenorhabditis elegans nematode model assays demonstrated a reduction in hemolytic activity and biofilm formation caused by P. aeruginosa and they promoted the growth of B. bifidum and L. plantarum probiotic strains. | [176] |
| Chitin-glucan (CG) | In vitro assays showed high levels of growth in all bifidobacterial species, particularly the Bifidobacterium breve 2L isolate in the in vivo Groningen rat model, which became more abundant in the gut of B. breve 2L. | [177] | |
| Phthalyl pullulan nanoparticles (PPNs) | A gut dysbiosis-induced murine model was used, and their restorative effect in the eubiosis microbiota was assessed using the pathogen, E. coli K99. | [167] | |
| Ursodeoxycholic acid (UDCA) | In vitro assays demonstrated a reduction in E. coli serotype O101:H9 growth, proinflammatory effects in Caco-2 cells, and cell integrity damage. Moreover, in vivo assays used on neonatal mice model also exhibited attenuated colitis symptoms and recovered colonic short-chain fatty acid (SCFA) production. | [178] | |
| Probiotics | Two Lactococcus lactis subsp. lactis strains | In vivo bacterial feeding of these probiotic strains for 30 days in a Swiss albino mice model was conducted, and they improved gut colonization and IgA levels. | [168] |
| Honey probiotic B. circulans isolate | In vitro co-culture of B. circulans and Cutibacterium acnes significantly suppressed pathogen growth. Moreover, in vivo assays using the ICR mice model, B. circulans, generated electrons that inhibited C. acnes growth and diminished inflammation. | [179] | |
| Bacillus amyloliquefaciens (BA PMC-80) | In vitro co-culture assays of BA PMC-80 and Clostridioides difficile demonstrated significant pathogen inhibition; an in vivo hamster model exhibited no toxicity, a less severe infection, and late death. | [180] | |
| Two new strains of B. subtilis (CH311 and S3B) | In vitro gut model demonstrated the ability of B. subtilis CH311 and S3B to reduce ESBL-E. coli titers using 4 log CFU/mL; however, the in vivo murine model showed no reduction in the ESBL-E. coli fecal titers. | [181] | |
| Synbiotics | Pediococcus acidilactici plus phthalyl inulin nanoparticles (PINs) | In vitro antimicrobial activity was tested using a cocultivation assay. A statistical reduction of more than 3 log CFU/mL of Salmonella enterica subsp. enterica serovar Gallinarum, together with P. acidilactici plus PINs, was observed when compared with the control and the PINs or probiotic groups alone. | [182] |
| Encapsulated S. cerevisiae plus Moringa oleifera leaf extract (MOLE) | In vivo rabbit model revealed no effects on interleukin-l or IgG and IgA levels, and it showed a significantly higher number of beneficial microbes. Moreover, a significant increase in in vitro inhibitory activities was observed against E. coli BA 12296B, S. aureus NCTC 10788, C. albicans ATCC MYA-2876, L. monocytogenes ATCC 19116, and Salmonella enterica subsp. enterica serovar Senftenberg ATCC 8400. | [183] | |
| PPNs plus L. plantarum | A gut dysbiosis-induced murine model was used and the E. coli K99 infection was markedly suppressed after several well-known beneficial bacteria, including Lactobacillus and Bifidobacterium, were incrementally introduced. | [167] | |
| Two L. lactis subsp. lactis strains with inulin | The Swiss albino mice model exhibited a significant reduction in IgA levels that is comparable with commercial probiotics and prebiotic consortiums on the market. | [168] | |
| Postbiotics | Lyophilized cell-free supernatants (LCFS) of Lactobacillus isolates | Demonstrated strong inhibition and eradication antibiofilm activities for A. baumannii and E. coli, and a reduction in nitric oxide production in the RAW 264.7 cell line was observed. | [164] |
| L. plantarum KM1, L. plantarum KM2, Bacillus velezensis KMU01 postbiotics mixtures 1:1:1 (vol/vol) | NK cell activation was significantly higher in the C57BL/6N mice model, and TNF-α levels in the RAW264.7 cell line was significantly reduced when compared with the LPS. | [184] | |
| Individual LCFS of L. fermentum, L. reuteri, and B. subtilis sp. natto in a postbiotic cold cream | All postbiotic cold creams exhibited different degrees of immunomodulatory, anti-inflammatory, and antimicrobial activities in the Sprague Dawley rat model when compared with controls (no treatment and only cold cream). | [185] | |
| Indole-3-carboxaldehyde (3-IAld) (a microbial tryptophan metabolite) | The use of 3-IAld inhalable dry powder demonstrated optimal pulmonary administration and toxicological safety, also reducing aspergillosis scores by acting on the infection and inflammation sites. | [186] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado, A.; Zamora-Mendoza, L.; Alexis, F.; Álvarez-Suarez, J.M. Use of Plant Extracts, Bee-Derived Products, and Probiotic-Related Applications to Fight Multidrug-Resistant Pathogens in the Post-Antibiotic Era. Future Pharmacol. 2023, 3, 535-567. https://doi.org/10.3390/futurepharmacol3030034
Machado A, Zamora-Mendoza L, Alexis F, Álvarez-Suarez JM. Use of Plant Extracts, Bee-Derived Products, and Probiotic-Related Applications to Fight Multidrug-Resistant Pathogens in the Post-Antibiotic Era. Future Pharmacology. 2023; 3(3):535-567. https://doi.org/10.3390/futurepharmacol3030034
Chicago/Turabian StyleMachado, António, Lizbeth Zamora-Mendoza, Frank Alexis, and José Miguel Álvarez-Suarez. 2023. "Use of Plant Extracts, Bee-Derived Products, and Probiotic-Related Applications to Fight Multidrug-Resistant Pathogens in the Post-Antibiotic Era" Future Pharmacology 3, no. 3: 535-567. https://doi.org/10.3390/futurepharmacol3030034