
Essential Fatty Acid Associated with Heart Rate Variability in Highly Trained Male Cross-Country Skiers: A Pilot Study

Abstract
:1. Introduction
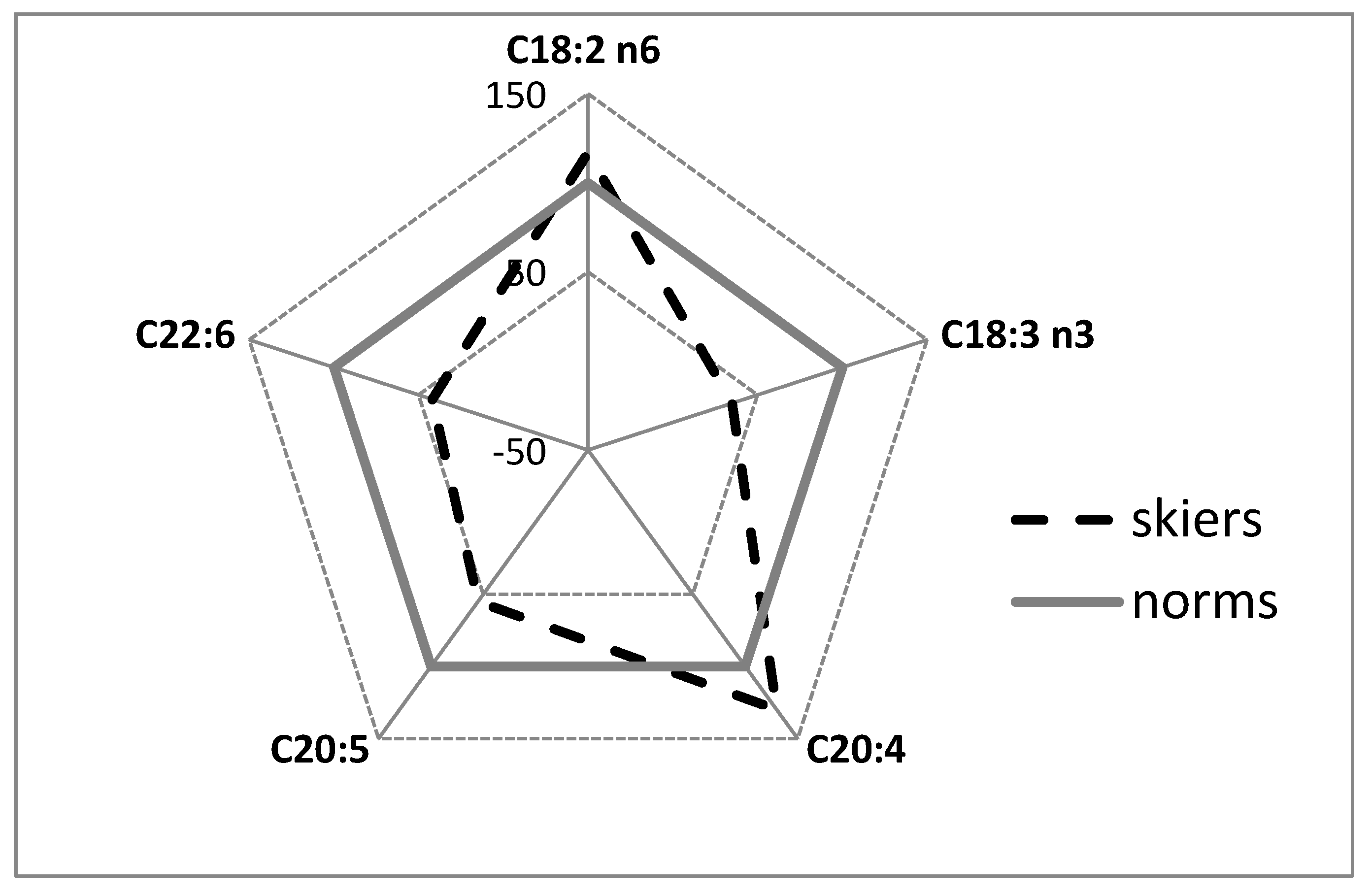
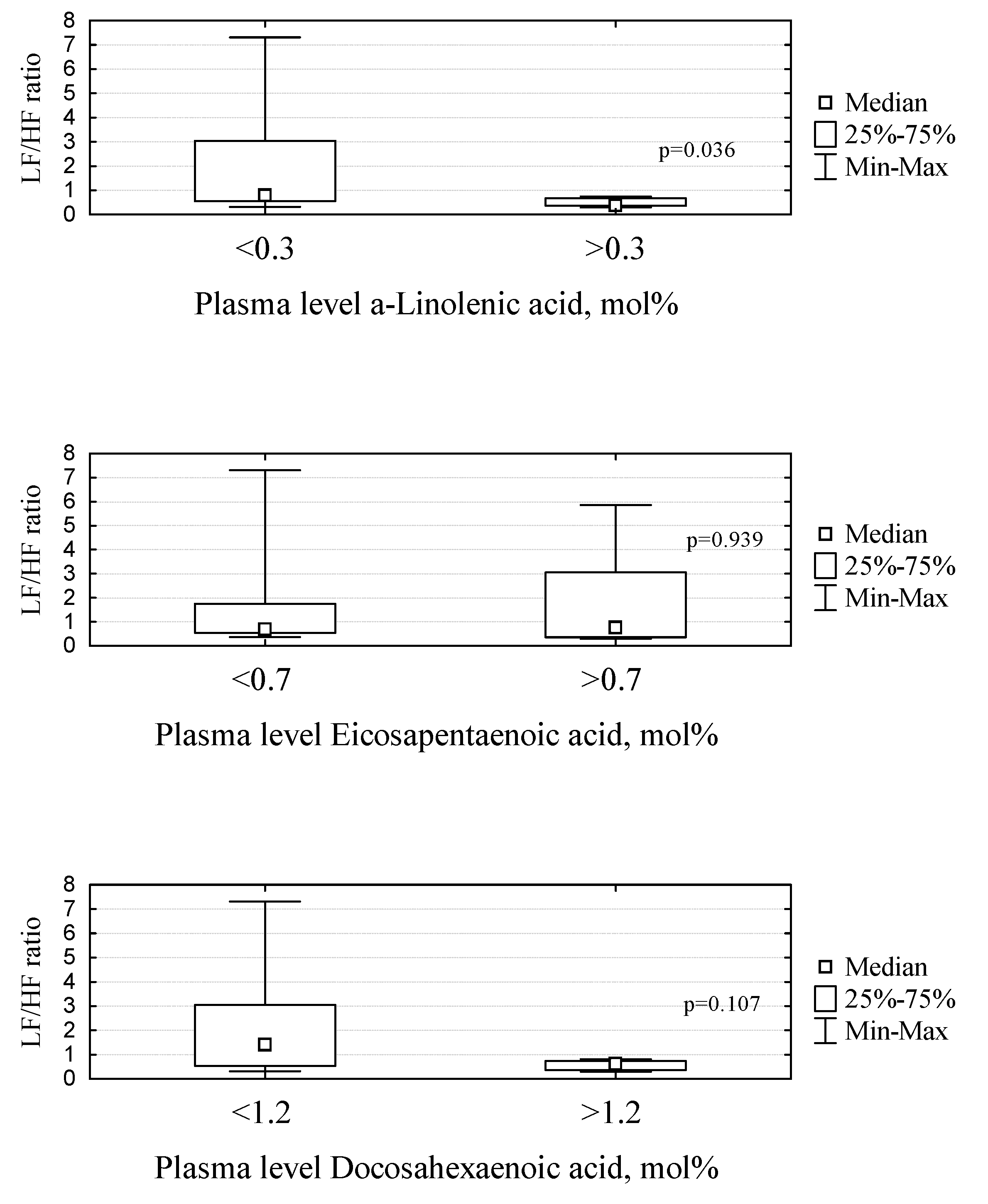
2. Results
3. Discussion
Limitations
4. Materials and Methods
4.1. Study Participants
4.2. Procedures
4.2.1. Body Composition Assessment
4.2.2. Heart Rate Variability
4.2.3. Measurement of Fatty Acids
4.2.4. Questionnaire of Fats (QFat)
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malik, M.; Bigger, J.T.; Camm, A.J.; Kleiger, R.E.; Malliani, A.; Moss, A.J.; Schwartz, P.J. Heart rate variability. Standards of measurement, physiological interpretation, and clinical use. Eur. Heart J. 1996, 17, 354–381. [Google Scholar] [CrossRef]
- Baevsky, R.M.; Baevsky, A.R.; Bersenev, E.Y.; Isaeva, O.N.; Pougachev, V.I.; Chernikova, A.G. Development of space technologies and problems of “home medicine”. Cardiometry 2015, 6, 30–40. [Google Scholar] [CrossRef]
- Markov, A.L. Heart rate variability in cross-country skiers with different level of maximum oxygen consumption. Ekol. Cheloveka Hum. Ecol. 2022, 29, 741–748. [Google Scholar] [CrossRef]
- Billman, G.E. The effects of omega-3 polyunsaturated fatty acids on cardiac rhythm: A critical reassessment. Pharmacol. Ther. 2013, 140, 53–80. [Google Scholar] [CrossRef] [PubMed]
- Drewery, M.L.; Spedale, S.B.; Lammi-Keefe, C.J. Modulation of heart rate and heart rate variability by n-3 long chain polyunsaturated fatty acids: Speculation on mechanism(s). Med. Hypotheses 2017, 107, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, H.; Larson, M.G.; Venditti, F.J.; Manders, E.S.; Evans, J.C.; Feldman, C.L.; Levy, D. Impact of reduced heart rate variability on risk for cardiac events. The Framingham Heart Study. Circulation 1996, 94, 2850–2855. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, D.; Gjerdalen, G.F.; Solberg, E.E.; Khokhlova, M.; Badtieva, V.; Herzig, D.; Trachsel, L.D.; Noack, P.; Karavirta, L.; Eser, P.; et al. Sex differences in heart rate variability: A longitudinal study in international elite cross-country skiers. Eur. J. Appl. Physiol. 2015, 115, 2107–2114. [Google Scholar] [CrossRef]
- Schmitt, L.; Willis, S.J.; Coulmy, N.; Millet, G.P. Effects of different training intensity distributions between elite cross-country skiers and nordic-combined athletes during live high-train low. Front. Physiol. 2018, 9, 932. [Google Scholar] [CrossRef]
- Lyudinina, A.Y.; Ivankova, G.E.; Bojko, E.R. Priority use of medium-chain fatty acids during high-intensity exercise in cross country skiers. J. Int. Soc. Sports Nutr. 2018, 15, 2–8. [Google Scholar] [CrossRef]
- Zhou, Y.; Khan, H.; Xiao, J.; Cheang, W.S. Effects of Arachidonic Acid Metabolites on Cardiovascular Health and Disease. Int. J. Mol. Sci. 2021, 22, 12029. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 fatty acids, exercise, physical activity and athletics. World Rev. Nutr. Diet. 2008, 98, 23–50. [Google Scholar] [CrossRef] [PubMed]
- Christensen, J.H. Omega-3 polyunsaturated fatty acids and heart rate variability. Front. Physiol. 2011, 2, 84. [Google Scholar] [CrossRef] [PubMed]
- Christensen, J.H.; Schmidt, E.B.; Molenberg, D.; Toft, E. Alpha-linolenic acid and heart rate variability in women examined for coronary artery disease. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Geelen, A.; Brouwer, I.A.; Geleijnse, J.M.; Zock, P.L.; Katan, M.B. Effect of fish oil on heart rate in humans: A meta-analysis of randomized controlled trials. Circulation 2005, 112, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, I.A.; Zock, P.L.; van Amelsvoort, L.G.; Katan, P.M.; Schouten, E.G. Association between n-3 fatty acid status in blood and electrocardiographic predictors of arrhythmia risk in healthy volunteers. Am. J. Cardiol. 2002, 89, 629–631. [Google Scholar] [CrossRef]
- Ebbesson, S.O.; Devereux, R.B.; Cole, S.; Ebbesson, L.O.; Fabsitz, R.R.; Haack, K.; Harris, W.S.; Howard, W.J.; Laston, S.; Lopez-Alvarenga, J.C.; et al. Heart rate is associated with red blood cell fatty acid concentration: The Genetics of Coronary Artery Disease in Alaska Natives (GOCADAN) study. Am. Heart J. 2010, 159, 1020–1025. [Google Scholar] [CrossRef]
- Xin, W.; Wei, W.; Li, X.Y. Short-term effects of fish-oil supplementation on heart rate variability in humans: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2013, 97, 926–935. [Google Scholar] [CrossRef]
- McLennan, P.L. Cardiac physiology and clinical efficacy of dietary fish oil clarified through cellular mechanisms of omega-3 polyunsaturated fatty acids. Eur. J. Appl. Physiol. 2014, 114, 1333–1356. [Google Scholar] [CrossRef]
- Barceló-Coblijn, G.; Murphy, E.J. Alpha-linolenic acid and its conversion to longer chain n-3 fatty acids: Benefits for human health and a role in maintaining tissue n-3 fatty acid levels. Prog. Lipid Res. 2009, 48, 355–374. [Google Scholar] [CrossRef]
- Burdge, G.C.; Calder, P.C. Dietary α-linolenic acid and health-related outcomes: A metabolic perspective. Nutr. Res. Rev. 2006, 19, 26–52. [Google Scholar] [CrossRef]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef]
- Zhang, T.; Yeung, S.L.A.; Schooling, C.M. Associations of Arachidonic Acid Synthesis with Cardiovascular Risk Factors and Relation to Ischemic Heart Disease and Stroke: A Univariable and Multivariable Mendelian Randomization Study. Nutrients 2021, 13, 1489. [Google Scholar] [CrossRef]
- Hamaad, A.; Lee, W.K.; Lip, G.Y.; MacFadyen, R.J. Oral omega n3-PUFA therapy (Omacor) has no impact on indices of heart rate variability in stable post myocardial infarction patients. Cardiovasc. Drugs Ther. 2006, 20, 359–364. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, M.K.; Lee, H.Y.; Kang, H.Y.; Kim, Y.J.; Kim, H.S. Prospective randomized comparison between omega-3 fatty acid supplements plus simvastatin versus simvastin alone in Korean patients with mixed dyslipidemia: Lipoprotein profiles and heart rate variability. Eur. J. Clin. Nutr. 2011, 65, 110–116. [Google Scholar] [CrossRef]
- Vedtofte, M.S.; Jakobsen, M.U.; Lauritzen, L.; Heitmann, B.L. Dietary α-linolenic acid, linoleic acid, and n-3 long-chain PUFA and risk of ischemic heart disease. Am. J. Clin. Nutr. 2011, 94, 1097–1103. [Google Scholar] [CrossRef]
- Nishizaki, T.; Ikeuchi, Y.; Matsuoka, T.; Sumikawa, K. Short-term depression and long-term enhancement of ACh-gated channel currents induced by linoleic and linolenic acid. Brain Res. 1997, 751, 253–258. [Google Scholar] [CrossRef]
- Ayalew-Pervanchon, A.; Rousseau, D.; Moreau, D.; Assayag, P.; Weill, P.; Grynberg, A. Long-term effect of dietary {alpha}-linolenic acid or decosahexaenoic acid on incorporation of decosahexaenoic acid in membranes and its influence on rat heart in vivo. American journal of physiology. Heart Circ. Physiol. 2007, 293, 2296–2304. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Stein, P.K.; Prineas, R.J.; Siscovick, D.S. Dietary fish and omega-3 fatty acid consumption and heart rate variability in US adults. Circulation 2008, 117, 1130–1137. [Google Scholar] [CrossRef]
- Guizy, M.; David, M.; Arias, C.; Zhang, L.; Cofán, M.; Ruiz-Gutiérrez, V.; Ros, E.; Lillo, M.P.; Martens, J.R.; Valenzuela, C. Modulation of the atrial specific Kv1.5 channel by the n-3 polyunsaturated fatty acid, alpha-linolenic acid. J. Mol. Cell. Cardiol. 2008, 44, 323–335. [Google Scholar] [CrossRef]
- Wilson, P.B.; Madrigal, L.A. Associations between whole blood and dietary omega-3 polyunsaturated fatty acid levels in collegiate athletes. Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 497–505. [Google Scholar] [CrossRef]
- La Rovere, M.T.; Staszewsky, L.; Barlera, S.; Maestri, R.; Mezzani, A.; Midi, P.; Marchioli, R.; Maggioni, A.P.; Tognoni, G.; Tavazzi, L.; et al. n-3PUFA and Holter-derived autonomic variables in patients with heart failure: Data from the Gruppo Italiano per lo Studio della Sopravvivenza nell’Insufficienza Cardiaca (GISSI-HF) Holter substudy. Heart Rhythm 2013, 10, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Lyudinina, A.Y.; Bushmanova, E.A.; Varlamova, N.G.; Boyko, E.R. Dietary and plasma blood α-linolenic acid as modulators of fat oxidation and predictors of aerobic performance. J. Int. Soc. Sports Nutr. 2020, 17, 57. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Parameters | Median (25–75th Percentiles) |
|---|---|
| HR (bpm) | 57.00 (51.50–64.00) |
| pNN50 (%) | 45.40 (28.60–54.50) |
| SDNN (ms) | 65.44 (60.11–87.37) |
| TP (ms2) | 3978.56 (3057.90–5972.84) |
| HF (ms2) | 1562.33 (988.92–1742.63) |
| LF (ms2) | 1177.46 (899.86–2166.45) |
| HF, % | 49.00 (25.75–57.55) |
| LF, % | 38.10 (31.15–53.40) |
| LF/HF | 0.79 (0.55–1.86) |
| Parameters | Alpha-Linolenic Acid (ALA) | Eicosapentaenoic Acid (EPA) | Docosahexaenoic Acid (DHA) | Linoleic Acid (LA) | Arachidonic Acid (ARA) | n6/n3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spearman, r | p-Level | Spearman, r | p-Level | Spearman, r | p-Level | Spearman, r | p-Level | Spearman, r | p-Level | Spearman, r | p-Level | |
| HR (bpm) | 0.11 | 0.652 | −0.47 | 0.037 | −0.49 | 0.026 | 0.23 | 0.335 | 0.29 | 0.215 | 0.575 | 0.009 |
| pNN50 (%) | 0.21 | 0.395 | 0.16 | 0.498 | 0.20 | 0.391 | −0.17 | 0.478 | −0.14 | 0.569 | −0.44 | 0.060 |
| SDNN (ms) | −0.28 | 0.247 | 0.14 | 0.569 | −0.02 | 0.919 | −0.18 | 0.454 | −0.28 | 0.222 | −0.18 | 0.454 |
| TP (ms2) | −0.41 | 0.077 | 0.12 | 0.613 | −0.08 | 0.743 | −0.27 | 0.237 | −0.36 | 0.113 | −0.13 | 0.586 |
| HF (ms2) | 0.16 | 0.509 | 0.05 | 0.835 | 0.10 | 0.663 | 0.10 | 0.673 | 0.27 | 0.256 | −0.26 | 0.283 |
| LF (ms2) | −0.49 | 0.028 | 0.13 | 0.537 | −0.10 | 0.672 | −0.27 | 0.256 | −0.38 | 0.111 | −0.07 | 0.775 |
| HF, % | 0.46 | 0.045 | −0.14 | 0.552 | 0.20 | 0.392 | 0.29 | 0.204 | 0.59 | 0.006 | −0.10 | 0.678 |
| LF, % | −0.48 | 0.038 | 0.04 | 0.865 | −0.18 | 0.442 | −0.24 | 0.308 | −0.52 | 0.022 | 0.11 | 0.639 |
| LF/HF | −0.49 | 0.032 | 0.08 | 0.724 | −0.19 | 0.423 | −0.25 | 0.291 | −0.54 | 0.017 | 0.11 | 0.639 |
| Characteristics | Median (25–75th Percentiles) |
|---|---|
| Age, years | 18.0 (18.0–29) |
| Body height, cm | 180.0 (167.0–190.1) |
| Body mass, kg | 70.3 (57.0–77.0) |
| Body mass index, kg/m2 | 22.5 (18.5–24.3) |
| Fat mass, % | 9.5 (4.8–13.9) |
| Fat-free mass, kg | 65.7 (48.5–68.8) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyudinina, A.Y.; Markov, A.L.; Bojko, E.R. Essential Fatty Acid Associated with Heart Rate Variability in Highly Trained Male Cross-Country Skiers: A Pilot Study. Physiologia 2024, 4, 54-63. https://doi.org/10.3390/physiologia4010003
Lyudinina AY, Markov AL, Bojko ER. Essential Fatty Acid Associated with Heart Rate Variability in Highly Trained Male Cross-Country Skiers: A Pilot Study. Physiologia. 2024; 4(1):54-63. https://doi.org/10.3390/physiologia4010003
Chicago/Turabian StyleLyudinina, Aleksandra Y., Alexander L. Markov, and Evgeny R. Bojko. 2024. "Essential Fatty Acid Associated with Heart Rate Variability in Highly Trained Male Cross-Country Skiers: A Pilot Study" Physiologia 4, no. 1: 54-63. https://doi.org/10.3390/physiologia4010003