

Effect of Short-Term Glycerin Supplementation on Follicle Dynamics and Pregnancy Rate in Goats
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location, Animal, and Experimental Treatments
2.2. Animal Response Measurements
2.2.1. Thermo-Physiological Measurement
2.2.2. Behavioral Evaluation
2.2.3. Peripheral Glycemia and Hematological Parameters
2.3. Ovarian Follicular Dynamics
2.4. Pregnancy Diagnosis and Post-Partum Measurements
2.5. Statistical Analysis
2.6. Ethical Statement
3. Results
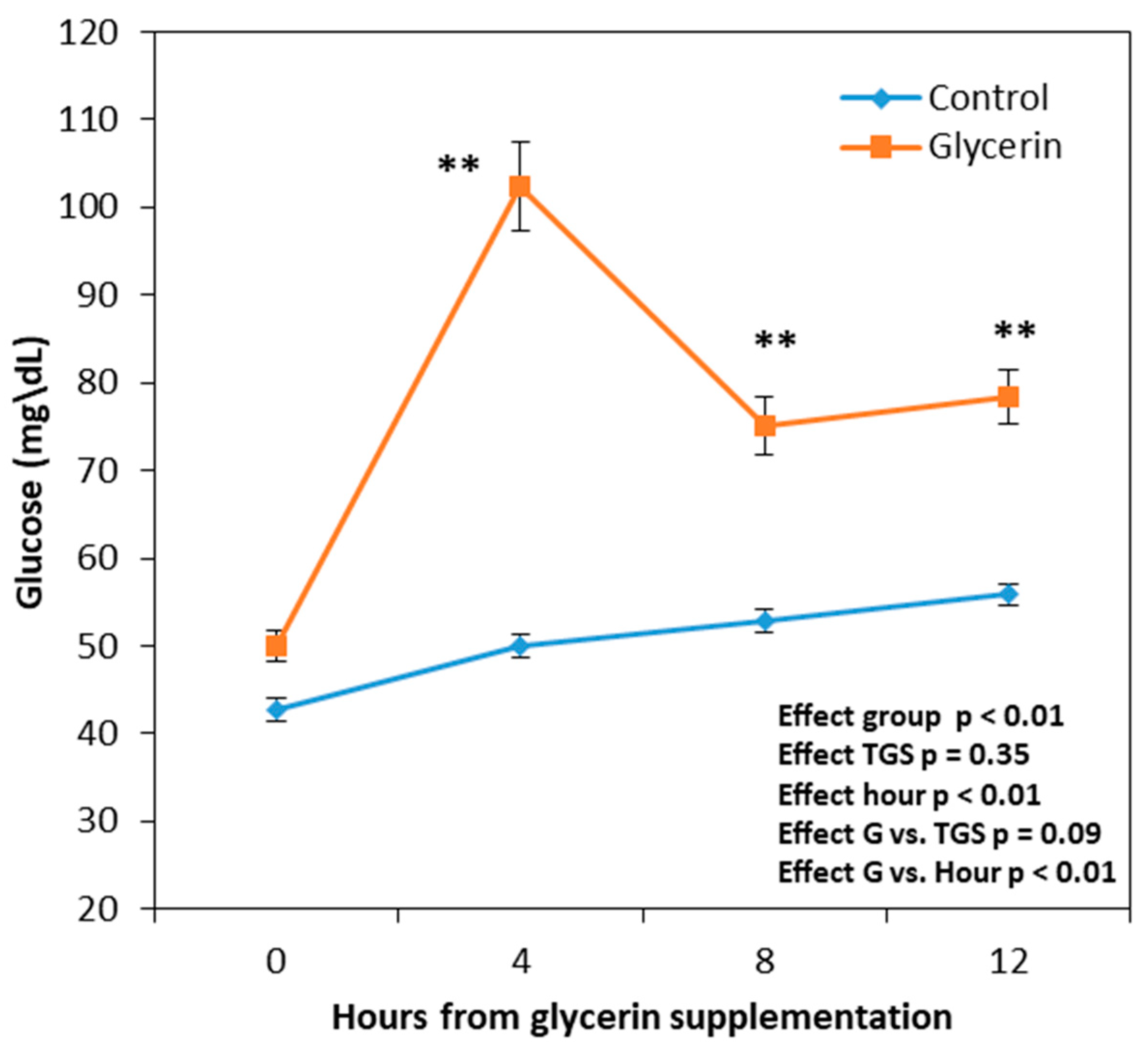
3.1. Peripheral Glucose Levels and Hematological Parameters
3.2. Physiological and Behavior Responses
3.3. Follicle Traits and Reproductive Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Agricultural Production Statistics 2000–2021.FAO. Available online: https://www.fao.org/3/cc3751en/cc3751en.pdf (accessed on 20 July 2023).
- Verruck, S.; Dantas, A.; Prudencio, E.S. Functionality of the components from goat’s milk, recent advances for functional dairy products development and its implications on human health. J. Funct. Foods 2019, 52, 243–257. [Google Scholar] [CrossRef]
- Silva, M.R.L.; Alves, J.P.M.; Fernandes, C.C.L.; Cavalcanti, C.M.; Conde, A.J.H.; Bezerra, A.F.; Soares, A.C.S.; Tetaping, G.M.; De Sá, N.A.R.; Teixeira, D.Í.A.; et al. Use of green microalgae Chlorella as a nutritional supplement to support oocyte and embryo production in goats. Anim. Reprod. Sci. 2023, 256, 107296. [Google Scholar] [CrossRef] [PubMed]
- Syahniar, T.M.; Andriani, M.; Ridla, M.; Laconi, E.B.; Nahrowi, N.; Jayanegara, A. Glycerine as a feed supplement for beef and dairy cattle: A meta-analysis on performance, rumen fermentation, blood metabolites and product characteristics. J. Anim. Physiol. Anim. Nutr. 2020, 105, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Almeida, V.V.S.; Oliveira, A.C.; Silva, R.R.; Ribeiro, J.S.; Oliveira, H.C.; Santos, L.S.; Lima, D.M., Jr. Crude glycerin in the supplementation of crossbred heifers in tropical pastures. Animals 2021, 15, 100088. [Google Scholar] [CrossRef] [PubMed]
- Freire, L.F.D.S.; Cruz, G.R.B.; Costa, R.G.; Ribeiro, N.L.; Guerra, R.R.; Sousa, S.; Nascimento, G.V.D.; Lima, A.M.; Silva, G.F.; Sant’ana, A.M.S.; et al. Glycerin diet affects the size of the fat globule and the fatty acid profile of goat’s milk. Food Sci. Technol. 2022, 42, e85821. [Google Scholar] [CrossRef]
- Merlim, F.A.; Silva Sobrinho, A.G.; Borghi, T.H.; Cirne, L.G.A.; Valença, R.L.; Almeida, F.A.; Viviane Endo, V.; Viegas, C.R.; Zeola, N.M.B.L. Crude glycerin is an efficient alternative to corn in the diet of feedlot lambs. Arch. Anim. Breed. 2021, 64, 387–393. [Google Scholar] [CrossRef]
- Gutierrez, C.G.; Ferraro, S.; Martinez, V.; Saharrea, A.; Cortez, C.; Lassala, A.; Basurto, H.; Hernandez, J. Increasing ovulation quota: More than a matter of energy. Acta Sci. Vet. 2011, 39, s305–s316. [Google Scholar]
- Rodrigues, F.V.; Silva, C.M.G.; Lima, I.M.T.; Silva, A.M.; Fernandes, C.C.L.; Rondina, D. Effect of oral drenching of glycerin as a source of pre-mating energetic supplementation on reproductive response in goats. Anim. Reprod. 2015, 12, 890–898. [Google Scholar]
- Silva, L.M.; Oliveira, C.H.A.; Silva, A.M.; Fernandes, C.C.L.; Duarte, S.S.; Lima, I.M.T.; Silva, C.M.G.; Martins, L.T.; Calderón, C.E.M.; Tavares, K.C.S.; et al. Gene expression and embryo quality in superovulated goats supplemented with crude glycerin after mating. Small Rumin. Res. 2014, 120, 71–77. [Google Scholar] [CrossRef]
- Kabir, M.E.; Miraz, F.H.; Alam, M.H.; Sarker, M.B.; Hashem, M.A.; Khandoker, M.Y.; Moniruzzaman, M. Dietary energy influences ovarian morphology and in vitro maturation of oocytes in goats. J. Appl. Anim. Res. 2022, 50, 47–53. [Google Scholar] [CrossRef]
- Childs, G.V.; Odle, A.K.; MacNicol, M.C.; MacNicol, A.M. The importance of leptin to reproduction. Endocrinology 2021, 162, bqaa204. [Google Scholar] [CrossRef] [PubMed]
- Bach, À. Effects of nutrition and genetics on fertility in dairy cows. Reprod. Fertil. Dev. 2019, 31, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Chaput, C.; Sirard, M.A. Embryonic response to high beta-hydroxybutyrate (BHB) levels in postpartum dairy cows. Domest. Anim. Endocrinol. 2020, 72, 106431. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, H.; Santos, E.; Oliveira, J.; Carvalho, G.; Silva, F.; Cassuce, M.; Perazzo, A.; Zanini, A.; Pinho, R. Performance and ruminal parameters of boer crossbred goats fed diets that contain crude glycerin. Animals 2019, 9, 967. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.M.; Cruz, G.R.B.; Costa, R.G.; Ribeiro, N.L.; Beltrão Filho, E.M.; Sousa, S.; Justino, E.S.; Santos, D.G. Physical-chemical and microbiological quality of milk and cheese of goats fed with bidestilated glycerin. Food Sci. Technol. 2021, 41, 25–33. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Small Ruminants; National Academy Press: Washington, DC, USA, 2007. [Google Scholar]
- Viñoles, C.; Paganoni, B.; Glover, K.M.M.; Milton, J.T.B.; Blache, D.; Blackberry, M.A.; Martin, G.B. The use of a’first-wave’model to study the effect of nutrition on ovarian follicular dynamics and ovulation rate in the sheep. Reproduction 2010, 140, 865. [Google Scholar] [CrossRef] [PubMed]
- Mach, N.; Bach, A.; Devant, M. Effects of crude glycerin supplementation on performance and meat quality of Holstein bulls fed high-concentrate diets. J. Anim. Sci. 2009, 87, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.C.L.; Aguiar, L.H.; Calder’on, C.E.M.; Silva, A.M.; Alves, J.P.M.; Rossetto, R.; Bertolini, L.R.; Bertolini, M.; Rondina, D. Nutritional impact on gene expression and competence of oocytes used to support embryo development and livebirth by cloning procedures in goats. Anim. Reprod. Sci. 2018, 188, 1–12. [Google Scholar] [CrossRef]
- Bateson, M.; Martin, P. Measuring Behaviour: An Introductory Guide, 3rd ed.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Fernandes, C.C.L.; Feltrin, C.; Martins, L.T.; Gaudêncio Neto, S.; Aguiar, L.H.; Silva, A.M.; Oliveira, C.H.A.; Silva, L.M.; Silva, C.M.G.; Bertolini, M.; et al. Goat oocyte quality and competence to undergo IVM and embryo development after parthenogenetic activation from goats fed with different levels of cashew nut bran as source of dietary lipids. Theriogenology 2014, 82, 332–337. [Google Scholar] [CrossRef]
- Ginther, O.J.; Kot, K. Follicular dynamics during the ovulatory season in goats. Theriogenology 1994, 42, 987–1001. [Google Scholar] [CrossRef]
- Quandt, J.E.; Barletta, M.; Cornell, K.K.; Giguère, S.; Hofmeister, E.H. Evaluation of a point-of-care blood glucose monitor in healthy goats. J. Vet. Emerg. Crit. Care 2018, 28, 45–53. [Google Scholar] [CrossRef]
- Chadt, A.; Al-Hasani, H. Glucose transporters in adipose tissue, liver, and skeletal muscle in metabolic health and disease. Pflugers Arch. Eur. J. Physiol. 2020, 472, 1273–1298. [Google Scholar] [CrossRef]
- Kalyesubula, M.; Rosov, A.; Alon, T.; Moallem, U.; Dvir, H. Intravenous infusions of glycerol versus propylene glycol for the regulation of negative energy balance in sheep: A randomized trial. Animals 2019, 9, 731. [Google Scholar] [CrossRef]
- Ferraro, S.M.; Mendoza, G.D.; Miranda, L.A.; Gutiérrez, C.G. In vitro ruminal fermentation of glycerol, propylene glycol and molasses combined with forages and their effect on glucose and insulin blood plasma concentrations after an oral drench in sheep. Anim. Feed. Sci. Technol. 2016, 213, 74–80. [Google Scholar] [CrossRef]
- James, D.E.; Stöckli, J.; Birnbaum, M.J. The aetiology and molecular landscape of insulin resistance. Nat. Rev. Mol. Cell Biol. 2021, 22, 751–771. [Google Scholar] [CrossRef]
- Porcu, C.; Manca, C.; Cabiddu, A.; Dattena, M.; Gallus, M.; Pasciu, V.; Succu, S.; Naitana, S.; Berlinguer, F.; Molle, G. Effects of short-term administration of a glucogenic mixture at mating on feed intake, metabolism, milk yield and reproductive performance of lactating dairy ewes. Anim. Feed. Sci. Technol. 2018, 243, 10–21. [Google Scholar] [CrossRef]
- van Cleef, E.H.C.B.; Almeida, M.T.C.; Perez, H.L.; Paschoaloto, J.R.; Castro Filho, E.S.; Ezequiel, J.M.B. Effects of partial or total replacement of corn cracked grain with high concentrations of crude glycerin on rumen metabolism of crossbred sheep. Small Rumin. Res. 2018, 159, 45–51. [Google Scholar] [CrossRef]
- Sharma, K.; Akre, S.; Chakole, S.; Wanjari, M.B. Stress-Induced Diabetes: A Review. Cureus 2022, 14, e29142. [Google Scholar] [CrossRef]
- Hillis, G.S.; Woodward, M.; Rodgers, A.; Chow, C.K.; Li, Q.; Zoungas, S.; Patel, A.; Webster, R.; Batty, G.D.; Ninomiya, T.; et al. Resting heart rate and the risk of death and cardiovascular complications in patients with type 2 diabetes mellitus. Diabetologia 2012, 55, 1283–1290. [Google Scholar] [CrossRef]
- Shah, A.; Wondisford, F.E. Gluconeogenesis Flux in Metabolic Disease. Annu. Rev. Nutr. 2023, 43, 153–177. [Google Scholar] [CrossRef]
- Andrade, M.A.M.M.; Alves, J.P.M.; Galvão, I.T.O.M.; Cavalcanti, C.M.; Silva, M.R.L.; Conde, A.J.H.; Bezerra, A.F.; Fernandes, C.C.L.; Teixeira, D.I.A.; Rondina, D. Glycerin supplementation strategies for three or seven days affects oxidative stress, follicle dynamics and ovulatory response in Morada Nova sheep. Anim. Reprod. 2022, 19, e20200025. [Google Scholar] [CrossRef]
- Oliveira, F.B.B.; da Silva, C.P.; Fernandes, C.C.L.; de Sousa, F.C.; Silva, A.M.; Pinheiro, D.C.N.; Rodrigues, L.F.S.; Rondina, D. Ovarian follicular growth in Morada Nova sheep supplemented with crude glycerin before mating. Rev. Acad. Ciênc. Anim. 2016, 14, 235. [Google Scholar] [CrossRef]
- Alves, J.P.M.; Fernandes, C.C.L.; Rossetto, R.; da Silva, C.P.; Galvão, I.T.O.M.; Bertolini, M.; Rondina, D. Impact of short nutrient stimuli with different energy source on follicle dynamics and quality of oocyte from hormonally stimulated goats. Reprod. Domest. Anim. 2019, 54, 1206–1216. [Google Scholar] [CrossRef]
- Guo, Y.X.; Duan, C.H.; Hao, Q.H.; Liu, Y.Q.; Li, T.; Zhang, Y.J. Effect of short-term nutritional supplementation on hormone concentrations in ovarian follicular fluid and steroid regulating gene mRNA abundances in granulosa cells of ewes. Anim. Reprod. Sci. 2019, 211, 106208. [Google Scholar] [CrossRef]
- Nogueira, D.M.; Eshtaeba, A.; Cavalieri, J.; Fitzpatrick, L.A.; Gummow, B.; Blache, D.; Parker, A.J. Short-term supplementation with maize increases ovulation rate in goats when dietary metabolizable energy provides requirements for both maintenance and 1.5 times maintenance. Theriogenology 2017, 89, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Habibizad, J.; Riasi, A.; Kohram, H.; Rahmani, H.R. Effect of long-term or short-term supplementation of high energy or high energy-protein diets on ovarian follicles and blood metabolites and hormones in ewes. Small Rumin. Res. 2015, 132, 37–43. [Google Scholar] [CrossRef]
- Fernandez, J.; Bruno-Galarraga, M.M.; Soto, A.T.; de la Sota, R.L.; Cueto, M.I.; Lacau, I.M.; Gibbons, A.E. Hormonal therapeutic strategy on the induction of accessory corpora lutea in relation to follicle size and on the increase of progesterone in sheep. Theriogenology 2018, 105, 184–188. [Google Scholar] [CrossRef]
- Van Eetvelde, M.; Heras, S.; Leroy, J.L.M.R.; Van Soom, A.; Opsomer, G. The importance of the periconception period: Immediate effects in cattle breeding and in assisted reproduction such as artificial insemination and embryo transfer. Periconception Physiol. Med. 2017, 1014, 41–68. [Google Scholar]
- Uhde, K.; van Tol, H.T.; Stout, T.A.; Roelen, B.A. Exposure to elevated glucose concentrations alters the metabolomic profile of bovine blastocysts. PLoS ONE 2018, 13, e0199310. [Google Scholar] [CrossRef]
- Dupont, J.; Scaramuzzi, R.J. Insulin signalling and glucose transport in the ovary and ovarian function during the ovarian cycle. Biochem. J. 2016, 473, 1483–1501. [Google Scholar] [CrossRef]
- Adamiak, S.J.; Powell, K.; Rooke, J.A.; Webb, R.; Sinclair, K.D. Body composition, dietary carbohydrates and fatty acids determine post-fertilisation development of bovine oocytes in vitro. Reproduction 2006, 131, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, D.; Båge, R.; Humblot, P.; Andersson, G.; Sirard, M.A.; Sjunnesson, Y. Insulin during in vitro oocyte maturation has an impact on development, mitochondria, and cytoskeleton in bovine day 8 blastocysts. Theriogenology 2017, 101, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Ciani, F.; Cocchia, N.; d’Angelo, D.; Tafuri, S. Influence of ROS on ovarian functions. New Discov. Embryol. 2015, 21, 41–73. [Google Scholar] [CrossRef]
- Senosy, W.; Mahmoud, G.B.; Abdel-Raheem, S.M. Influence of short-term energy supplementation on estrus, ovarian activity, and blood biochemistry in Ossimi ewes synchronized with fluorogestone acetate in the subtropics. Theriogenology 2017, 88, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Scaramuzzi, R.J.; Zouaïdi, N.; Menassol, J.B.; Dupont, J. The effects of intravenous, glucose versus saline on ovarian follicles and their levels of some mediators of insulin signalling. Reprod. Biol. Endocrinol. 2015, 13, 6. [Google Scholar] [CrossRef]
- Sánchez, F.; Romero, S.; Smitz, J. Oocyte and cumulus cell transcripts from cultured mouse follicles are induced to deviate from normal in vivo conditions by combinations of insulin, follicle-stimulating hormone, and human chorionic gonadotropin. Biol. Reprod. 2011, 85, 565–574. [Google Scholar] [CrossRef]
- Viñoles, C.; Forsberg, M.; Martin, G.B.; Cajarville, C.; Repetto, J.; Meikle, A. Short-term nutritional supplementation of ewes in low body condition affects follicle development due to an increase in glucose and metabolic hormones. Reproduction 2005, 129, 299–309. [Google Scholar] [CrossRef]
- Stewart, R.; Oldham, C.M. Feeding lupins for 4 days during the luteal phase can increase ovulation rate. Proc. Aust. Soc. Anim. Prod. 1986, 16, 367. [Google Scholar]
- Muñoz-Gutiérrez, M.; Blache, D.; Martin, G.B.; Scaramuzzi, R.J. Folliculogenesis and ovarian expression of mRNA encoding aromatase in anoestrous sheep after 5 days of glucose or glucosamine infusion or supplementary lupin feeding. Reproduction 2002, 124, 721–731. [Google Scholar] [CrossRef]
- Viñoles, G.C. Effect of Nutrition of Follicle Development and Ovulation Rate in the Ewe. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2003. [Google Scholar]


{kind=link}
| Parameters | Group | p Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | Glycerin | SEM | Group | TGS | Hour | G × TGS | G × Hour | |
| Hematological response * | ||||||||
| WBC, ×103/µL | 11.6 | 11.8 | 0.33 | 0.22 | 0.94 | - | 0.88 | - |
| RBC, ×106/µL | 13.4 | 14.2 | 0.53 | 0.84 | 0.52 | - | 0.67 | - |
| Physiological response | ||||||||
| HR, beat\min | 66.4 | 90.6 | 0.85 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| RF, breaths\min | 32.2 | 37.2 | 0.71 | <0.01 | <0.01 | <0.01 | 0.04 | 0.74 |
| ST, °C | 35.5 | 35.3 | 0.07 | 0.13 | <0.01 | <0.01 | 0.20 | 0.06 |
| RT, °C | 38.8 | 38.9 | 0.02 | <0.01 | 0.22 | <0.01 | <0.01 | <0.01 |
| Behavior response | ||||||||
| Rumination, n | 1.3 | 0.4 | 0.05 | <0.01 | <0.01 | <0.01 | 0.07 | <0.01 |
| Idle, n | 1.7 | 2.6 | 0.09 | <0.01 | 0.64 | <0.01 | 0.20 | 0.38 |
| Drinking, n | 0.00 | 0.3 | 0.02 | <0.01 | 0.05 | <0.01 | 0.05 | <0.01 |
| Feeding, n | 2.9 | 2.6 | 0.11 | 0.60 | <0.01 | <0.01 | 0.57 | <0.01 |
| Urination, n | 0.1 | 0.3 | 0.01 | <0.01 | 0.33 | 0.61 | 0.31 | 0.88 |
| Defecation, n | 0.1 | 0.1 | 0.01 | 0.13 | 0.42 | 0.04 | 0.01 | 0.36 |
| Stress indicators, n | 2.8 | 3.1 | 0.19 | 0.02 | <0.01 | <0.01 | 0.71 | <0.01 |
| Parameters | Group | p Value | ||||
|---|---|---|---|---|---|---|
| Control | Glycerin | SEM | Group | TGS | G × TGS | |
| Follicle traits before ovulation induction * | ||||||
| Follicles > 4 mm, n\ovary | 0.01 | 0.06 | 0.01 | 0.04 | 0.38 | 0.04 |
| Total follicles, n\ovary | 3.4 | 3.4 | 0.06 | 0.53 | 0.51 | 0.63 |
| Follicle diameter, mm | 3.4 | 3.7 | 0.05 | <0.01 | 0.03 | 0.78 |
| Follicle traits after ovulation induction ** | ||||||
| Follicles > 4 mm, n\ovary | 0.08 | 0.2 | 0.01 | 0.01 | 0.04 | 0.61 |
| Total follicles, n\ovary | 3.4 | 3.5 | 0.05 | 0.31 | 0.69 | 0.79 |
| Follicle diameter, mm | 3.9 | 4.2 | 0.04 | <0.01 | <0.01 | 0.92 |
| Reproductive response | ||||||
| Pregnancy rate, % (n/n) | 66.7 (8/12) | 76.9 (10/13) | - | 0.57 | - | - |
| Twinning rate, % (n/n) | 25.0 (2/8) | 30.0 (3/10) | - | 0.74 | - | - |
| Kidding rate, % (n/n) | 62.5 (5/8) | 80.0 (8/10) | - | 0.27 | - | - |
| Litter size | 1.5 | 1.4 | 0.15 | 0.84 | - | - |
| Kid weight at partum/doe, kg | 4.9 | 5.2 | 0.56 | 0.86 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, C.P.; Fernandes, C.C.L.; Alves, J.P.M.; de Oliveira, F.B.B.; Silva, A.M.; de Souza, F.C.; Cavalcante, C.M.; Conde, A.J.F.; do Rego, A.C.; Rondina, D. Effect of Short-Term Glycerin Supplementation on Follicle Dynamics and Pregnancy Rate in Goats. Ruminants 2023, 3, 445-456. https://doi.org/10.3390/ruminants3040036
da Silva CP, Fernandes CCL, Alves JPM, de Oliveira FBB, Silva AM, de Souza FC, Cavalcante CM, Conde AJF, do Rego AC, Rondina D. Effect of Short-Term Glycerin Supplementation on Follicle Dynamics and Pregnancy Rate in Goats. Ruminants. 2023; 3(4):445-456. https://doi.org/10.3390/ruminants3040036
Chicago/Turabian Styleda Silva, Caroline Pessoa, César Carneiro Linhares Fernandes, Juliana Paula Martins Alves, Felipe Brener Bezerra de Oliveira, Aline Maia Silva, Francisco Carlos de Souza, Camila Muniz Cavalcante, Alfredo José Ferreira Conde, Anibal Coutinho do Rego, and Davide Rondina. 2023. "Effect of Short-Term Glycerin Supplementation on Follicle Dynamics and Pregnancy Rate in Goats" Ruminants 3, no. 4: 445-456. https://doi.org/10.3390/ruminants3040036