
Exploring the Pharmacological Potential of Glycyrrhizic Acid: From Therapeutic Applications to Trends in Nanomedicine


Abstract
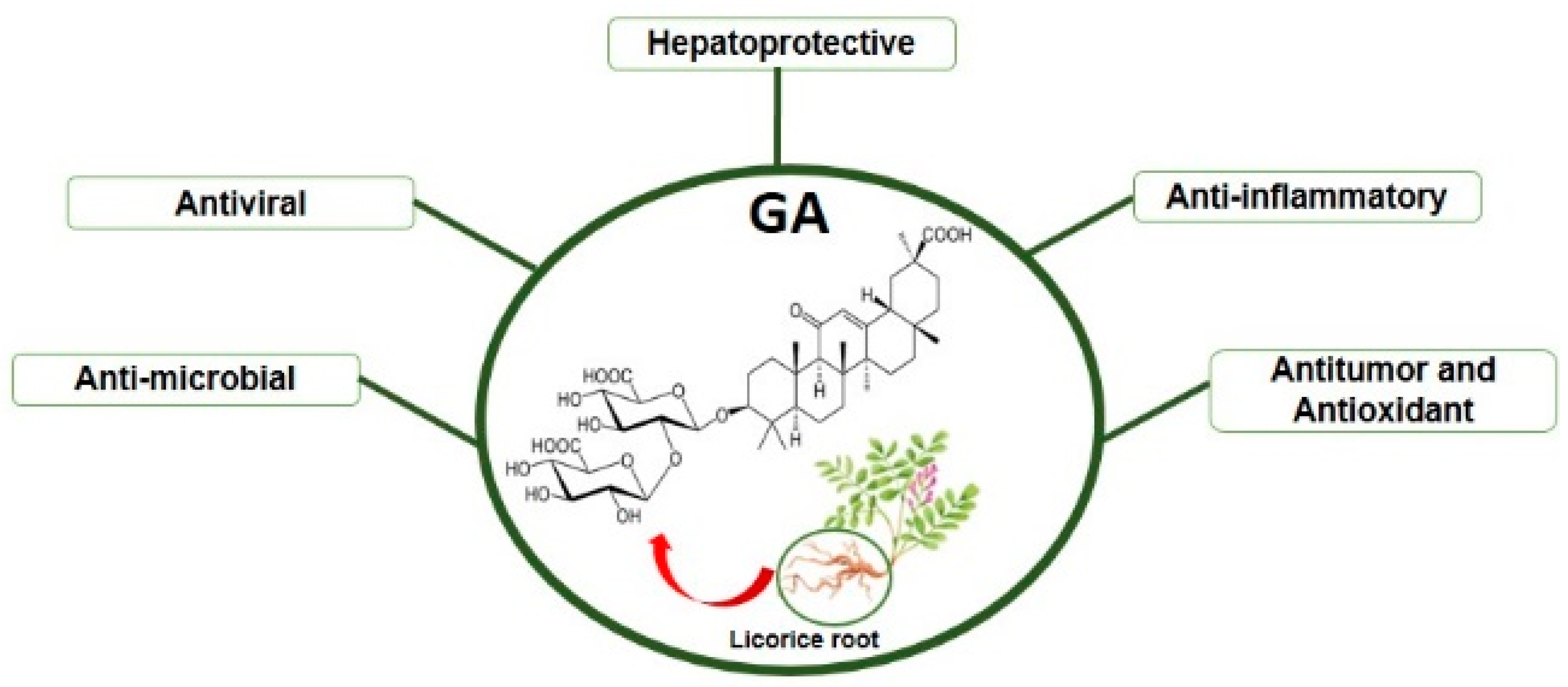
:1. Introduction
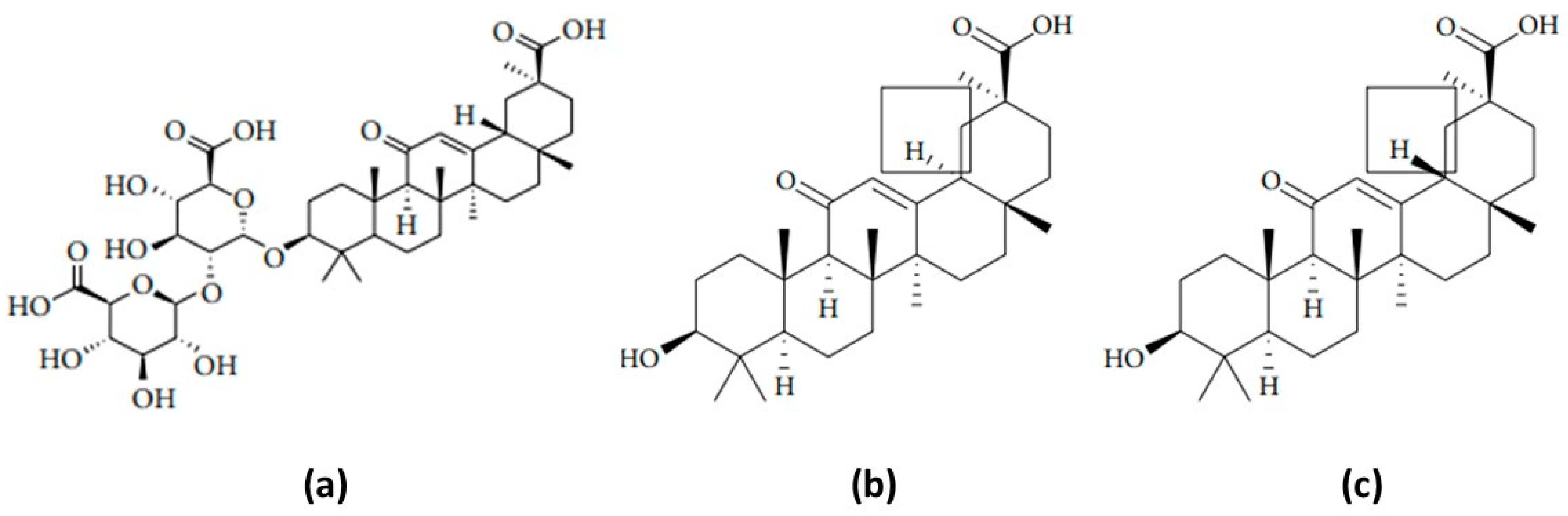
2. GA: Molecular Mechanisms of Action and Pharmacological Applications
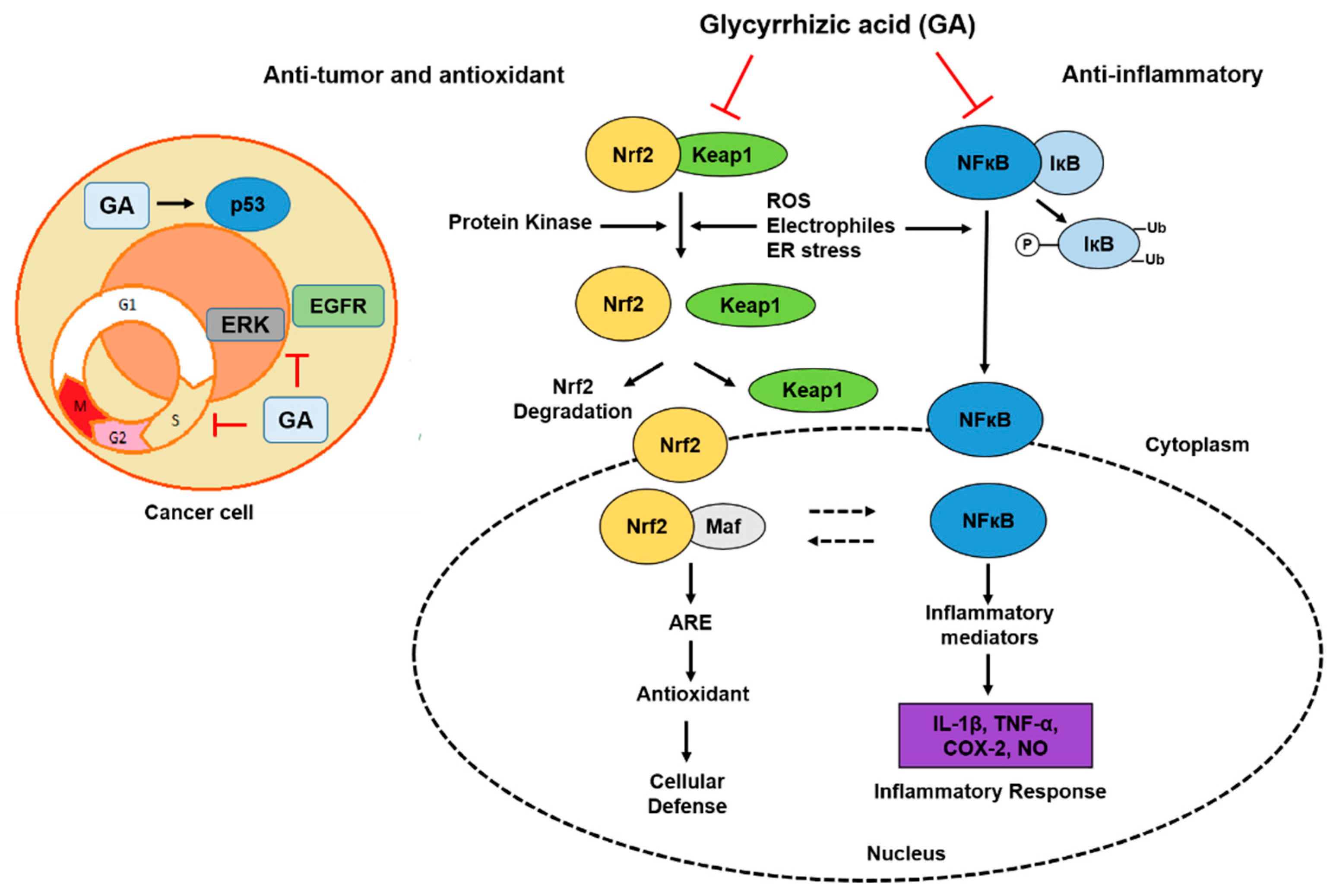
2.1. Anti-Inflammatory and Hepatoprotective Activity
2.2. Antiviral and Anti-Parasitic Activity
2.3. Antibacterial Activity
2.4. Antitumor and Antioxidant Activities
2.5. Some Evidence for GA Efficacy: Clinical Trials

{kind=link}
{kind=link}
{kind=link}
| Pharmacological Activities | Main Results | References |
|---|---|---|
| Anti-inflammatory | GA exhibits antiinflammatory effects through inhibition of MIP-1 in a mouse model of acute P. acnes-induced inflammatory liver injury. | [18] |
| Anti-inflammatory | GA interrupted JNK/c-Jun and IκB/NF-κB signaling pathways, which decrease activator protein-1 (AP-1) and NF-κB mediated ICAM-1 expressions. | [19] |
| Anti-inflammatory | GA inhibits the expression of TNF-α, IL-6, IL-1β and RANTES in LPS-stimulated macrophages. | [20] |
| Anti-inflammatory | GA inhibits IL-1β induced inflammation by blocking PI3K/Akt phosphorylation and NF-κB activation and alleviated OA progression in surgical-induced DMM mouse model. | [22] |
| Anti-inflammatory | GA inhibits the LPS/D-galactosamine-induced liver injury through preventing inflammatory responses and IL-18 production. | [23] |
| Anti-inflammatory | GA may prevent tissue injury in chronic hepatitis and in many autoimmune diseases by suppressing the lytic pathway of the complement system. | [24] |
| Anti-inflammatory | GA was effective and reduced atopic dermatitis in a human and children double-blind clinical trial. | [56,57,58] |
| Anti-inflammatory | GA promoted ulcer healing in patiets with peptic ulcer. | [6,63,64] |
| Antiviral | GA augmented IFN-induced reduction of virus in the HCVcc system (cell culture produced HCV). | [15] |
| Antioxidant | GA were able to increase the intracellular reduced glutathione concentration, in AFB1-treated cells. | [25] |
| Anti-apoptotic | GA has inhibitory effects on hepatocyte apoptosis and liver fibrosis. | [26] |
| Antiviral | GA increased the number of OKT4 lymphocytes, and demonstrated a suitable treatment for preventing the development of asymptomatic carrier (AC) in hemophilia patients into AIDS. | [27] |
| Antiviral | GA attenuated inflammatory responses in HSV by inhibition of adhesion between CCEC and PMN. | [28] |
| Antiviral | Water Extract of GA has inhibitory Inhibited Enterovirus 71 in a Human Foreskin Fibroblast Cell Line. | [29] |
| Antiviral | Inhibitory effect of GA on HIV replication in patients with AIDS. | [31] |
| Antiviral | GA has inhibitory effect on a chemoattractant (RANTES) released by influenza A virus (H1N1)-infected human bronchial epithelial cells. | [32] |
| Antiviral | GA inhibits replication, adsorption, and penetration of the virus during the early steps of the replica-tive cycle in Vero cells. GA inhibits replication of the SARS-associated virus. | [33] |
| Antiviral | GA inhibits replication of the SARS-associated virus. | [34] |
| Anti-parasitic | GA suppressed inflammation in Leishmania donovani infection by inhibiting COX-2-mediated PGE2 release. | [38] |
| Anti-parasitic | GA adecreased hepatic and splenic parasite burden and increased T cell proliferation in Leishmania-infected BALB/c mice. | [39] |
| Anti-parasitic | GA has anthelmintic activity against gastrointestinal nematodes of small ruminants. | [40] |
| Anti-parasitic | GA has a potential antileishmanial chemotherapeutic agent by killing the parasite affecting sterol biosynthetic pathway. | [41] |
| Antibacterial | GA inhibited Arylamine N-acetyltransferase (NAT) activities in a strain of H. pylori. | [42] |
| Antibacterial | GA promoted antibacterial resistance of severely burned mice to P. aeruginosa burn wound infection. | [43] |
| Antibacterial | GA reduced skin lesion size and attenuates expression of key virulence genes in a mouse model of S. aureus. | [44] |
| Antibacterial | GA showed a significant antibacterial activity against S. aureus, E. coli, P. fluorescens, and Bacillus cereus. | [45] |
| Antibacterial | GA increased the eradication rate of H. pylori in patients with gastrointestinal disorders. | [60,61,62] |
| Antioxidant | GA were able to increase the intracellular reduced glutathione concentration, in AFB1-treated cells. | [25] |
| Anti-apoptotic | GA has inhibitory effects on hepatocyte apoptosis and liver fibrosis. | [26] |
| Antitumor | GA exhibited anti-tumor property in astric ancer cells partly by inducing apoptosis and cell cycle arrest. | [47] |
| Antitumor and Antioxidat | GA induces apoptotic cell death in SiHa cells and exhibits a synergistic effect against antibiotic, anti-cancer and drug toxicity. | [48] |
| Antitumor | GA has chemopreventive effect via modulation of inflammatory markers and induction of apoptosis in human hepatoma cell line (HepG2). | [50] |
| Antibacterial | GA suppresses the development of precancerous lesions via regulating the hyperproliferation, inflammation, angiogenesis and apoptosis in the colon of wistar rats. | [52] |
| Antitumor and Antioxidat | GA induced apoptosis and was found to modulate critical end points of oxidative stress in c.ultured primary rat hepatocytes. | [54] |
| Antitumor and Antioxidat | GA induces programmed cell death, probably inhibiting the liver enzyme 11-hydroxysteroid dehydrogenase type. | [55] |
| Antitumor and Antiviral | GA reduced the increased risk of hepatocellular carcinoma in patients with HCC. | [6,59] |
2.6. Glycyrrhizic Acid and Their Therapeutic Associations—The Role of Nanomedicine
3. Final Considerations
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tomazzoni, M.I.; Negrelle, R.R.B.; Centa, M.D.L. Fitoterapia Popular: A Busca Instrumental Enquanto Prática Terapêuta. Texto Context. -Enferm. 2006, 15, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Cao, H.Y.; Liu, P.; Cheng, G.H.; Sun, M.Y. Glycyrrhizic Acid in the Treatment of Liver Diseases: Literature Review. Biomed Res. Int. 2014, 872139. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, G.; Curcio, M.; Parisi, O.I.; Puoci, F.; Iemma, F.; Spizzirri, U.G.; Restuccia, D.; Picci, N. Molecularly Imprinted Polymers for the Selective Extraction of Glycyrrhizic Acid from Liquorice Roots. Food Chem. 2011, 125, 1058–1063. [Google Scholar] [CrossRef]
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues, F.; Oliveira, M.B.P.P. Liquorice (Glycyrrhiza glabra): A Phytochemical and Pharmacological Review. Phyther. Res. 2018, 32, 2323–2339. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Chang, Z.; Liu, J.; Sun, X.; Hu, X.; Liu, H. Separation of Glycyrrhizic Acid and Liquiritin from Glycyrrhiza uralensis Fisch Extract by Three-Liquid-Phase Extraction Systems. Sep. Purif. Technol. 2007, 53, 216–223. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Son, D.H.; Chung, T.H.; Lee, Y.J. A Review of the Pharmacological Efficacy and Safety of Licorice Root from Corroborative Clinical Trial Findings. J. Med. Food 2020, 23, 12–20. [Google Scholar] [CrossRef]
- Obolentseva, G.V.; Litvinenko, V.I.; Ammosov, A.S.; Popova, T.P.; Sampiev, A.M. Pharmacological and Therapeutic Properties of Licorice Preparations (A Review). Pharm. Chem. J. 1999, 33, 427–434. [Google Scholar] [CrossRef]
- Ploeger, B.; Mensinga, T.; Sips, A.; Seinen, W.; Meulenbelt, J.; DeJongh, J. The Pharmacokinetics of Glycyrrhizic Acid Evaluated by Physiologically Based Pharmacokinetic Modeling. Drug Metab. Rev. 2001, 33, 125–147. [Google Scholar] [CrossRef]
- Ming, L.J.; Yin, A.C.Y. Therapeutic Effects of Glycyrrhizic Acid. Nat. Prod. Commun. 2013, 8, 415–418. [Google Scholar] [CrossRef] [Green Version]
- Selyutina, O.Y.; Polyakov, N.E. Glycyrrhizic acid as a multifunctional drug carrier—From physicochemicalproperties to biomedical applications: A modern insight on the ancient drug. Int. J. Pharm. 2019, 559, 271–279. [Google Scholar] [CrossRef]
- Pan, X.; Liu, H.; Jia, G.; Shu, Y.Y. Microwave-Assisted Extraction of Glycyrrhizic Acid from Licorice Root. Biochem. Eng. J. 2000, 5, 173–177. [Google Scholar] [CrossRef]
- Niu, G.G.; Xie, Y.C.; Lou, J.F.; Liu, H.Z. Isolation and Purification of Glycyrrhizic Acid with Solvent Extraction. Sep. Purif. Technol. 2005, 44, 189–196. [Google Scholar] [CrossRef]
- Tian, M.; Yan, H.; Row, K.H. Extraction of Glycyrrhizic Acid and Glabridin from Licorice. Int. J. Mol. Sci. 2008, 9, 571–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takei, M.; Kobayashi, M.; Herndon, D.N.; Pollard, R.B.; Suzuki, F. Glycyrrhizin Inhibits the Manifestations of Anti-Inflammatory Responses That Appear in Association with Systemic Inflammatory Response Syndrome (SIRS)-like Reactions. Cytokine 2006, 35, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Matsuura, T.; Aoyagi, H.; Matsuda, M.; Hmwe, S.S.; Date, T.; Watanabe, N.; Watashi, K.; Suzuki, R.; Ichinose, S.; et al. Antiviral Activity of Glycyrrhizin against Hepatitis C Virus in vitro. PLoS ONE 2013, 8, e68992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.Y.; Ha, J.Y.; Kim, K.M.; Jung, Y.S.; Jung, J.C.; Oh, S. Anti-Inflammatory Activities of Licorice Extract and Its Active Compounds, Glycyrrhizic Acid, Liquiritin and Liquiritigenin, in BV2 Cells and Mice Liver. Molecules 2015, 20, 13041–13054. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Gong, X.; Zhang, L.; Jiang, R.; Kuang, G.; Wang, B.; Chen, X.; Wan, J. Glycyrrhetinic Acid Attenuates Lipopolysaccharide-Induced Fulminant Hepatic Failure in D-Galactosamine-Sensitized Mice by up-Regulating Expression of Interleukin-1 Receptor-Associated Kinase-M. Toxicol. Appl. Pharmacol. 2017, 320, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xu, J.; Mao, C.; Jin, M.; Wu, Q.; Zou, J.; Gu, Q.; Zhang, Y.; Zhang, Y. 18β-Glycyrrhetinic Acid Ameliorates Acute Propionibacterium Acnes-Induced Liver Injury through Inhibition of Macrophage Inflammatory Protein-1α. J. Biol. Chem. 2010, 285, 1128–1137. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.L.; Chen, C.L.; Kuo, C.L.; Chen, B.C.; You, J.S. Glycyrrhetinic Acid Inhibits ICAM-1 Expression via Blocking JNK and NF-B Pathways in TNF-α-Activated Endothelial Cells. Acta Pharmacol. Sin. 2010, 31, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Zhou, E.; Wei, Z.; Song, X.; Liu, Z.; Wang, T.; Wang, W.; Zhang, N.; Liu, G.; Yang, Z. Glycyrrhizin Inhibits Lipopolysaccharide-Induced Inflammatory Response by Reducing TLR4 Recruitment into Lipid Rafts in RAW264.7 Cells. Biochim. Biophys. Acta-Gen. Subj. 2014, 1840, 1755–1764. [Google Scholar] [CrossRef]
- Wang, X.; Gu, X.; Wang, H.; Sun, Y.; Wu, H.; Mao, S. Synthesis, Characterization and Liver Targeting Evaluation of Self-Assembled Hyaluronic Acid Nanoparticles Functionalized with Glycyrrhetinic Acid. Eur. J. Pharm. Sci. 2017, 96, 255–262. [Google Scholar] [CrossRef]
- Jiang, R.H.; Xu, J.J.; Zhu, D.C.; Li, J.F.; Zhang, C.X.; Lin, N.; Gao, W.Y. Glycyrrhizin Inhibits Osteoarthritis Development through Suppressing the PI3K/AKT/NF-ΚB Signaling Pathway: In Vivo and in Vitro. Food Funct. 2020, 11, 2126–2136. [Google Scholar] [CrossRef]
- Yoshida, T.; Abe, K.; Ikeda, T.; Matsushita, T.; Wake, K.; Sato, T.; Sato, T.; Inoue, H. Inhibitory Effect of Glycyrrhizin on Lipopolysaccharide and D-Galactosamine-Induced Mouse Liver Injury. Eur. J. Pharmacol. 2007, 576, 136–142. [Google Scholar] [CrossRef]
- Fujisawa, Y.; Sakamoto, M.; Matsushita, M.; Fujita, T.; Nishioka, K. Glycyrrhizin Inhibits the Lytic Pathway of Complement—Possible Mechanism of Its Anti-Inflammatory Effect on Liver Cells in Viral Hepatitis. Microbiol. Immunol. 2000, 44, 799–804. [Google Scholar] [CrossRef]
- Chan, H.T.; Chan, C.; Ho, J.W. Inhibition of Glycyrrhizic Acid on Aflatoxin B1-Induced Cytotoxicity in Hepatoma Cells. Toxicology 2003, 188, 211–217. [Google Scholar] [CrossRef]
- Liang, B.; Guo, X.L.; Jin, J.; Ma, Y.C.; Feng, Z.Q. Glycyrrhizic Acid Inhibits Apoptosis and Fibrosis in Carbontetrachloride-Induced Rat Liver Injury. World J. Gastroenterol. 2015, 21, 5271–5280. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Sakai, H.; Suzuki, S.; Akutsu, Y.; Ishikawa, M.; Imaizumi, M.; Tada, K.; Aihara, M.; Sawada, Y.; Yokoyama, M.; et al. Effects of Glycyrrhizin (SNMC: Stronger Neo-Minophagen G®) in Hemophilia Patients with HIV-1 Infection. Tohoku J. Exp. Med. 1990, 162, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Chen, X.; Li, Q.; Li, P.; Zhao, G.; Xu, M.; Xie, P. Inhibition of Intercellular Adhesion in Herpex Simplex Virus Infection by Glycyrrhizin. Cell Biochem. Biophys. 2012, 62, 137–140. [Google Scholar] [CrossRef]
- Kuo, K.K.; Chang, J.S.; Wang, K.C.; Chiang, L.C. Water Extract of Glycyrrhiza Uralensis Inhibited Enterovirus 71 in a Human Foreskin Fibroblast Cell Line. Am. J. Chin. Med. 2009, 37, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xiao, Y.; Xu, L.; Liu, Y.; Jiang, G.; Wang, W.; Li, B.; Zhu, T.; Tan, Q.; Tang, L.; et al. Glycyrrhizic Acid Nanoparticles as Antiviral and Anti-Inflammatory Agents for COVID-19 Treatment. ACS Appl. Mater. Interfaces 2021, 13, 20995–21006. [Google Scholar] [CrossRef] [PubMed]
- Hattori, T.; Ikematsu, S.; Koito, A.; Matsushita, S.; Maeda, Y.; Hada, M.; Fujimaki, M.; Takatsuki, K. Preliminary Evidence for Inhibitory Effect of Glycyrrhizin on HIV Replication in Patients with AIDS. Antivir. Res. 1989, 11, 255–261. [Google Scholar] [CrossRef]
- Ko, H.C.; Wei, B.L.; Chiou, W.F. The Effect of Medicinal Plants Used in Chinese Folk Medicine on RANTES Secretion by Virus-Infected Human Epithelial Cells. J. Ethnopharmacol. 2006, 107, 205–210. [Google Scholar] [CrossRef]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H.W. Glycyrrhizin, an Active Component of Liquorice Roots, and Replication of SARS-Associated Coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef] [Green Version]
- Hoever, G.; Baltina, L.; Michaelis, M.; Kondratenko, R.; Baltina, L.; Tolstikov, G.A.; Doerr, H.W.; Cinatl, J. Antiviral Activity of Glycyrrhizic Acid Derivatives against SARS-Coronavirus. J. Med. Chem. 2005, 48, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; Vergoten, G. Glycyrrhizin: An alternative drug for the treatment of COVID-19 infection and the associated respiratory syndrome? Pharmacol. Ther. 2020, 214, 107618. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Q.; Guo, D. Emerging Coronaviruses: Genome Structure, Replication, and Pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Gan, W.; Nie, Y.; Ying, R.; Tan, Y.; Chen, J.; Chen, M.; Zhang, C. Clinical Efficacy and Security of Glycyrrhizic Acid Preparation in the Treatment of Anti-SARS-CoV-2 Drug-Induced Liver Injury: A Protocol of Systematic Review and Meta-Analysis. BMJ Open 2021, 11, 1–6. [Google Scholar] [CrossRef]
- Ukil, A.; Kar, S.; Srivastav, S.; Ghosh, K.; Das, P.K. Curative Effect of 18β-Glycyrrhetinic Acid in Experimental Visceral Leishmaniasis Depends on Phosphatase-Dependent Modulation of Cellular MAP Kinases. PLoS ONE 2011, 6, e29062. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S.; Bhattacharjee, A.; Majumder, S.; Majumdar, S.B.; Majumdar, S. Glycyrrhizic Acid Suppresses Cox-2-Mediated Anti-Inflammatory Responses during Leishmania Donovani Infection. J. Antimicrob. Chemother. 2012, 67, 1905–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maestrini, M.; Molento, M.B.; Forzan, M.; Perrucci, S. In Vitro Anthelmintic Activity of an Aqueous Extract of Glycyrrhiza glabra and of Glycyrrhetinic Acid against Gastrointestinal Nematodes of Small Ruminants. Parasite 2021, 28, 64. [Google Scholar] [CrossRef]
- Dinesh, N.; Neelagiri, S.; Kumar, V.; Singh, S. Glycyrrhizic Acid Attenuates Growth of Leishmania Donovani by Depleting Ergosterol Levels Neeradi. Exp. Parasitol. 2017, 176, 21–29. [Google Scholar] [CrossRef]
- Chung, J.G.; Wu, L.T.; Chang, S.H.; Lo, H.H.; Hsieh, S.E.; Li, Y.C.; Hung, C.F. Inhibitory Actions of Berberine on Growth and Arylamine N- Acetyltransferase Activity in Strains of Helicobacter pylori from Peptic Ulcer Patients. Int. J. Toxicol. 1998, 18, 35–40. [Google Scholar] [CrossRef]
- Yoshida, T.; Yoshida, S.; Kobayashi, M.; Herndon, D.N.; Suzuki, F. Pivotal Advance: Glycyrrhizin Restores the Impaired Production of β-Defensins in Tissues Surrounding the Burn Area and Improves the Resistance of Burn Mice to Pseudomonas Aeruginosa Wound Infection. J. Leukoc. Biol. 2010, 87, 35–41. [Google Scholar] [CrossRef]
- Long, D.R.; Mead, J.; Hendricks, J.M.; Hardy, M.E.; Voyich, J.M. 18β-Glycyrrhetinic Acid Inhibits Methicillin-Resistant Staphylococcus aureus Survival and Attenuates Virulence Gene Expression. Antimicrob. Agents Chemother. 2013, 57, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Rodino, S.; Butu, A.; Butu, M.; Cornea, P.C. Comparative Studies on Antibacterial Activity of Licorice, Elderberry and Dandelion. Dig. J. Nanomater. Biostructures 2015, 10, 947–955. [Google Scholar]
- Cherng, J.M.; Tsai, K.D.; Yu, Y.W.; Lin, J.C. Molecular Mechanisms Underlying Chemopreventive Activities of Glycyrrhizic Acid against UVB-Radiation-Induced Carcinogenesis in SKH-1 Hairless Mouse Epidermis. Radiat. Res. 2011, 176, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ge, X.; Qu, H.; Wang, N.; Zhou, J.; Xu, W.; Xie, J.; Zhou, Y.; Shi, L.; Qin, Z.; et al. Glycyrrhizic Acid Inhibits Proliferation of Gastric Cancer Cells by Inducing Cell Cycle Arrest and Apoptosis. Cancer Manag. Res. 2020, 12, 2853–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.S.; Kim, Y.J.; Lee, M.S.; Han, E.S.; Lee, S.J. 18β-Glycyrrhetinic Acid Induces Apoptotic Cell Death in SiHa Cells and Exhibits a Synergistic Effect against Antibiotic Anti-Cancer Drug Toxicity. Life Sci. 2008, 83, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.Y.; Hao, M.; Yang, X.Y.; Ba, Q.; Li, M.; Ni, S.J.; Wang, L.S.; Du, X. Licochalcone A Inhibits Growth of Gastric Cancer Cells by Arresting Cell Cycle Progression and Inducing Apoptosis. Cancer Lett. 2011, 302, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.K.; Siddiqi, A.; Nafees, S.; Ali, N.; Rashid, S.; Ali, R.; Shahid, A.; Sultana, S. Chemopreventive Effect of 18β-Glycyrrhetinic Acid via Modulation of Inflammatory Markers and Induction of Apoptosis in Human Hepatoma Cell Line (HepG2). Mol. Cell. Biochem. 2016, 416, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.K.; Ara, I.; Mondal, M.S.A.; Kabir, Y. Phytochemistry, Pharmacological Activity, and Potential Health Benefits of Glycyrrhiza Glabra. Heliyon 2021, 7, e07240. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Khan, A.Q.; Lateef, A.; Rehman, M.U.; Tahir, M.; Ali, F.; Hamiza, O.O.; Sultana, S. Glycyrrhizic Acid Suppresses the Development of Precancerous Lesions via Regulating the Hyperproliferation, Inflammation, Angiogenesis and Apoptosis in the Colon of Wistar Rats. PLoS ONE 2013, 8, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konovalova, G.G.; Tikhaze, A.K.; Lankin, V.Z. Antioxidant Activity of Parapharmaceutics Containing Natural Inhibitors of Free Radical Processes. Bull. Exp. Biol. Med. 2000, 130, 658–660. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.; Singh, B.K.; Kakkar, P. Glycyrrhizic Acid Modulates T-BHP Induced Apoptosis in Primary Rat Hepatocytes. Food Chem. Toxicol. 2009, 47, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Horigome, H.; Homma, M.; Hirano, T.; Oka, K. Glycyrrhetinic Acid Induced Apoptosis in Murine Splenocytes. Biol. Pharm. Bull. 2001, 24, 54–58. [Google Scholar] [CrossRef] [Green Version]
- Kalinowska-Lis, A.K. 18 b-Glycyrrhetinic Acid: Its Core Biological Properties and Dermatological Applications. Int. J. Cosmet. Sci. 2019, 41, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Saeedi, M.; Semnani, K.; Ghoreishi, M. The Treatment of Atopic Dermatitis with Licorice Gel. J. Dermatolog. Treat. 2003, 14, 153–157. [Google Scholar] [CrossRef]
- Boguniewicz, M.; Zeichner, J.A.; Eichenfield, L.F.; Hebert, A.A.; Jarratt, M.; Lucky, A.W.; Paller, A.S. MAS063DP Is Effective Monotherapy for Mild to Moderate Atopic Dermatitis in Infants and Children: A Multicenter, Randomized, Vehicle-Controlled Study. J. Pediatr. 2008, 152, 854–859. [Google Scholar] [CrossRef]
- Ikeda, K.; Arase, Y.; Kobayashi, M.; Saitoh, S.; Someya, T.; Hosaka, T.; Sezaki, H.; Akuta, N.; Suzuki, Y.; Suzuki, F.; et al. A Long-Term Glycyrrhizin Injection Therapy Reduces Hepatocellular Carcinogenesis Rate in Patients with Interferon-Resistant Active Chronic Hepatitis C: A Cohort Study of 1249 Patients. Dig. Dis. Sci. 2006, 51, 603–609. [Google Scholar] [CrossRef]
- Hajiaghamohammadi, A.A.; Zargar, A.; Oveisi, S.; Samimi, R.; Reisian, S. To Evaluate of the Effect of Adding Licorice to the Standard Treatment Regimen of Helicobacter Pylori. Braz. J. Infect. Dis. 2016, 20, 534–538. [Google Scholar] [CrossRef] [Green Version]
- Wittschier, N.; Faller, G.; Hensel, A. Aqueous Extracts and Polysaccharides from Liquorice Roots (Glycyrrhiza glabra L.) Inhibit Adhesion of Helicobacter Pylori to Human Gastric Mucosa. J. Ethnopharmacol. 2009, 125, 218–223. [Google Scholar] [CrossRef]
- An, B.; Moon, B.S.; Kim, H.; Lim, H.C.; Lee, Y.C.; Lee, G.; Kim, S.-H.; Park, M.; Kim, J.B. Antibiotic Resistance in Helicobacter Pylori Strains and Its Effect on H. Pylori Eradication Rates in a Single Center in Korea. Ann. Lab. Med. 2013, 33, 415–419. [Google Scholar] [CrossRef] [Green Version]
- Turpie, A.G.G.; Thomson, T.J. Carbenoxolone Sodium in the Treatment of Gastric Ulcer with Special Reference to Side-Effects. Gut 1965, 5, 591–594. [Google Scholar] [CrossRef] [Green Version]
- Pinder, R.M.; Brogden, R.N.; Sawyer, P.R.; Speight, T.M.; Spencer, R.; Avery, G.S. Evaluations on New Drugs. Drugs 1976, 11, 245–307. [Google Scholar] [CrossRef]
- Ochi, M.M.; Amoabediny, G.; Rezayat, S.M.; Akbarzadeh, A.; Ebrahimi, B. In Vitro Co-Delivery Evaluation of Novel Pegylated Nano-Liposomal Herbal Drugs of Silibinin and Glycyrrhizic Acid (Nano-Phytosome) to Hepatocellular Carcinoma Cells. Cell J. 2016, 18, 135–148. [Google Scholar] [CrossRef]
- Kurosaki, T.; Kawanabe, S.; Kodama, Y.; Fumoto, S.; Nishida, K.; Nakagawa, H.; Higuchi, N.; Nakamura, T.; Kitahara, T.; Sasaki, H. Hepatic Gene Delivery System Electrostatically Assembled with Glycyrrhizin. Mol. Pharm. 2014, 11, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Wu, L.; Hu, M.; Dong, W.; Xu, M.; Zhang, P. Glycyrrhizic Acid: A Promising Carrier Material for Anticancer Therapy. Biomed. Pharmacother. 2017, 95, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, K.; Miyajima, R.; Ishida, Y.; Karasawa, S.; Yoshimura, T. Aggregate Formation of Glycyrrhizic Acid. Colloids Surf. A Physicochem. Eng. Asp. 2016, 500, 112–117. [Google Scholar] [CrossRef]
- Borisenko, S.N.; Lekar’, A.V.; Vetrova, E.V.; Filonova, O.V.; Borisenko, N.I. A Mass Spectrometry Study of the Self-Association of Glycyrrhetinic Acid Molecules. Russ. J. Bioorganic Chem. 2016, 42, 716–720. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, Z.; Zheng, Y.; Geng, Y.; Han, C.; Shi, Y.; Sun, H.; Zhang, C.; Chen, Y.; Zhang, L.; et al. Glycyrrhetinic Acid Functionalized Graphene Oxide for Mitochondria Targeting and Cancer Treatment In Vivo. Small 2018, 14, 1–16. [Google Scholar] [CrossRef]
- Wang, W.; Luo, M.; Fu, Y.; Wang, S.; Efferth, T.; Zu, Y. Glycyrrhizic Acid Nanoparticles Inhibit LPS-Induced Inflammatory Mediators in 264.7 Mouse Macrophages Compared with Unprocessed Glycyrrhizic Acid. Int. J. Nanomed. 2013, 8, 1377–1383. [Google Scholar] [CrossRef] [Green Version]
- Zu, Y.; Meng, L.; Zhao, X.; Ge, Y.; Yu, X.; Zhang, Y.; Deng, Y. Preparation of 10-Hydroxycamptothecin-Loaded Glycyrrhizic Acid-Conjugated Bovine Serum Albumin Nanoparticles for Hepatocellular Carcinoma-Targeted Drug Delivery. Int. J. Nanomed. 2013, 8, 1207–1222. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.H.; Zhang, Q.; Liang, Q.Y.; Wang, S.Q.; Zhao, B.X.; Wang, Y.T.; Cai, Y.; Li, G.F. Bioavailability Enhancement of Paclitaxel via a Novel Oral Drug Delivery System: Paclitaxel-Loaded Glycyrrhizic Acid Micelles. Molecules 2015, 20, 4337–4356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeeshan, M.; Ali, H.; Khan, S.; Mukhtar, M.; Khan, M.I.; Arshad, M. Glycyrrhizic Acid-Loaded PH-Sensitive Poly-(Lactic-Co-Glycolic Acid) Nanoparticles for the Amelioration of Inflammatory Bowel Disease. Nanomedicine 2019, 14, 1945–1969. [Google Scholar] [CrossRef] [PubMed]
- Bernela, M.; Ahuja, M.; Thakur, R. Enhancement of Anti-Inflammatory Activity of Glycyrrhizic Acid by Encapsulation in Chitosan-Katira Gum Nanoparticles. Eur. J. Pharm. Biopharm. 2016, 105, 141–147. [Google Scholar] [CrossRef]
- Tong, T.; Hu, H.; Zhou, J.; Deng, S.; Zhang, X.; Tang, W.; Fang, L.; Xiao, S.; Liang, J. Glycyrrhizic-Acid-Based Carbon Dots with High Antiviral Activity by Multisite Inhibition Mechanisms. Small 2020, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, B.; Wang, S.; Liang, Q.; Cai, Y.; Yang, F.; Li, G. Formulation and Evaluation of Novel Glycyrrhizic Acid Micelles for Transdermal Delivery of Podophyllotoxin. Drug Deliv. 2016, 23, 1623–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, T.; Han, H.S.; Jeong, J.; Park, E.M.; Shim, K.S. A Novel Computational Approach for the Discovery of Drug Delivery System Candidates for COVID-19. Int. J. Mol. Sci. 2021, 22, 2815. [Google Scholar] [CrossRef]



| Nanomaterial | Composition | Main Results | References |
|---|---|---|---|
| Nanoparticles | Glycyrrhizic acid-based nanoparticle suspension | GA nanoparticle inhibited the activity of LPS-induced inflammatory cytokine (NO, PGE2, TNF-α, and IL-6) production in macrophage cells. | [71] |
| Nanoparticles | 10-hydroxycamptothecin (HCPT)-loaded glycyrrhizic acid-conjugated bovine serum albumin nanoparticles | GA-BSA-HCPT nanoparticles can target liver tumor cells. | [72] |
| Nanoparticles | pDNA-polyethylenimine-glycyrrhizic acid | pDNA/PEI/GL showed high gene expressions in the liver, especially in parenchymal cells after intravenous administration. | [66] |
| Nanoparticles | Chitosan-katira gum nanoparticles | GA encapsulated in chitosan-katira gum nanoparticles enhanced its anti-inflammatory activity. | [75] |
| Nanoparticles | Hyaluronic acid-glycyrrhizic acid succinate copolymers | Enhanced liver-targeting, and all the copolymers presented no significant cytotoxicity to HepG2 cells. | [21] |
| Nanoparticles | Glycyrrhizic acid-functionalized graphene oxide | GA-GO@DOX induced mitochondria-mediated apoptosis (MMA) of cancer cells. | [70] |
| Nanoparticles | Glycyrrhizic acid-loaded pH-sensitive poly-(lactic-co-glycolic acid) | GA-loaded nanoparticles release GA to the colon and treat bowel mucosal inflammation. | [74] |
| Nanoparticles | Glycyrrhizic acid | GANPs had antiviral, anti-inflammatory, and antioxidant effects in vitro and in vivo. | [30] |
| Micelles | Paclitaxel-loaded glycyrrhizic acid micelles | PTX-loaded GA micelles demonstrated a significant improvement in the pharmacokinetic parameters of PTX after oral administration. | [73] |
| Micelles | Podophyllotoxin-loaded glycyrrhizic acid micelles | The POD-loaded GA micelles caused less skin inflammation than traditional POD tincture. | [77] |
| Liposomes | Pegylatedliposomes | Nano-liposome encapsulation of silibinin with glycyrrhizic acid increased the biological activity of the free drugs. | [65] |
| Micelles | Glycyrrhizic acid-nafamostat mesilate | A computational method for screening candidates for drug delivery systems selected GA and nafamostat mesilate (NM), which were converted into micelle nanoparticles to improve drug stability and to effectively treat COVID-19. | [78] |
| Carbon dots | Glycyrrhizic acid-based carbon dots | Biological experiments demonstrated that Gly-CDs have excellent antiviral activity against the porcine reproductive and respiratory syndrome virus (PRRSV). | [76] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimento, M.H.M.d.; de Araújo, D.R. Exploring the Pharmacological Potential of Glycyrrhizic Acid: From Therapeutic Applications to Trends in Nanomedicine. Future Pharmacol. 2022, 2, 1-15. https://doi.org/10.3390/futurepharmacol2010001
Nascimento MHMd, de Araújo DR. Exploring the Pharmacological Potential of Glycyrrhizic Acid: From Therapeutic Applications to Trends in Nanomedicine. Future Pharmacology. 2022; 2(1):1-15. https://doi.org/10.3390/futurepharmacol2010001
Chicago/Turabian StyleNascimento, Mônica Helena Monteiro do, and Daniele Ribeiro de Araújo. 2022. "Exploring the Pharmacological Potential of Glycyrrhizic Acid: From Therapeutic Applications to Trends in Nanomedicine" Future Pharmacology 2, no. 1: 1-15. https://doi.org/10.3390/futurepharmacol2010001