1. Introduction
The cow–calf sector of the beef industry has a disproportional impact on the environmental sustainability of beef production [
1]. Additionally, grazing management, which is most applicable in the cow–calf sector, has the potential to sequester carbon in the soil, offsetting a substantial portion of the greenhouse gas emissions from forage-fed cattle [
2]. Grassland-beef systems are complex, involving the biological processes of soil, plants, and animals, and the impact of management practices are dependent upon characteristics (soil type, forage species, climate, etc.) of the ecosystem [
3]. The future sustainability of beef production depends upon developing a better understanding of the interactions between ecosystem characteristics and management practices, to develop optimal grassland-beef systems specific to the ecosystem.
The multifactorial nature of grassland-beef systems and the long-time horizons for detecting significant changes in soil parameters creates substantial difficulties in designing and performing field experiments to understand the interactions between ecosystem characteristics and management practices. Computer simulation models have been used extensively to evaluate complex biological systems [
4]. Process-based soil and plant computer models have been used to evaluate effects of animal grazing on soil parameters [
5,
6,
7,
8,
9], and to predict the performance of growing and finishing cattle from nutrition and environmental conditions [
10,
11]. A recent dynamic, stochastic cow herd model has been developed to evaluate the sustainability of management decisions over long time horizons [
12,
13], but the current model is inadequate in predicting the growth of suckling beef calves consuming forage.
Extensive research has been performed in the postweaning growing and finishing phases of cattle production systems to determine their feed intake, growth, and chemical body composition [
14,
15,
16,
17,
18,
19,
20,
21], but few studies have evaluated the feed intake and growth of preweaned calves [
22,
23,
24]. The available equations from the literature to predict the forage intake and/or growth of suckling calves are inaccurate and imprecise [
25]; however, this analysis used data accumulated over the 240-day suckling period that was compiled over three decades with calves fed a mixed diet in drylot, which may not accurately represent forage grazing and nutritive value dynamics experienced by suckling calves on pasture. Additionally, most forage intake equations were developed using data from Holstein dairy calves fed in hutches rather than with grazing beef calves. Thus, the objective of this study was to evaluate and develop equations to predict forage intake and growth throughout the suckling period of beef calves grazing on forage or dairy calves fed harvested forage.
4. Discussion
The measurement of forage intake by grazing cattle is fraught with errors regardless of the indirect method used [
44,
45,
46,
47,
48,
49], making the prediction of forage intake difficult. Many factors affect the intake of grazed forage, i.e., selective grazing [
50,
51,
52,
53], sward structure [
50,
54,
55,
56], forage species [
45,
57], and changing plant morphology and nutritive value affects the forage intake during the growing season [
45,
49,
57,
58]. In suckling calves, the animal has multiple choices of nutrients, milk, and forage, further complicating the prediction of forage intake. Numerous studies [
22,
59,
60,
61,
62,
63,
64,
65,
66,
67,
68] have demonstrated that forage intake has a negative relationship with milk intake. Additionally, Webb et al. [
69] demonstrated that calves preferentially consume the available milk supply before consuming forage. The negative relationship of forage intake with milk intake and the preferential consumption of milk allows development of equations to predict forage intake on the assumption that the available milk produced by the dam is consumed first, and subsequently forage intake can be predicted from the negative relationship between milk and forage intakes.
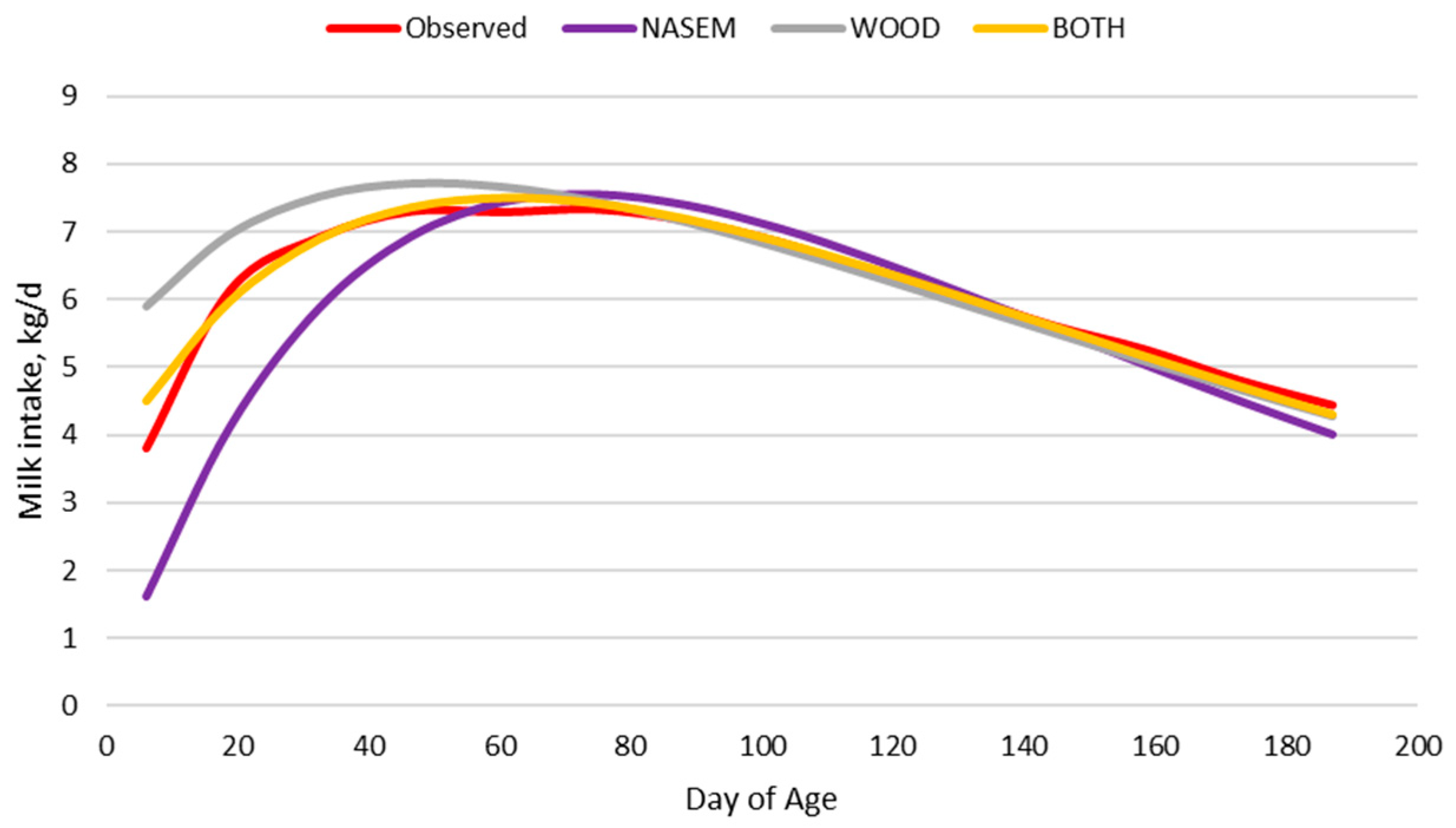
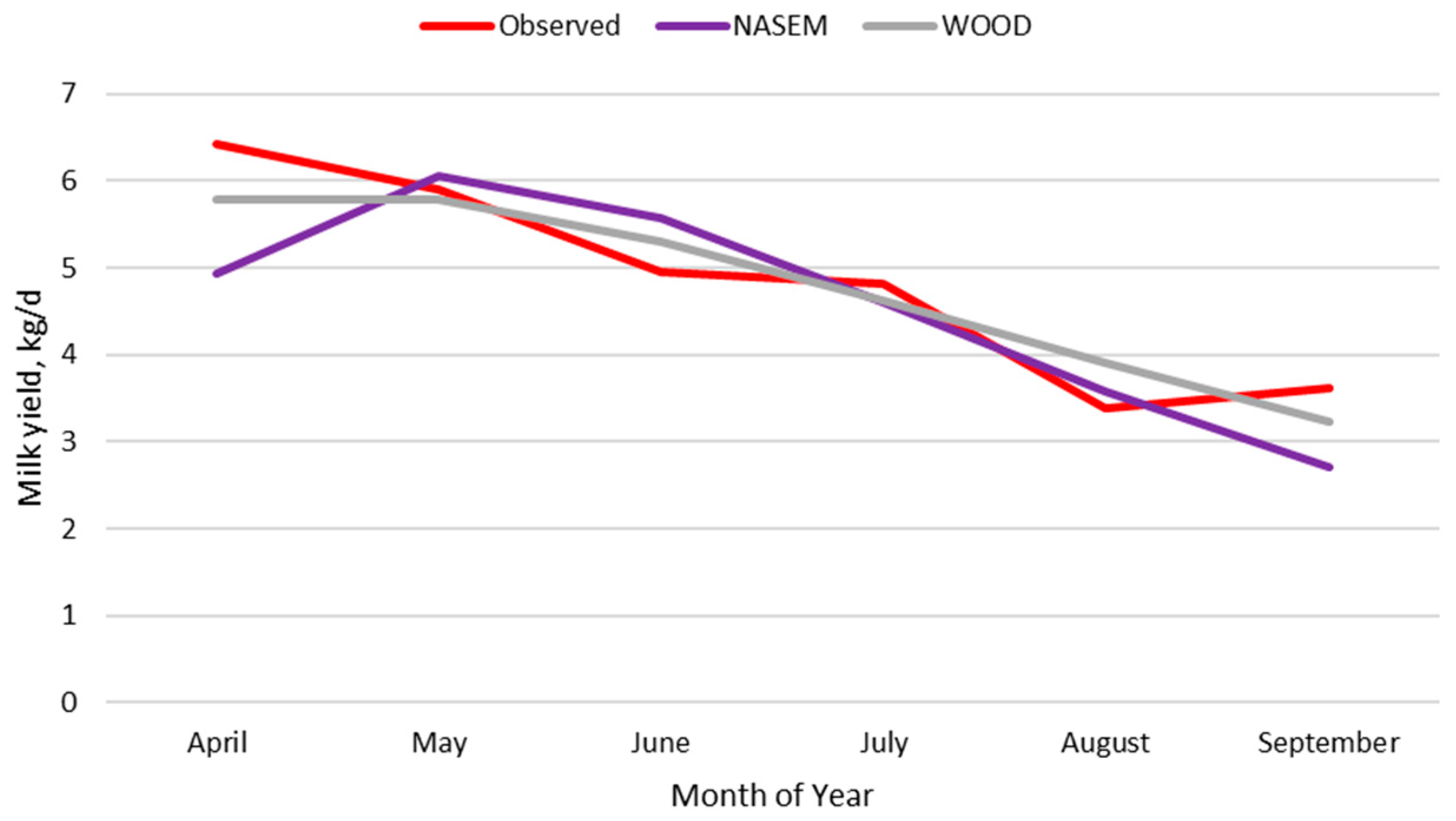
Milk production and milk intake are in dynamic balance. Tedeschi and Fox [
24] reported that calves offered milk at high peak milk curves based on Wood [
26] milk yield equation did not consume all the milk shortly after birth. Dams giving birth to heavier calves produce more milk than those giving birth to lighter calves, possibly due to the greater suckling stimulus and milk intake early in the lactation by heavier calves [
70,
71,
72,
73]. Additionally, milk production is influenced by the nutrient intake of the dam [
74,
75,
76], resulting in varying levels of milk production in the same dams across years, and possibly affecting the shape of the lactation curve [
76,
77]. In dairy cattle, milk composition is also dynamic in that the concentration of components varies with breed, milk yield, and the stage of lactation [
78,
79,
80]; however, a constant energy value of 0.72 Mcal/kg fluid milk is typically used in beef cattle [
10]. Evaluation of the relationship of milk yield and days in milk with milk components indicated that milk composition is not constant in beef cows of various breed types. Breed type influenced the average milk composition but did not interact with milk yield or days in milk such that all breed types followed a similar pattern. In dairy cattle, milk fat and protein are at the greatest concentrations in the first few days after calving, declining steadily until peak milk yield is reached, after which concentrations increase steadily through late lactation such that component yield (milk yield × component concentration) is virtually constant throughout lactation [
78]. The results from the current analysis of milk composition indicate similar relationships of milk yield and days in milk with milk components in beef cows as previously reported in dairy cows. Several studies [
81,
82,
83] have reported increasing concentration of milk components in late lactation of beef cows. Similarly, Peischel [
60] reported that milk protein and digestible energy consumed (kg/d) by suckling beef calves was relatively constant across 7 months of lactation. Increasing energy concentration in late lactation as part of the nutrition model significantly changed the predicted energy intake and growth of calves in late lactation to better align with the observed body weight values.
Throughout the lactation period, milk intake declines but calf body weight increases. Heavier calves will consume more forage and thus the nutrition model must be dynamic to reflect the ever-changing relationship between milk intake, body weight, and forage intake, which complicates the prediction of forage intake and calf growth. The dairy calf intake and body weight dataset with the direct measurements of milk and forage intake and energy concentrations allowed a better evaluation of these relationships, where the model performed reasonably well in predicting forage intake. However, predicting alfalfa hay intake is not the same as predicting grazed forage intake. Forage digestibility influences the rate of fermentation and thus passage rate out of the rumen [
84]; thus, as forage plants mature, the decreasing digestibility results in less forage intake as a percentage of body weight [
10]. Of the forage intake equations evaluated, Eq25, Eq17, and Eq21 adjust for digestibility of the forage; however, these were the least precise and accurate to predict forage intake in the beef calf intake and body weight dataset with changing forage digestibility throughout the grazing season. This could be due to the inaccurate estimates from the in vitro methods of determining forage digestibility used by Boggs [
29]. Most published equations relating in vitro forage dry matter digestibility with in vivo forage dry matter digestibility have an intercept greater than zero and a slope lesser than one [
36,
37,
38,
39,
40,
41], indicating that in vivo forage digestibility is underpredicted at lesser forage digestibility and overpredicted at greater forage digestibility. Forages with lesser digestibility ferment at slower rates, resulting in longer rumen residence times, and forages with greater digestibility ferment at faster rates, resulting in shorter rumen residence times, whereas in vitro methods use a constant rumen fluid incubation time, which does not account for the differing rumen residence times. Bryan [
62] used serial in situ incubation times (12, 24, 36, 48, and 72 h) for forage samples collected from pastures grazed by suckling calves: in vivo forage digestibility throughout the grazing season was best estimated by differing incubation times. The average intercept and slope of the published equations is 8.075 and 0.861, respectively, which is similar to the intercept and slope of the newly developed equation; thus, the new equation may be robust across forage species and maturities.
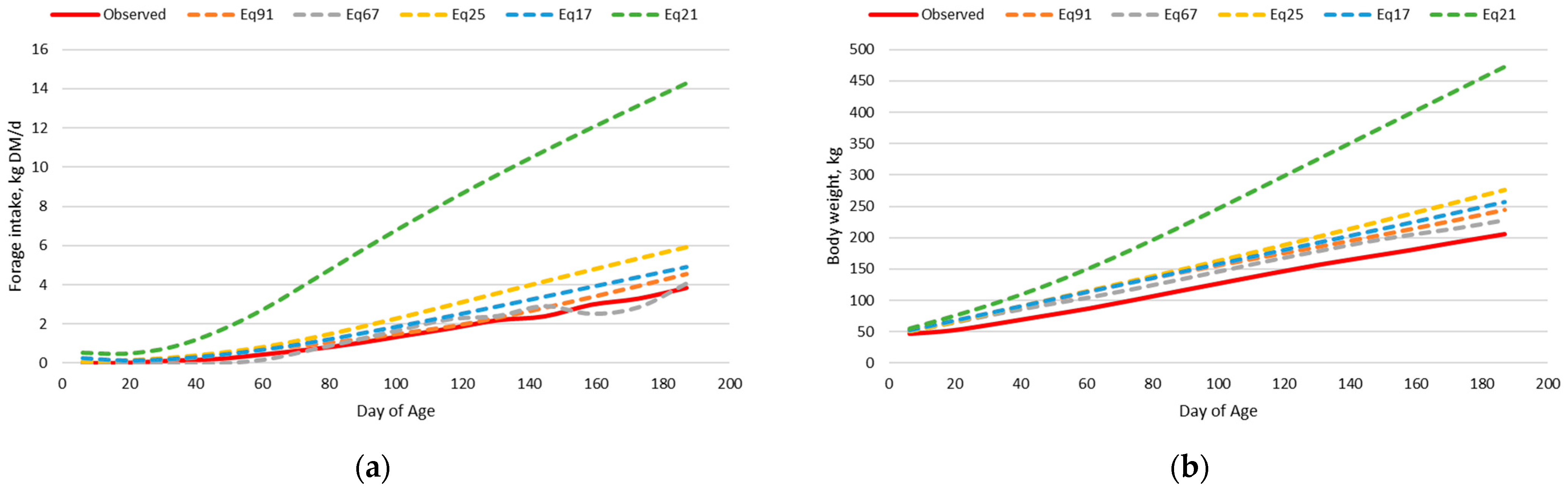
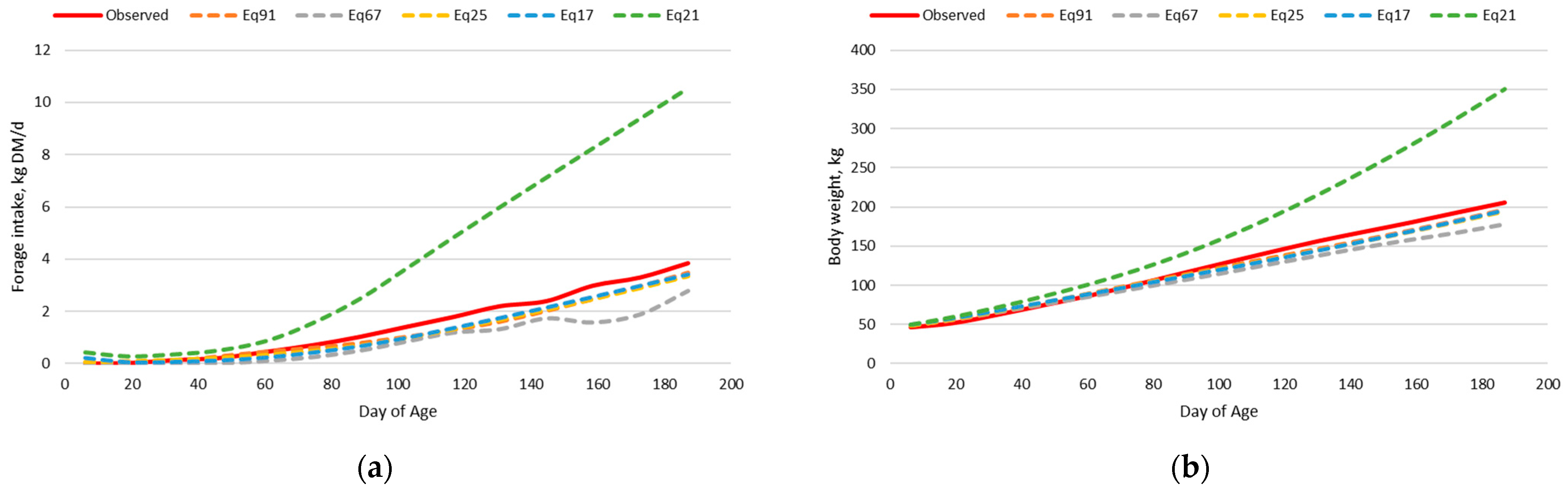
Sequentially dissecting the model into the prediction of forage intake and body weight separately allowed the determination of the aspects leading to imprecision and inaccuracy of the nutrition model. For both the dairy and beef calf intake and body weight datasets, when observed body weight was used in the nutrition model, and thus body weight did not depend upon the prediction from retained energy, the prediction of forage intake was greatly improved. However, when the observed forage intake was used and predicted body weight did depend upon the prediction from retained energy, body weight prediction was imprecise and inaccurate. Thus, indicating that the relationship between retained energy and empty body weight gain may not be the same for very lightweight preweaned calves compared to heavier postweaned cattle. The relationship of retained energy and empty body weight gain is dependent upon the composition of gain [
10,
14,
15], which was the reason for multiple equations based on sex and frame size in the 1984 Nutrient Requirements of Beef Cattle [
85] and the equivalent empty body weight concept in the 2000 Nutrient Requirements of Beef Cattle [
86]. However, these equations were built on data from postweaned cattle. Hildebrand et al. [
87] reported that the relationship between retained energy and empty body weight gain was different between pre- and postweaned cattle and between beef and dairy type cattle, likely due to the different relationships of empty body weight gain with protein and fat gain. Hildebrand et al. [
87] reported that empty body protein gain had a stronger correlation with empty body weight gain than empty body fat gain in preweaning calves (r = 0.971 vs. 0.839), but empty body protein gain had similar correlation as fat gain with empty body weight gain in postweaning calves (r = 0.805 and 0.807). Additionally, the exponent of the equation relating empty body weight gain with retained energy was significantly greater in preweaned versus postweaned cattle and beef versus dairy cattle (0.718, 1.124, 0.356, and 0.762 for preweaned dairy, preweaned beef, postweaned dairy, and postwean beef cattle, respectively). Adjustment of the nutrition model with the new empty body weight gain equations based on pre- and postweaned dairy or beef cattle greatly improved the prediction of body weight in both the dairy and beef calf intake and body weight datasets, indicating that further evaluation of the relationship between empty body weight gain and retained energy from birth to slaughter may be warranted.
The final adjusted beef calf nutrition model had less precision and accuracy than expected; however, further evaluation indicated that calves born earlier relative to the initial milk and forage intake measurements had greater deviation from model predictions. The reason for this is unclear, but not collecting milk intake measurements near birth could have skewed the prediction of milk intake, leading to an inaccurate prediction of forage intake and body weight. Additionally, calves born in March in the Kansas Flint Hills with warm-season native prairie forages may not consume as much forage in the first couple of months after birth, resulting in overprediction by the model. Baker and Barker [
59] reported that forage intake of high nutritive value perennial ryegrass pasture (average in vitro organic matter digestibility = 80.3%) by milk-fed calves was decreased when herbage allowance was less than 40 g/kg LW/day, suggesting that March-born calves in the Kansas Flint Hills may not consume much low-quality forage during the first 2 months of age. Peischel [
60] reported a negative regression coefficient between calf age and grazed Kansas Flint Hills native prairie forage intake in July and August for calves born between March 1 and May 1, indicating that older calves consumed less forage. However, even Eq67, which assumes calves consume no forage in the first 2 months after birth, overpredicted forage intake for calves in birth periods 1 and 2. Bottle-fed dairy calves that consume less starter feed early in life do not have the same extent of rumen development and feed digestion capacity as those that consume more starter feed [
88,
89,
90,
91,
92]. Thus, calves born in early spring may not consume much forage prior to new-growth grass being available, possibly due to the lower nutritive value of hay, and thus may not consume as much forage as expected even at 2 to 3 months of age after lush grass is available. In the dataset from Boggs [
29], calves born in April consumed more forage at 60 days of age in June than March-born calves at 60 days of age in May. Thus, further adjustments to the nutrition model may be needed for calf age relative to the availability of lush grass.
Variation among individual animals in diet digestibility exists [
93,
94,
95,
96]; thus, assuming a constant forage digestibility based on in vitro techniques can result in a significant error (−16 to +25%) in estimating intake [
46]. Additionally, the nutrition model which uses the in vitro forage dry matter digestibility assumes that calves can digest forage equally well in the first couple of months after birth compared with 5 to 6 months after birth. Preston et al. [
97] reported that milk-fed Holstein calves had similar digestibility of a high-quality forage (average dry matter digestibility = 74.6%) at 3 to 6 weeks of age as the same calves at 7 to 10 weeks of age. McCullough and Sisk [
98] reported no difference in alfalfa pellet dry matter digestibility (average = 55%) from 6 to 12 weeks of age in milk-fed calves even though calves younger than 10 weeks of age consumed only small amounts of alfalfa pellets. Godfrey [
99] reported that pasture digestibility (average organic matter digestibility = 75.8%) was similar in 5-, 8-, 11-, and 14-week-old milk-fed dairy calves. Additionally, calves fed an all-milk diet until 8 weeks of age achieved rumen digestive function equal to milk-fed calves allowed to graze pasture since 2 weeks of age in approximately 1 week. Broesder et al. [
63] reported that younger calves (72 days of age) had greater particulate passage rate and shorter rumen retention times, but greater ruminal forage dry matter digestibility than older calves (108 to 151 days of age) when fed alfalfa hay ad libitum. Similarly, Lamothe et al. [
100] reported similar microbial crude protein efficiency in suckling calves from June to September, suggesting that rumen fermentation was equally efficient as in younger suckling calves. Thus, it seems that the capability to digest forage is developed rapidly upon the consumption of forage and that in vitro dry matter digestibility values can be applied to all ages in the nutrition model.
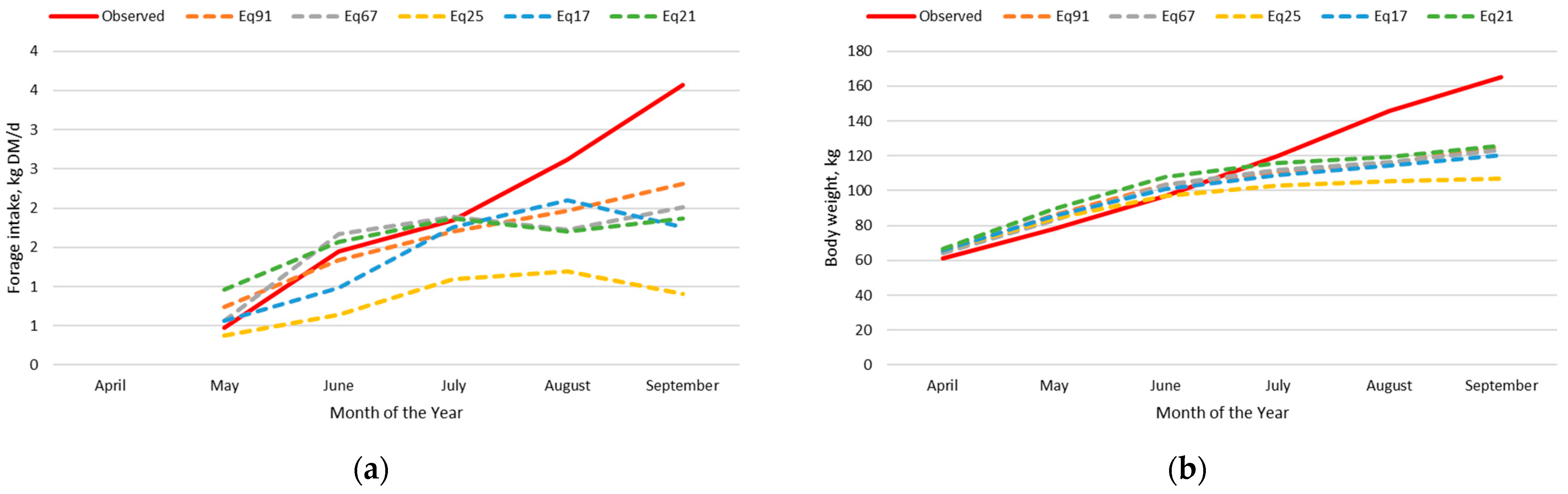
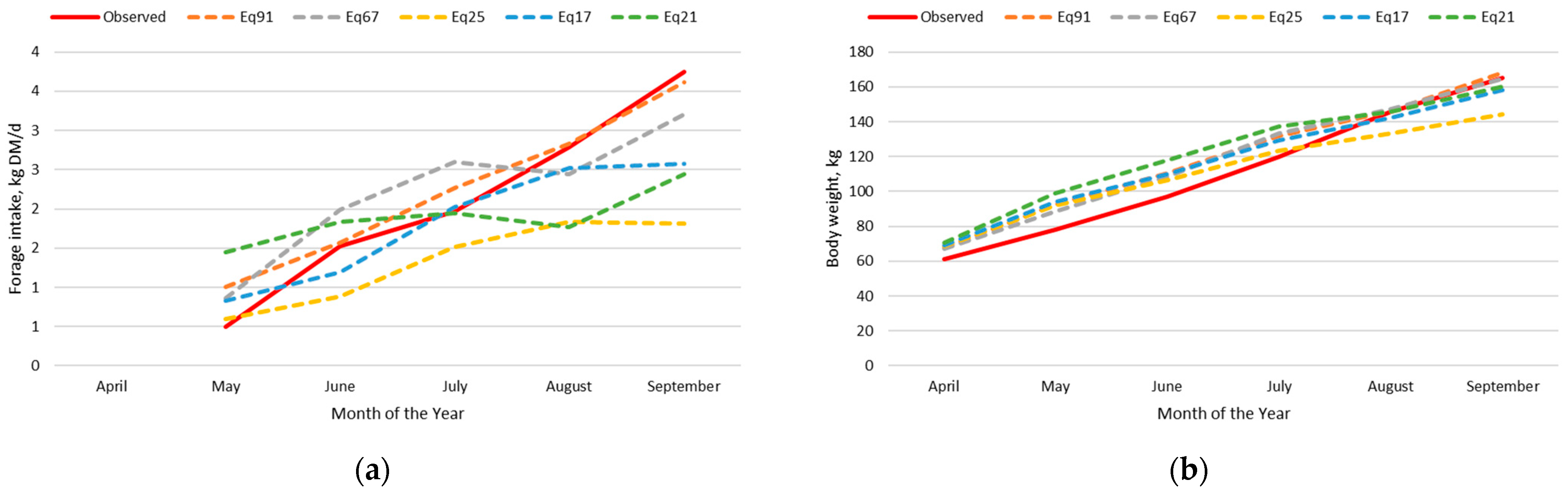
For the dairy calf intake and body weight dataset, Eq91, Eq25, and Eq17 had the greatest precision and accuracy in predicting forage intake and body weight simultaneously in the adjusted nutrition model. This is expected provided that these forage intake equations were developed using this same dataset; however, an adjustment to the empty body weight gain equation for dairy type cattle was necessary to achieve this level of precision and accuracy.
For the beef calf intake and body weight dataset, Eq91 and Eq67 had the best combination of precision and accuracy in predicting forage intake and body weight simultaneously in the adjusted nutrition model. Both equations were developed from bottle fed calves; however, Eq91 was developed from data on Holstein steer calves fed alfalfa hay in confinement, whereas Eq67 was developed with Hereford x Holstein cross steer calves grazing perennial ryegrass pastures through the summer months. The precision of predicting forage intake was lesser with Eq67 than Eq91, but the intercept and the slope for Eq67 were not different than zero and one, respectively, for the prediction of forage intake. Additionally, Eq67 had an intercept not different from zero and a slope near one for the prediction of body weight.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}