
Ecology Meets Physiology: Phenotypic Plasticity and the Ability of Animals to Adjust to Changing Environmental Conditions

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
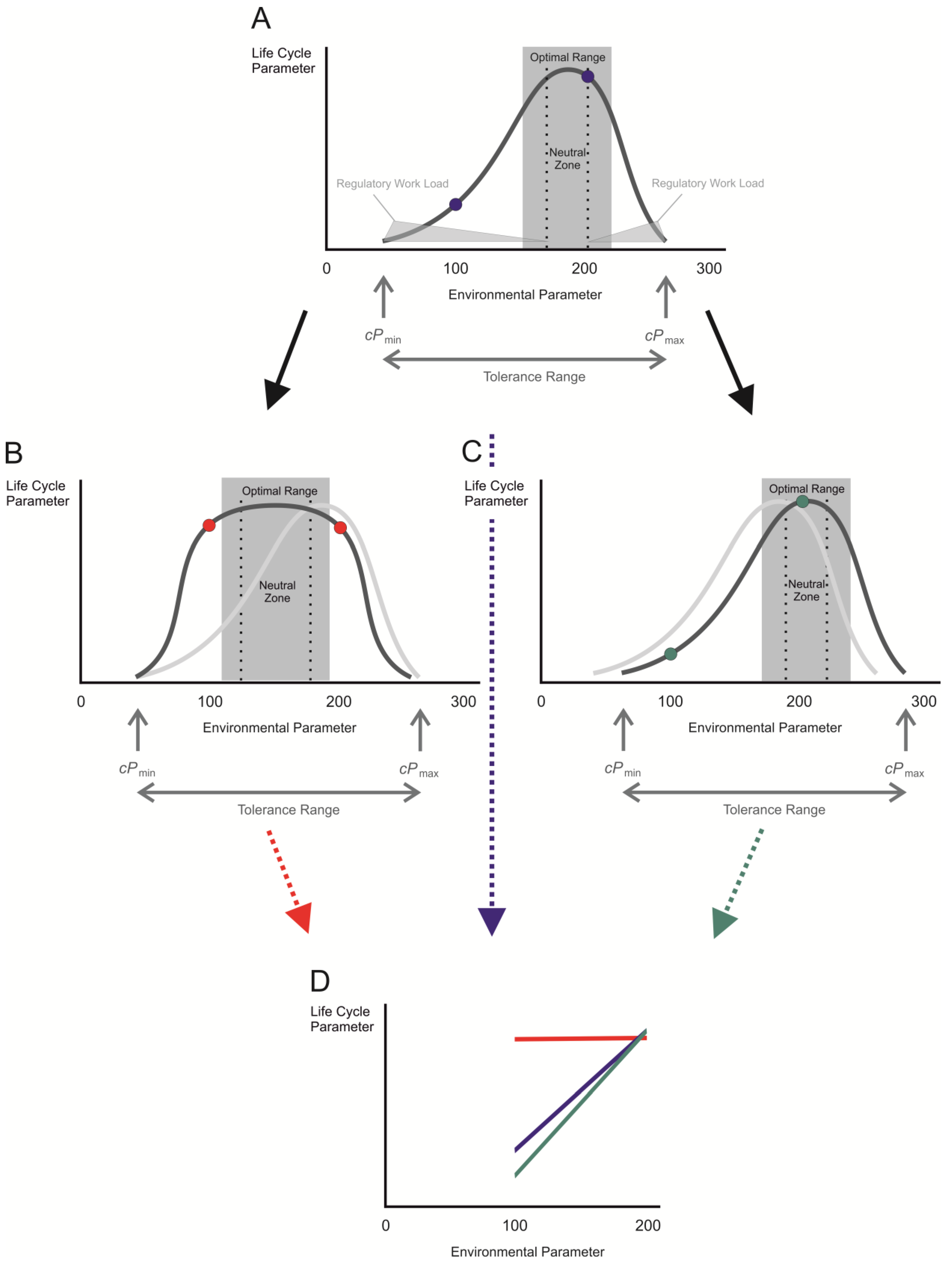
2. Measure for Plasticity in Response to Environmental Change: The Reaction Norm
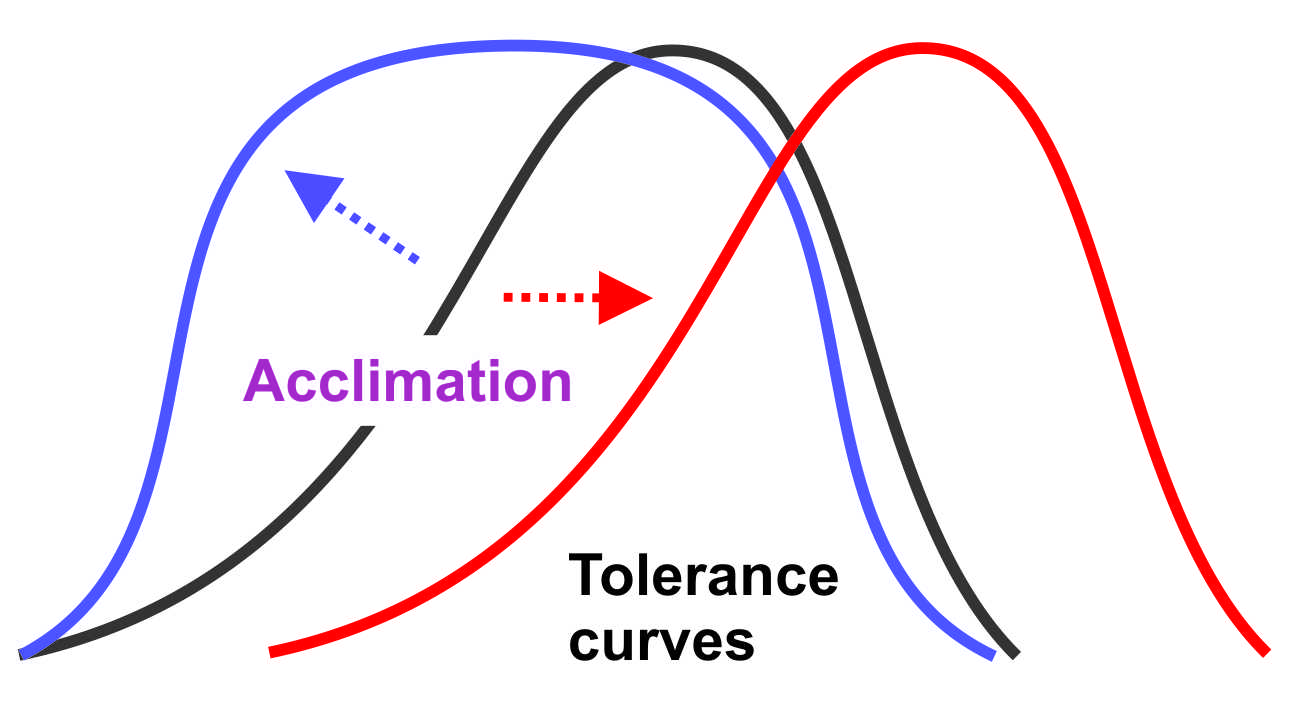
3. Acclimation
4. Reversibility of Plastic Changes
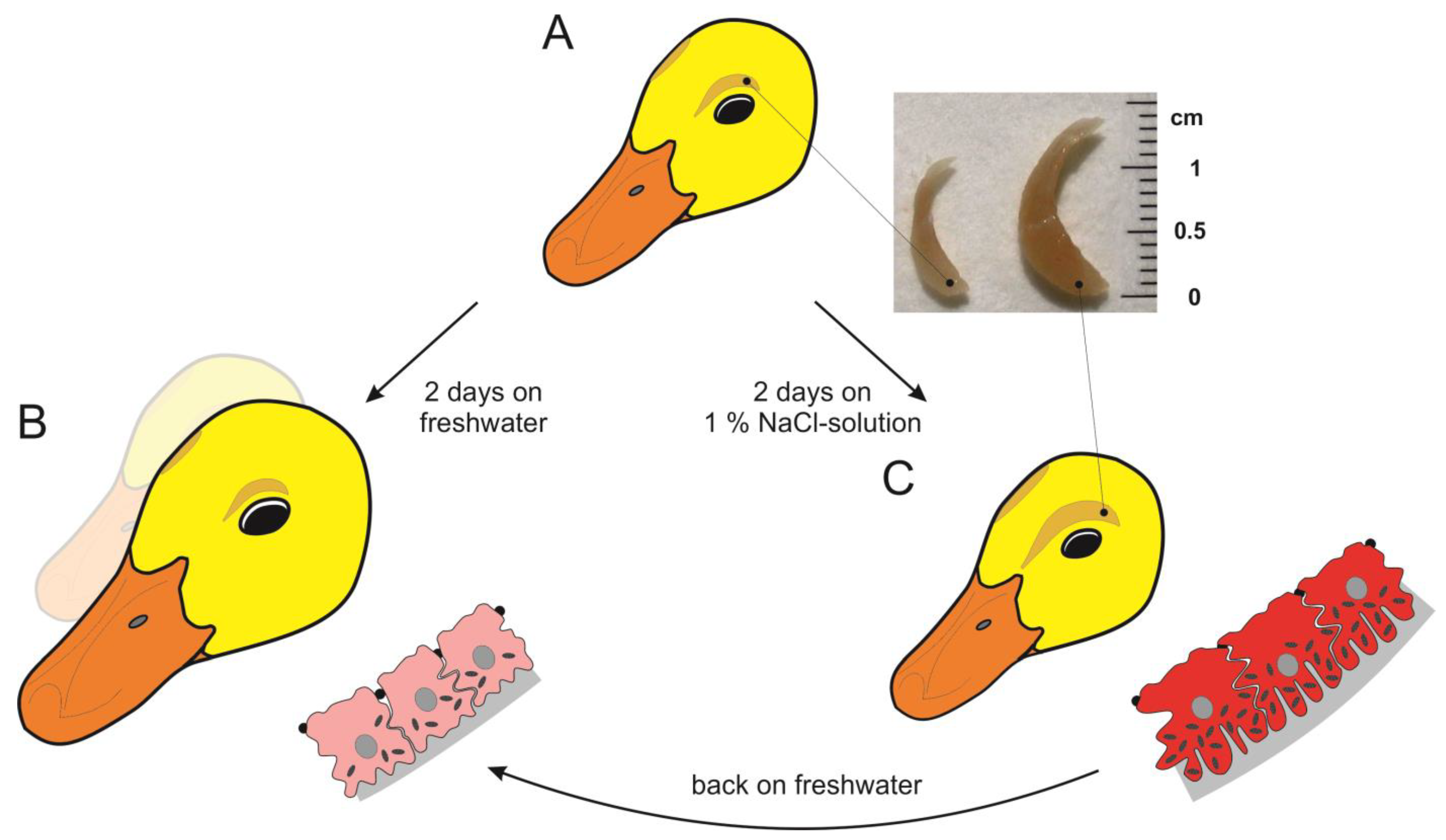
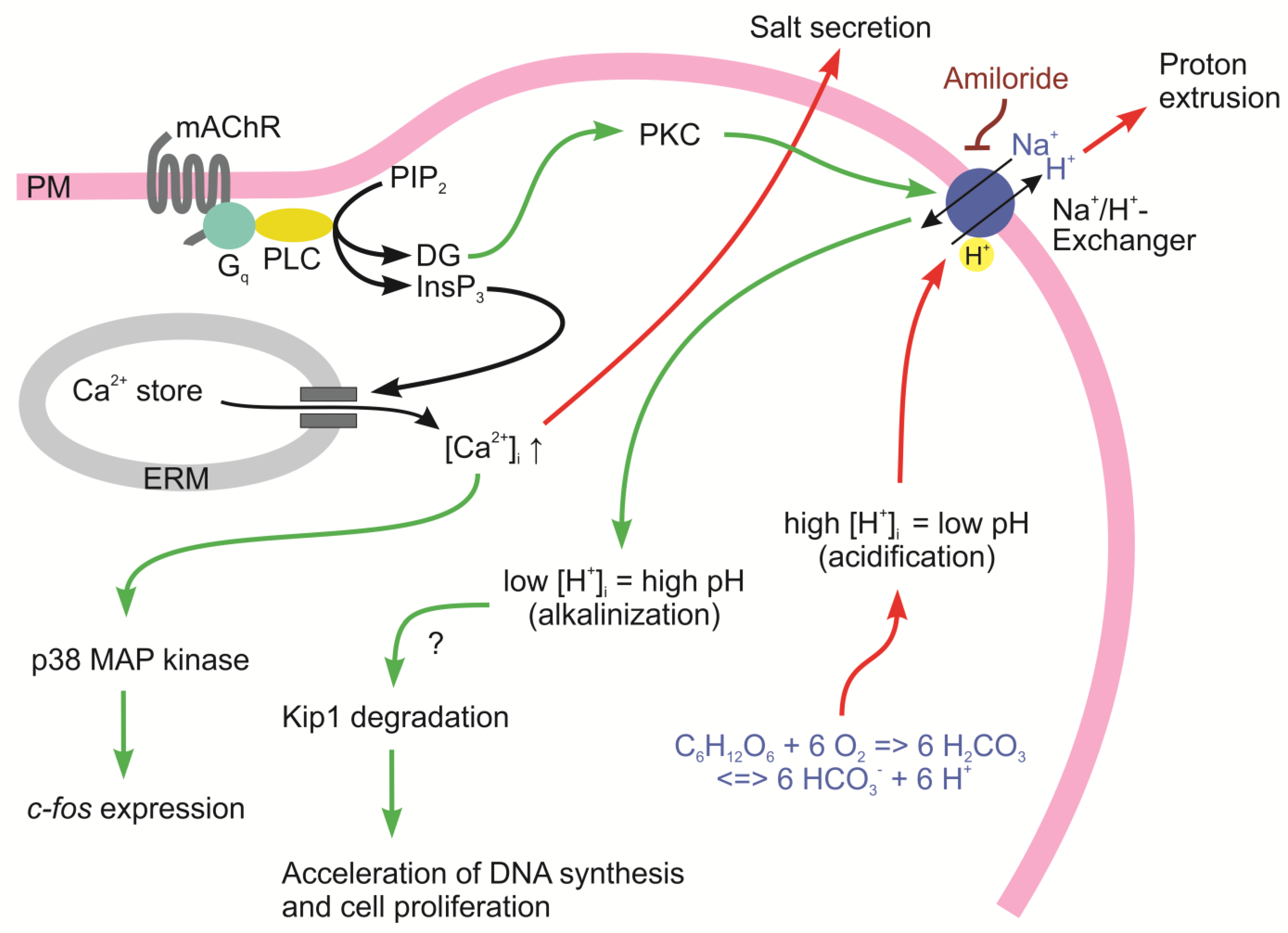
5. The Avian Salt Gland as an Example of Environmentally Induced Irreversible and Reversible Phenotypic Changes
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pigliucci, M.; Schlichting, C.D.; Jones, C.S.; Schwenk, K. Developmental reaction norms: The interactions among allometry, ontogeny and plasticity. Plant Species Biol. 1996, 11, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Weinkove, D.; Leevers, S.J. The genetic control of organ growth: Insights from Drosophila. Curr. Opin. Genet. Dev. 2000, 10, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Ferenci, T.; Maharjan, R. Mutational heterogeneity: A key ingredient of bet-hedging and evolutionary divergence? Bioessays 2015, 37, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Hoban, S.; Kelley, J.L.; Lotterhos, K.E.; Antolin, M.F.; Bradburd, G.; Lowry, D.B.; Poss, M.L.; Reed, L.K.; Storfer, A.; Whitlock, M.C. Finding the genomic basis of local adaptation: Pitfalls, practical solutions, and future directions. Am. Nat. 2016, 188, 379–397. [Google Scholar] [CrossRef] [Green Version]
- Charmantier, A.; Garant, D. Environmental quality and evolutionary potential: Lessons from wild populations. Proc. Biol. Sci. 2005, 272, 1415–1425. [Google Scholar] [CrossRef] [Green Version]
- Woods, H.A. Mosaic physiology from developmental noise: Within-organism physiological diversity as an alternative to phenotypic plasticity and phenotypic flexibility. J. Exp. Biol. 2014, 217, 35–45. [Google Scholar] [CrossRef] [Green Version]
- de Villemereuil, P.; Gaggiotti, O.E.; Mouterde, M.; Till-Bottraud, I. Common garden experiments in the genomic era: New perspectives and opportunities. Heredity 2016, 116, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Tejedor, J.R.; Fraga, M.F. Interindividual epigenetic variability: Sound or noise? Bioessays 2017, 39, 1700055. [Google Scholar] [CrossRef]
- Cossins, A.; Fraser, J.; Hughes, M.; Gracey, A. Post-genomic approaches to understanding the mechanisms of environmentally induced phenotypic plasticity. J. Exp. Biol. 2006, 209, 2328–2336. [Google Scholar] [CrossRef] [Green Version]
- Pigliucci, M.; Murren, C.J.; Schlichting, C.D. Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 2006, 209, 2362–2367. [Google Scholar] [CrossRef] [Green Version]
- Fusco, G.; Minelli, A. Phenotypic plasticity in development and evolution: Facts and concepts. Philos. Trans. R. Soc. B 2010, 365, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaman, J.E.; White, C.R.; Seebacher, F. Evolution of plasticity: Mechanistic link between development and reversible acclimation. Trends Ecol. Evol. 2016, 31, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Chevin, L.-M.; Collins, S.; Lefèvre, F. Phenotypic plasticity and evolutionary demographic responses to climate change: Taking theory out to the field. Funct. Ecol. 2013, 27, 967–979. [Google Scholar] [CrossRef] [Green Version]
- Minelli, A.; Fusco, G. Developmental plasticity and the evolution of animal complex life cycles. Philos. Trans. R. Soc. B 2010, 365, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Moczek, A.P.; Sultan, S.; Foster, S.; Ledon-Rettig, C.; Dworkin, I.; Nijhout, H.F.; Abouheif, E.; Pfennig, D.W. The role of developmental plasticity in evolutionary innovation. Proc. Biol. Sci. 2011, 278, 2705–2713. [Google Scholar] [CrossRef] [Green Version]
- Peirson, B.R.E. Plasticity, stability, and yield: The origins of Anthony David Bradshaw’s model of adaptive phenotypic plasticity. Stud. Hist. Philos. Sci. Part C Stud. Hist. Philos. Biol. Biomed. Sci. 2015, 50, 51–66. [Google Scholar] [CrossRef]
- Pfennig, D.W.; McGee, M. Resource polyphenism increases species richness: A test of the hypothesis. Philos. Trans. R. Soc. B 2010, 365, 577–591. [Google Scholar] [CrossRef] [Green Version]
- Schlichting, C.D.; Smith, H. Phenotypic plasticity: Linking molecular mechanisms with evolutionary outcomes. Evol. Ecol. 2002, 16, 189–211. [Google Scholar] [CrossRef]
- Sommer, R.J. Phenotypic plasticity: From theory and genetics to current and future challenges. Genetics 2020, 215, 1–13. [Google Scholar] [CrossRef]
- Joschinski, J.; Bonte, D. Transgenerational plasticity and bet-hedging: A framework for reaction norm evolution. Front. Ecol. Evol. 2020, 8, 517183. [Google Scholar] [CrossRef]
- Waldvogel, A.-M.; Feldmeyer, B.; Rolshausen, G.; Exposito-Alonso, M.; Rellstab, C.; Kofler, R.; Mock, T.; Schmid, K.; Schmitt, I.; Bataillon, T.; et al. Evolutionary genomics can improve prediction of species’ responses to climate change. Evol. Lett. 2020, 4, 4–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroi, A.M.; Bennett, A.F.; Lenski, R.E. Temperature acclimation and competitive fitness: An experimental test of the beneficial acclimation assumption. Proc. Natl. Acad. Sci. USA 1994, 91, 1917–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacio-Lopez, K.; Beckage, B.; Scheiner, S.; Molofsky, J. The ubiquity of phenotypic plasticity in plants: A synthesis. Ecol. Evol. 2015, 5, 3389–3400. [Google Scholar] [CrossRef] [PubMed]
- van Kleunen, M.; Fischer, M. Constraints on the evolution of adaptive phenotypic plasticity in plants. New Phytol. 2005, 166, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S.; Franklin, C.E. The detrimental acclimation hypothesis—Response. Trends Ecol. Evol. 2002, 17, 408. [Google Scholar] [CrossRef]
- Acasuso-Rivero, C.; Murren, C.J.; Schlichting, C.D.; Steiner, U.K. Adaptive phenotypic plasticity for life-history and less fitness-related traits. Proc. Biol. Sci. 2019, 286, 20190653. [Google Scholar] [CrossRef] [Green Version]
- Loeschcke, V.; Hoffmann, A.A. The detrimental acclimation hypothesis. Trends Ecol. Evol. 2002, 17, 407–408. [Google Scholar] [CrossRef]
- Lalejini, A.; Ferguson, A.J.; Grant, N.A.; Ofria, C. Adaptive phenotypic plasticity stabilizes evolution in fluctuating environments. Front. Ecol. Evol. 2021, 9, 715381. [Google Scholar] [CrossRef]
- Romero-Mujalli, D.; Rochow, M.; Kahl, S.; Paraskevopoulou, S.; Folkertsma, R.; Jeltsch, F.; Tiedemann, R. Adaptive and nonadaptive plasticity in changing environments: Implications for sexual species with different life history strategies. Ecol. Evol. 2021, 11, 6341–6357. [Google Scholar] [CrossRef]
- Gabriel, W.; Lynch, M. The selective advantage of reaction norms for environmental tolerance. J. Evol. Biol. 1992, 5, 41–59. [Google Scholar] [CrossRef] [Green Version]
- Scharloo, W. Developmental and physiological aspects of reaction norms—Drosophila data link genetic variation and phenotypic response to the environment. BioScience 1989, 39, 465–471. [Google Scholar] [CrossRef]
- Schulte, P.M. What is environmental stress Insights from fish living in a variable environment. J. Exp. Biol. 2014, 217, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ecker, S.; Pancaldi, V.; Valencia, A.; Beck, S.; Paul, D.S. Epigenetic and transcriptional variability shape phenotypic plasticity. Bioessays 2018, 40, 1700148. [Google Scholar] [CrossRef]
- Conlon, I.; Raff, M. Size control in animal development. Cell 1999, 96, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costanzo, J.P.; Lee, R.E. Avoidance and tolerance of freezing in ectothermic vertebrates. J. Exp. Biol. 2013, 216, 1961–1967. [Google Scholar] [CrossRef] [Green Version]
- Duman, J.G. Animal ice-binding (antifreeze) proteins and glycolipids: An overview with emphasis on physiological function. J. Exp. Biol. 2015, 218, 1846–1855. [Google Scholar] [CrossRef] [Green Version]
- Kültz, D. Molecular and evolutionary basis of the cellular stress response. Annu. Rev. Physiol. 2005, 67, 225–257. [Google Scholar] [CrossRef]
- Mymrikov, E.V.; Seit-Nebi, A.S.; Gusev, N.B. Large potentials of small heat shock proteins. Physiol. Rev. 2011, 91, 1123–1159. [Google Scholar] [CrossRef] [Green Version]
- Prosser, C.L. Adaptational Biology: Molecules to Organisms; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Bradshaw, W.E.; Holzapfel, C.M. Light, time, and the physiology of biotic response to rapid climate change in animals. Annu. Rev. Physiol. 2010, 72, 147–166. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, J.J.C. The emerging role of RNA editing in plasticity. J. Exp. Biol. 2015, 218, 1812–1821. [Google Scholar] [CrossRef] [Green Version]
- Lamka, G.F.; Harder, A.M.; Sundaram, M.; Schwartz, T.S.; Christie, M.R.; DeWoody, J.A.; Willoughby, J.R. Epigenetics in ecology, evolution, and conservation. Front. Ecol. Evol. 2022, 10, 871791. [Google Scholar] [CrossRef]
- Bryant, P.J.; Simpson, P. Intrinsic and extrinsic control of growth in developing organs. Q. Rev. Biol. 1984, 59, 387–415. [Google Scholar] [CrossRef]
- Crickmore, M.A.; Mann, R.S. The control of size in animals: Insights from selector genes. Bioessays 2008, 30, 843–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafen, E.; Stocker, H. How are the sizes of cells, organs, and bodies controlled? PLoS Biol. 2003, 1, e86. [Google Scholar] [CrossRef] [Green Version]
- Nijhout, H.F.; Riddiford, L.M.; Mirth, C.; Shingleton, A.W.; Suzuki, Y.; Callier, V. The developmental control of size in insects. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 113–134. [Google Scholar] [CrossRef] [PubMed]
- Parker, J. Morphogens, nutrients, and the basis of organ scaling. Evol. Dev. 2011, 13, 304–314. [Google Scholar] [CrossRef]
- Potter, C.J.; Xu, T. Mechanisms of size control. Curr. Opin. Genet. Dev. 2001, 11, 279–286. [Google Scholar] [CrossRef]
- Yang, X.; Xu, T. Molecular mechanism of size control in development and human diseases. Cell Res. 2011, 21, 715–729. [Google Scholar] [CrossRef]
- Lessells, C.K. Neuroendocrine control of life histories: What do we need to know to understand the evolution of phenotypic plasticity? Philos. Trans. R. Soc. B 2008, 363, 1589–1598. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, T.P.; Caldez, M.J.; Kaldis, P.; Björklund, M. Cell size control—A mechanism for maintaining fitness and function. Bioessays 2017, 39, 1700058. [Google Scholar] [CrossRef] [Green Version]
- Costantini, D.; Metcalfe, N.B.; Monaghan, P. Ecological processes in a hormetic framework. Ecol. Lett. 2010, 13, 1435–1447. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Karl, I. Exploring plastic and genetic responses to temperature variation using copper butterflies. Clim. Res. 2010, 43, 17–30. [Google Scholar] [CrossRef]
- DeWitt, T.J.; Sih, A.; Wilson, D.S. Costs and limits of phenotypic plasticity. Trends Ecol. Evol. 1998, 13, 77–81. [Google Scholar] [CrossRef]
- Fischer, K.; Eenhoorn, E.; Bot, A.N.M.; Brakefield, P.M.; Zwaan, B.J. Cooler butterflies lay larger eggs: Developmental plasticity versus acclimation. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 2051–2056. [Google Scholar] [CrossRef]
- Nijhout, H.F. Development and evolution of adaptive polyphenisms. Evol. Dev. 2003, 5, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Aubry, L.M.; Williams, C.T. Vertebrate phenological plasticity: From molecular mechanisms to ecological and evolutionary implications. Integr. Comp. Biol. 2022, 62, 958–971. [Google Scholar] [CrossRef]
- Fu, R.; Gillen, A.E.; Grabek, K.R.; Riemondy, K.A.; Epperson, L.E.; Bustamante, C.D.; Hesselberth, J.R.; Martin, S.L. Dynamic RNA regulation in the brain underlies physiological plasticity in a hibernating mammal. Front. Physiol. 2021, 11, 624677. [Google Scholar] [CrossRef]
- Piersma, T.; Drent, J. Phenotypic flexibility and the evolution of organismal design. Trends Ecol. Evol. 2003, 18, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Piersma, T.; Lindström, A. Rapid reversible changes in organ size as a component of adaptive behaviour. Trends Ecol. Evol. 1997, 12, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Piersma, T.; van Gils, J.A. The Flexible Phenotype—A Body-Centered Integration of Ecology, Physiology, and Behaviour; Oxford University Press: Oxford, NY, USA, 2011. [Google Scholar]
- Hildebrandt, J.-P.; Wiesenthal, A.A.; Müller, C. Phenotypic plasticity in animals exposed to osmotic stress—Is it always adaptive? Bioessays 2018, 40, e1800069. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K. Animal Physiology: Adaptation and Environment; Cambridge University Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Dantzler, W.H. Comparative Physiology of the Vertebrate Kidney; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Schmidt-Nielsen, K. The salt-secreting gland of marine birds. Circulation 1960, 21, 955–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Technau, G. Die Nasendrüse der Vögel. J. Ornithol. 1936, 84, 511–617. [Google Scholar] [CrossRef]
- Frizzell, R.A.; Hanrahan, J.W. Physiology of epithelial chloride and fluid secretion. Cold Spring Harb. Perspect. Med. 2012, 2, a009563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrnett, R.J.; Mazurkiewicz, J.E.; Addis, J.S. Avian salt gland: A model for the study of membrane biogenesis. Methods Enzymol. 1983, 96, 627–659. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.A.; Goertemiller, C.C.; DeLellis, R.A.; Kablotsky, Y.H. The effect of a salt water regimen on the development of the salt glands of domestic ducklings. Dev. Biol. 1963, 8, 286–308. [Google Scholar] [CrossRef]
- Ernst, S.A.; Ellis, R.A. The development of surface specialization in the secretory epithelium of the avian salt gland in response to osmotic stress. J. Cell. Biol. 1969, 40, 305–321. [Google Scholar] [CrossRef]
- Fletcher, G.L.; Stainer, I.M.; Holmes, W.N. Sequential changes in the adenosinetriphosphatase activity and the electrolyte excretory capacity of the nasal glands of the duck (Anas platyrhynchos) during the period of adaptation to hypertonic saline. J. Exp. Biol. 1967, 47, 375–391. [Google Scholar] [CrossRef]
- Hildebrandt, J.-P. Changes in Na+/K+-ATPase expression during adaptive cell differentiation in avian nasal salt gland. J. Exp. Biol. 1997, 200, 1895–1904. [Google Scholar] [CrossRef]
- Fänge, R.; Schmidt-Nielsen, K.; Robinson, M. Control of secretion from the avian salt gland. Am. J. Physiol. 1958, 195, 321–326. [Google Scholar] [CrossRef]
- Krohn, M.; Hildebrandt, J.-P. Cross-talk of phosphoinositide- and cyclic nucleotide-dependent signaling pathways in differentiating avian nasal gland cells. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2004, 174, 461–470. [Google Scholar] [CrossRef]
- Müller, C.; Hildebrandt, J.-P. Salt glands—The perfect way to get rid of too much sodium chloride. Biologist 2003, 50, 255–258. [Google Scholar]
- Bentz, C.; Schwarz, M.; Hildebrandt, J.-P. Cytosolic pH affects DNA-synthesis in nasal gland cells of osmotically stressed ducklings, Anas platyrhynchos. Zoology 1999, 102, 10–17. [Google Scholar]
- Hanwell, A.; Peaker, M. The control of adaptive hypertrophy in the salt glands of geese and ducks. J. Physiol. 1975, 248, 193–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, W.N.; Stewart, D.J. Changes in the nucleic acid and protein composition of the nasal glands from the duck (Anas platyrhynchos) during the period of adaptation to hypertonic saline. J. Exp. Biol. 1968, 48, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, J.-P. Coping with excess salt: Adaptive functions of extrarenal osmoregulatory organs in vertebrates. Zoology 2001, 104, 209–220. [Google Scholar] [CrossRef]
- Peaker, M. Salt glands in marine birds: What triggers secretion and what makes them grow? In Comparative Physiology: Water, Ions and Fluid Mechanics; Schmidt-Nielsen, K., Bolis, L., Maddrell, S.H.P., Eds.; Cambridge University Press: Cambridge, MA, USA, 1978; pp. 207–212. [Google Scholar]
- Hildebrandt, J.-P.; Shuttleworth, T.J. Muscarinic receptor characterization in differentiating avian exocrine cells. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1994, 266, R674–R681. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, J.-P.; Shuttleworth, T.J. A Gq-type G protein couples muscarinic receptors to inositol phosphate and calcium signaling in exocrine cells from the avian salt gland. J. Membr. Biol. 1993, 133, 183–190. [Google Scholar] [CrossRef]
- Hildebrandt, J.-P.; Shuttleworth, T.J. Inositol phosphates and [Ca2+]i signals in a differentiating exocrine cell. Am. J. Physiol. Cell Physiol. 1991, 261, C210–C217. [Google Scholar] [CrossRef]
- Shuttleworth, T.J. Temporal relationships between Ca2+ store mobilization and Ca2+ entry in an exocrine cell. Cell Calcium 1994, 15, 457–466. [Google Scholar] [CrossRef]
- Lowy, R.J.; Dawson, D.C.; Ernst, S.A. Primary culture of duck salt gland. II. Neurohormonal stimulation of active transport. Am. J. Physiol. 1985, 249, C41–C47. [Google Scholar] [CrossRef]
- Shuttleworth, T.J.; Hildebrandt, J.-P. Vertebrate salt glands—Short- and long-term regulation of function. J. Exp. Zool. 1999, 283, 689–701. [Google Scholar] [CrossRef]
- Hildebrandt, J.-P.; Gerstberger, R.; Schwarz, M. In vivo and in vitro induction of c-fos in avian exocrine salt gland cells. Am. J. Physiol. Cell Physiol. 1998, 275, C951–C957. [Google Scholar] [CrossRef] [PubMed]
- Dalhäusser, A.K.; Rössler, O.G.; Thiel, G. Regulation of c-fos gene transcription by stimulus-responsive protein kinases. Gene 2022, 821, 146284. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, J.-P.; Prowald, A. Ca2+ and p38 MAP kinase regulate mAChR-mediated c-Fos expression in avian exocrine cells. Am. J. Physiol. Cell Physiol. 2000, 278, C879–C884. [Google Scholar] [CrossRef] [Green Version]
- Rohlfing, A.-K.; Schill, T.; Müller, C.; Hildebrandt, P.; Prowald, A.; Hildebrandt, J.-P. Attenuation of cell cycle regulator p27(Kip1) expression in vertebrate epithelial cells mediated by extracellular signals in vivo and in vitro. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2005, 175, 511–522. [Google Scholar] [CrossRef]
- Bencivenga, D.; Stampone, E.; Roberti, D.; Della Ragione, F.; Borriello, A. p27(Kip1), an intrinsically unstructured protein with scaffold properties. Cells 2021, 10, 2254. [Google Scholar] [CrossRef]
- Bendris, N.; Lemmers, B.; Blanchard, J.M. Cell cycle, cytoskeleton dynamics and beyond: The many functions of cyclins and CDK inhibitors. Cell Cycle 2015, 14, 1786–1798. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Hunter, T. Ubiquitylation and proteasomal degradation of the p21(Cip1), p27(Kip1) and p57(Kip2) CDK inhibitors. Cell Cycle 2010, 9, 2342–2352. [Google Scholar] [CrossRef] [Green Version]
- Bentz, C.; Hildebrandt, J.-P. Adaptive differentiation of avian exocrine cells alters their pHi response to mAChR activation. Am. J. Physiol. Cell Physiol. 1995, 268, C944–C951. [Google Scholar] [CrossRef]
- Moolenaar, W.H.; Defize, L.H.; De Laat, S.W. Ionic signalling by growth factor receptors. J. Exp. Biol. 1986, 124, 359–373. [Google Scholar] [CrossRef]
- Grinstein, S.; Rotin, D.; Mason, M.J. Na+/H+ exchange and growth factor-induced cytosolic pH changes. Role in cellular proliferation. Biochim. Biophys. Acta 1989, 988, 73–97. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C. Protein kinase C: Perfectly balanced. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 208–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakabayashi, S.; Fafournoux, P.; Sardet, C.; Pouyssegur, J. The Na+/H+ antiporter cytoplasmatic domain mediates growth factor signals and controls “H+-sensing”. Proc. Natl. Acad. Sci. USA 1992, 89, 2424–2428. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S.C.; Malik, A.; Basit, H. Physiology, Exocrine Gland; StatPearls: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK542322/ (accessed on 19 April 2023).
- Khan, S.; Fitch, S.; Knox, S.; Arora, R. Exocrine gland structure-function relationships. Development 2022, 149, dev197657. [Google Scholar] [CrossRef]
- Govindarajan, V.; Ito, M.; Makarenkova, H.P.; Lang, R.A.; Overbeek, P.A. Endogenous and ectopic gland induction by FGF-10. Dev. Biol. 2000, 225, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, A.; Itoh, N.; Kato, S.; Thiery, J.P.; Czernichow, P.; Bellusci, S.; Scharfmann, R. FGF10 is essential for maintaining the proliferative capacity of epithelial progenitor cells during early pancreatic organogenesis. Development 2001, 128, 5109–5117. [Google Scholar] [CrossRef] [PubMed]
- Entesarian, M.; Matsson, H.; Klar, J.; Bergendal, B.; Olson, L.; Arakaki, R.; Hayashi, Y.; Ohuchi, H.; Falahat, B.; Bolstad, A.I.; et al. Mutations in the gene encoding fibroblast growth factor 10 are associated with aplasia of lacrimal and salivary glands. Nat. Genet. 2005, 37, 125–128. [Google Scholar] [CrossRef]
- Knosp, W.M.; Knox, S.M.; Hoffman, M.P. Salivary gland organogenesis. WIREs Dev. Biol. 2012, 1, 69–82. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, J.; Liu, Y.; Dattilo, L.K.; Huh, S.-H.; Ornitz, D.; Beebe, D.C. FGF signaling activates a Sox9-Sox10 pathway for the formation and branching morphogenesis of mouse ocular glands. Development 2014, 141, 2691–2701. [Google Scholar] [CrossRef] [Green Version]
- Chatzeli, L.; Gaete, M.; Tucker, A.S. FGF10 and SOX9 are essential for the establishment of distal progenitor cells during mouse salivary gland development. Development 2017, 144, 2294–2305. [Google Scholar] [CrossRef] [Green Version]
- Paine, I.S.; Lewis, M.T. The terminal end bud: The little engine that could. J. Mammary Gland Biol. Neopl. 2017, 22, 93–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivetti, S.; Chen, C.; Chen, C.; Bellusci, S. FGF10/FGFR2b signaling in mammary gland development, homeostasis, and cancer. Front. Cell Dev. Biol. 2020, 8, 415. [Google Scholar] [CrossRef] [PubMed]
- Seymour, P.A.; Freude, K.K.; Tran, M.N.; Mayes, E.E.; Jensen, J.; Kist, R.; Scherer, G.; Sander, M. SOX9 is required for maintenance of the pancreatic progenitor cell pool. Proc. Nat. Acad. Sci. USA 2007, 104, 1865–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athwal, H.K.; Murphy, G.; Tibbs, E.; Cornett, A.; Hill, E.; Yeoh, K.; Berenstein, E.; Hoffman, M.P.; Lombaert, I.M.A. Sox10 regulates plasticity of epithelial progenitors toward secretory units of exocrine glands. Stem Cell Rep. 2019, 12, 366–380. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.; Zhang, X. Lacrimal gland development: From signaling interactions to regenerative medicine. Dev. Dyn. 2017, 246, 970–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhang, J.; Liu, J.; Zhou, Y.; Cai, C.; Xu, L.; Dai, X.; Feng, S.; Guo, C.; Rao, J.; et al. A new duck genome reveals conserved and convergently evolved chromosome architectures of birds and mammals. Gigascience 2021, 10, giaa142. [Google Scholar] [CrossRef]
- Huang, Y.; Li, Y.; Burt, D.W.; Chen, H.; Zhang, Y.; Qian, W.; Kim, H.; Gan, S.; Zha, Y.; Li, J.; et al. The duck genome and transcriptome provide insight into an avian influenza virus reservoir species. Nat. Genet. 2013, 45, 776–783. [Google Scholar] [CrossRef] [Green Version]
- McClelland, G.B.; Scott, G.R. Muscle plasticity. In The Physiology of Fishes, 4th ed.; Evans, D.H., Claiborne, J.B., Currie, S., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 1–31. [Google Scholar]
- Pörtner, H.-O. Oxygen- and capacity-limitation of thermal tolerance: A matrix for integrating climate-related stressor effects in marine ecosystems. J. Exp. Biol. 2010, 213, 881–893. [Google Scholar] [CrossRef] [Green Version]
- Somero, G.N. Unity in diversity: A perspective on the methods, contributions, and future of comparative physiology. Ann. Rev. Physiol. 2000, 62, 927–937. [Google Scholar] [CrossRef] [Green Version]
- Fields, P.A.; Dong, Y.; Meng, X.; Somero, G.N. Adaptations of protein structure and function to temperature: There is more than one way to ‘skin a cat’. J. Exp. Biol. 2015, 218, 1801–1811. [Google Scholar] [CrossRef] [Green Version]
- Ho, D.H.; Burggren, W.W. Epigenetics and transgenerational transfer: A physiological perspective. J. Exp. Biol. 2010, 213, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockwood, B.L.; Somero, G.N. Transcriptomic responses to salinity stress in invasive and native blue mussels (genus Mytilus). Mol. Ecol. 2011, 20, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B. Regulation of hypometabolism: Insights into epigenetic controls. J. Exp. Biol. 2015, 218, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Tomanek, L.; Somero, G.N. Interspecific- and acclimation-induced variation in levels of heat-shock proteins 70 (hsp 70) and 90 (hsp 90) and heat-shock transcription factor-1 (HSF1) in congeneric marine snails (genus Tegula): Implications for regulation of hsp gene expression. J. Exp. Biol. 2002, 205, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Burggren, W. Epigenetic inheritance and its role in evolutionary biology: Re-evaluation and new perspectives. Biology 2016, 5, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapelle, V.; Silvestre, F. Population epigenetics: The extent of DNA methylation variation in wild animal populations. Epigenomes 2022, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Duncan, E.J.; Cunningham, C.B.; Dearden, P.K. Phenotypic plasticity: What has DNA methylation got to do with it? Insects 2022, 13, 110. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hildebrandt, J.-P. Ecology Meets Physiology: Phenotypic Plasticity and the Ability of Animals to Adjust to Changing Environmental Conditions. Physiologia 2023, 3, 366-380. https://doi.org/10.3390/physiologia3020025
Hildebrandt J-P. Ecology Meets Physiology: Phenotypic Plasticity and the Ability of Animals to Adjust to Changing Environmental Conditions. Physiologia. 2023; 3(2):366-380. https://doi.org/10.3390/physiologia3020025
Chicago/Turabian StyleHildebrandt, Jan-Peter. 2023. "Ecology Meets Physiology: Phenotypic Plasticity and the Ability of Animals to Adjust to Changing Environmental Conditions" Physiologia 3, no. 2: 366-380. https://doi.org/10.3390/physiologia3020025