

Effect of Tannin Inclusion on the Enhancement of Rumen Undegradable Protein of Different Protein Sources
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location and Ethical Approval
2.2. Experimental Designs and Chemical Analysis
2.3. In Situ Procedures and Calculations
2.4. Intestinal Digestibility Procedures and Calculations
2.5. Statistical Analysis
3. Results
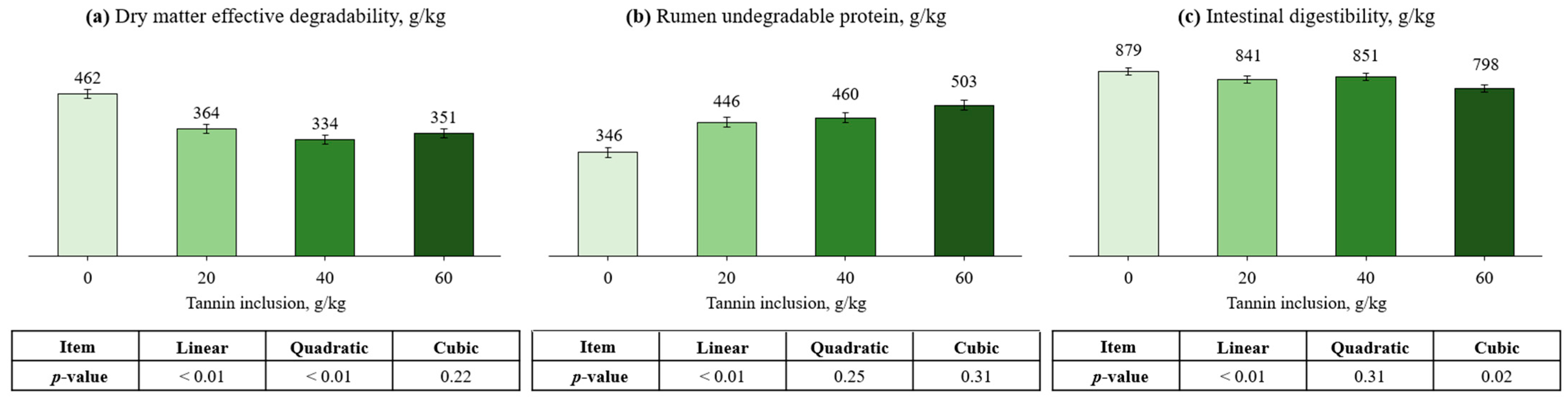
3.1. Cottonseed Meal
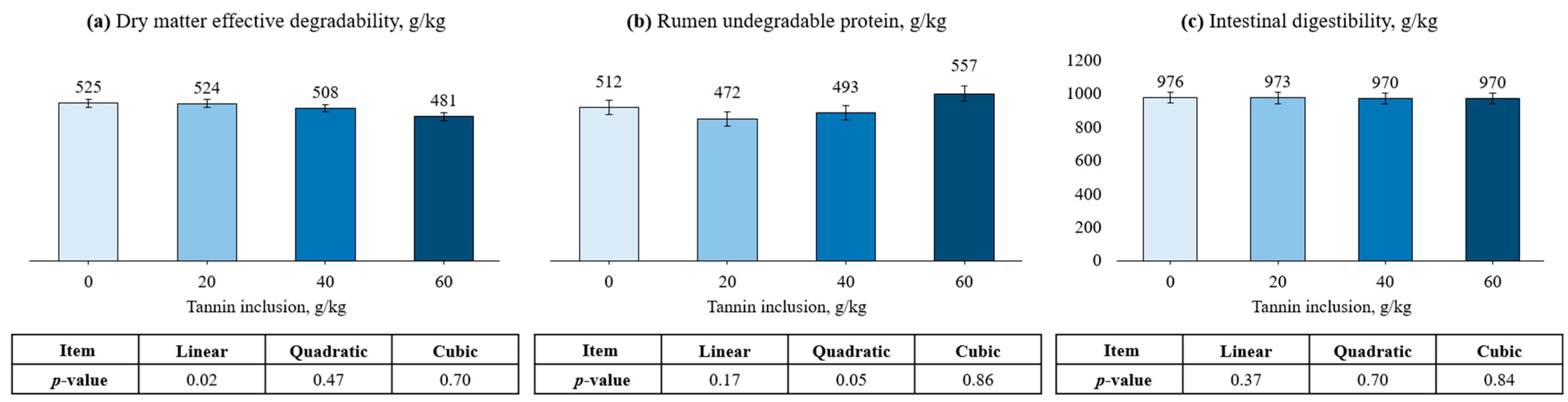
3.2. Peanut Meal
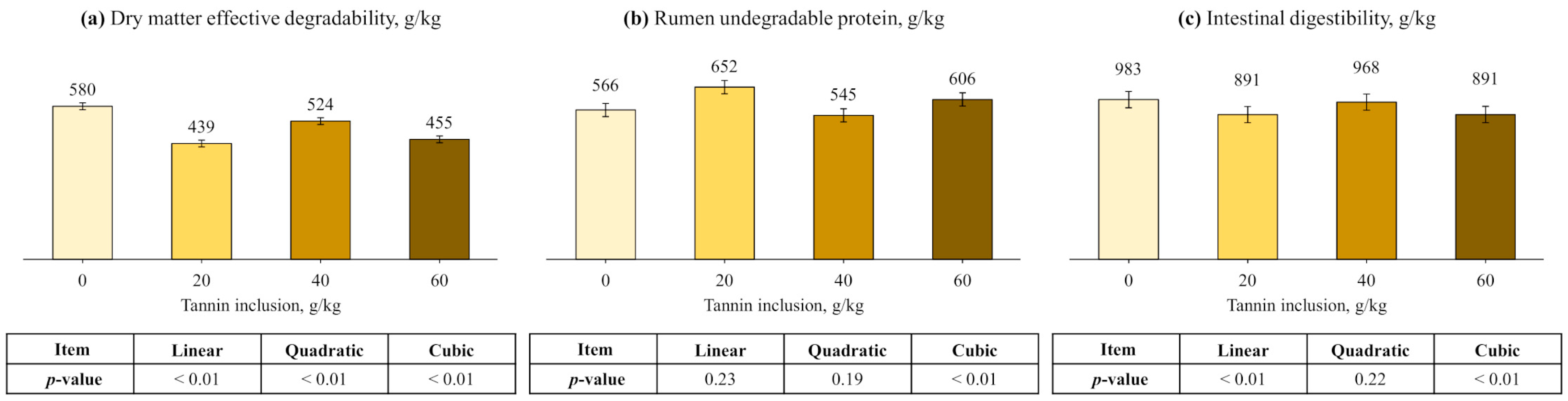
3.3. Soybean Meal
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albino, F.T. Frango de Corte—Manual Prático de Manejo e Produção; Aprenda Fácil: Viçosa, MG, Brasil, 1998; p. 72. [Google Scholar]
- Pires, A.V. Bovinocultura de Corte—Volume I; FEALQ: Piracicaba, SP, Brasil, 2010; Volume 1, p. 1760. ISBN 978-85-7133-069-6. [Google Scholar]
- Rostagno, H.; Albino, L.F.T.; Donzele, J.P.; Gomes, P.C.; Oliveira, R.F.D.; Lopes, D.C.; Ferreira, A.S.; Barreto, S.L.d.T.; Euclides, R.F. Tabelas Brasileiras Para Aves e Suínos, 3rd ed.; UFV: Viçosa, MG, Brasil, 2011; p. 252. ISBN 978-85-6024-97-2-5. [Google Scholar]
- Silva, F.A.S.; Benedeti, P.D.B.; Silva, L.F.C.E.; Rotta, P.P.; Menezes, A.C.B.; Marcondes, M.I.; Valadares Filho, S.C. Protein and Amino Acids Requirements for Beef Cattle. In Nutrient Requirements of Zebu and Crossbred Cattle, 4th ed.; Valadares Filho, S.C., Saraiva, D.T., Benedeti, P.D.B., Silva, F.A.S., Chizzotti, M.L., Eds.; Independent Production: Visconde de Rio Branco, MG, Brazil, 2023; pp. 201–232. [Google Scholar] [CrossRef]
- Yousefinejad, S.; Fattahnia, F.; Kazemi-Bonchenari, M.; Nobari, B.; Ghaffari, M.H. Effects of Protein Content and Rumen-Undegradable to Rumen-Degradable Protein Ratio in Finely Ground Calf Starters on Growth Performance, Ruminal and Blood Parameters, and Urinary Purine Derivatives. J. Dairy Sci. 2021, 104, 8798–8813. [Google Scholar] [CrossRef] [PubMed]
- National Academies of Sciences, Engineering, and Medicine, Nutrient Requirements of Beef Cattle: Eighth Revised Edition; National Academy Press: Washington, DC, USA, 2016.
- Pinto, A.C.J.; Millen, D.D. Nutritional Recommendations and Management Practices Adopted by Feedlot Cattle Nutritionists: The 2016 Brazilian Survey. Can. J. Anim. Sci. 2019, 99, 392–407. [Google Scholar] [CrossRef]
- Molosse, V.L.; Pereira, D.A.B.; Rigon, F.; Loregian, K.E.; Magnani, E.; Marcondes, M.I.; Branco, R.H.; Benedeti, P.D.B.; Paula, E.M. Use of Heating Methods and Xylose to Increase Rumen Undegradable Protein of Alternative Protein Sources: (2) Cottonseed Meal. Animals 2022, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Rigon, F.; Pereira, D.A.B.; Loregian, K.E.; Magnani, E.; Marcondes, M.I.; Branco, R.H.; Benedeti, P.D.B.; Paula, E.M. Use of Heating Methods and Xylose to Increase Rumen Undegradable Protein of Alternative Protein Sources: (1) Peanut Meal. Animals 2022, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Brand, T.S.; Jordaan, L.; Dreyer, O. Effect of Extrusion with Molasses on the Rumen Undegradable Protein Fraction of Canola Oilcake Meal and Sweet Lupins. S. Afr. J. Anim. Sci. 2022, 52, 792–801. [Google Scholar] [CrossRef]
- Samadi; Yu, P. Dry and Moist Heating-Induced Changes in Protein Molecular Structure, Protein Subfraction, and Nutrient Profiles in Soybeans. J. Dairy Sci. 2011, 94, 6092–6102. [Google Scholar] [CrossRef] [PubMed]
- Shishir, M.S.R.; Brodie, G.; Cullen, B.; Kaur, R.; Cho, E.; Cheng, L. Microwave Heat Treatment Induced Changes in Forage Hay Digestibility and Cell Microstructure. Appl. Sci. 2020, 10, 8017. [Google Scholar] [CrossRef]
- Paya, H.; Taghizadeh, A.; Janmohammadi, H.; Moghaddam, G.A.; Khani, A.H.; Alijani, S. Effects of microwave irradiation on in vitro ruminal fermentation and ruminal and post-ruminal disappearance of safflower seed. J. Biodivers. Environ. Sci. 2014, 5, 349–356. [Google Scholar]
- Akbarian, A.; Khorvash, M.; Ghorbani, G.R.; Ghasemi, E.; Dehghan-Banadaky, M.; Shawrang, P.; Hosseini Ghaffari, M. Effects of Roasting and Electron Beam Irradiating on Protein Characteristics, Ruminal Degradability and Intestinal Digestibility of Soybean and the Performance of Dairy Cows. Livest. Sci. 2014, 168, 45–52. [Google Scholar] [CrossRef]
- Mojarrad, M.T.; Seidavi, A.; Dadashbeiki, M.; Roca-Fernández, A.I. Short communication. Effect of soybean meal heat procedures on growth performance of broiler chickens. Span. J. Agric. Res. 2014, 12, 180–185. [Google Scholar] [CrossRef]
- Vanegas, J.L.; González, J.; Carro, M.D. Influence of Protein Fermentation and Carbohydrate Source on In Vitro Methane Production. J. Anim. Physiol. Anim. Nutr. 2017, 101, e288–e296. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.A.; Cheng, K.-J.; Beauchemin, K.A.; Bailey, D.R.C.; Pickard, M.D.; Gilbert, R.P. Use of Lignosulfonate to Decrease the Rumen Degradability of Canola Meal Protein. Can. J. Anim. Sci. 1993, 73, 211–215. [Google Scholar] [CrossRef]
- Lima, P.R.; Apdini, T.; Freire, A.S.; Santana, A.S.; Moura, L.M.L.; Nascimento, J.C.S.; Rodrigues, R.T.S.; Dijkstra, J.; Garcez Neto, A.F.; Queiroz, M.A.Á.; et al. Dietary Supplementation with Tannin and Soybean Oil on Intake, Digestibility, Feeding Behavior, Ruminal Protozoa and Methane Emission in Sheep. Anim. Feed Sci. Technol 2019, 249, 10–17. [Google Scholar] [CrossRef]
- Roca-Fernández, A.I.; Dillard, S.L.; Soder, K.J. Ruminal fermentation and enteric methane production of legumes containing condensed tannins fed in continuous culture. J. Dairy Sci. 2020, 103, 7028–7038. [Google Scholar] [CrossRef] [PubMed]
- Mezzomo, R.; Paulino, P.V.R.; Detmann, E.; Valadares Filho, S.C.; Paulino, M.F.; Monnerat, J.P.I.S.; Duarte, M.S.; Silva, L.H.P.; Moura, L.S. Influence of Condensed Tannin on Intake, Digestibility, and Efficiency of Protein Utilization in Beef Steers Fed High Concentrate Diet. Livest. Sci. 2011, 141, 1–11. [Google Scholar] [CrossRef]
- Verma, S.; Taube, F.; Malisch, C.S. Examining the Variables Leading to Apparent Incongruity between Antimethanogenic Potential of Tannins and Their Observed Effects in Ruminants—A Review. Sustainability 2021, 13, 2743. [Google Scholar] [CrossRef]
- Kelln, B.M.; Penner, G.B.; Acharya, S.N.; McAllister, T.A.; Lardner, H.A. Impact of Condensed Tannin-Containing Legumes on Ruminal Fermentation, Nutrition, and Performance in Ruminants: A Review. Can. J. Anim. Sci. 2021, 101, 210–223. [Google Scholar] [CrossRef]
- Broderick, G.A.; Wallace, R.J.; Ørskov, E.R. Control of Rate and Extent of Protein Degradation. In Physiological Aspects of Digestion and Metabolism in Ruminants: Proceedings of the Seventh International Symposium on Ruminant Physiology; Tsuda, T., Sasaki, Y., Kawashima, R., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 541–592. [Google Scholar] [CrossRef]
- Benedeti, P.D.B.; Valadares Filho, S.D.C.; Zanetti, D.; Fonseca, E.; Silva, F.; Silva, B.D.C.; Alhadas, H.M.; Vieira Pereira, J.M.; Pacheco, M.V.C.; Pucetti, P.; et al. Prediction of In Vivo Organic Matter Digestibility of Beef Cattle Diets from Degradation Parameters Estimated from In Situ and In Vitro Incubations. J. Agric. Sci. 2019, 157, 711–720. [Google Scholar] [CrossRef]
- Billman, E.D.; Dillard, S.L.; Roca-Fernández, A.I.; Soder, K.J. Supplementation of Oilseeds to an Herbage Diet High in Condensed Tannins Affects Methane Production with Minimal Impact on Ruminal Fermentation in Continuous Culture. Fermentation 2022, 8, 109. [Google Scholar] [CrossRef]
- Detmann, E.; Souza, M.S.; Valadares Filho, S.C.; Queiroz, A.; Berchielli, T.; Saliba, E.O.; Cabral, L.S.; Pina, D.S.; Ladeira, M.; Azevedo, J. Métodos Para Análise de Alimentos. In INCT; Suprema: Visconde do Rio Branco, MG, Brazil, 2012; p. 214. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Orskov, E.; McDonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef]
- Denham, S.C.; Morantes, G.A.; Bates, D.B.; Moore, J.E. Comparison of Two Models Used to Estimate In Situ Nitrogen Disappearance. J. Dairy Sci. 1989, 72, 708–714. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2001; ISBN 0309069971. [Google Scholar]
- Calsamiglia, S.; Stern, M.D. A Three-Step In Vitro Procedure for Estimating Intestinal Digestion of Protein in Ruminants. J. Anim. Sci 1995, 73, 1459–1465. [Google Scholar] [CrossRef] [PubMed]
- Gargallo, S.; Calsamiglia, S.; Ferret, A. Technical Note: A Modified Three-Step In Vitro Procedure to Determine Intestinal Digestion of Proteins. J. Anim. Sci 2006, 84, 2163–2167. [Google Scholar] [CrossRef] [PubMed]
- Mezzomo, R.; Paulino, P.V.R.; Detmann, E.; Teixeira, C.R.V.; Alves, L.C.; Assunção, R.N. Tannin on Non-Degradable Digestible Protein from Proteic Sources in Cattle Rumen. Acta Sci. Anim. Sci. 2015, 37, 389. [Google Scholar] [CrossRef]
- O’Connor, J.D.; Sniffen, C.J.; Fox, D.G.; Chalupa, W. A Net Carbohydrate and Protein System for Evaluating Cattle Diets: IV. Predicting Amino Acid Adequacy. J. Anim. Sci 1993, 71, 1298–1311. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Bannink, A.; Crompton, L.A.; Huhtanen, P.; Kreuzer, M.; McGee, M.; Nozière, P.; Reynolds, C.K.; Bayat, A.R.; Yáñez-Ruiz, D.R.; et al. Invited Review: Nitrogen in Ruminant Nutrition: A Review of Measurement Techniques. J. Dairy Sci 2019, 102, 5811–5852. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. Exploitation of Dietary Tannins to Improve Rumen Metabolism and Ruminant Nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef]
- Castro-Montoya, J.; Westreicher-Kristen, E.; Henke, A.; Diaby, M.; Susenbeth, A.; Dickhoefer, U. In Vitro Microbial Protein Synthesis, Ruminal Degradation and Post-Ruminal Digestibility of Crude Protein of Dairy Rations Containing Quebracho Tannin Extract. J. Anim. Physiol. Anim. Nutr. 2018, 102, e77–e86. [Google Scholar] [CrossRef]
- Min, B.R.; Castleberry, L.; Allen, H.; Parker, D.; Waldrip, H.; Brauer, D.; Willis, W. Associative Effects of Wet Distiller’s Grains plus Solubles and Tannin-Rich Peanut Skin Supplementation on In Vitro Rumen Fermentation, Greenhouse Gas Emissions, and Microbial Changes. J. Anim. Sci 2019, 97, 4668–4681. [Google Scholar] [CrossRef]
- Dijkstra, J.; France, J.; Forbes, J.M. Introduction: Quantitative aspects of ruminant digestion and metabolism. In Quantitative Aspects of Ruminant Digestion and Metabolism, 2nd ed.; Dijkstra, J., Forbes, J.M., France, J., Eds.; CABI publishing: Washington, DC, USA, 2005; pp. 13–22. ISBN 0851998143. [Google Scholar]
- Yanza, Y.R.; Fitri, A.; Suwignyo, B.; Elfahmi; Hidayatik, N.; Kumalasari, N.R.; Irawan, A.; Jayanegara, A. The Utilisation of Tannin Extract as a Dietary Additive in Ruminant Nutrition: A Meta-Analysis. Animals 2021, 11, 3317. [Google Scholar] [CrossRef]
- Ahnert, S.; Dickhoefer, U.; Schulz, F.; Susenbeth, A. Influence of Ruminal Quebracho Tannin Extract Infusion on Apparent Nutrient Digestibility, Nitrogen Balance, and Urinary Purine Derivatives Excretion in Heifers. Livest. Sci. 2015, 177, 63–70. [Google Scholar] [CrossRef]
- Aboagye, I.A.; Beauchemin, K.A. Potential of Molecular Weight and Structure of Tannins to Reduce Methane Emissions from Ruminants: A Review. Animals 2019, 9, 856. [Google Scholar] [CrossRef]
- Adejoro, F.A.; Hassen, A.; Thantsha, M.S. Preparation of Acacia Tannin Loaded Lipid Microparticles by Solid-in-Oil-in-Water and Melt Dispersion Methods, Their Characterization and Evaluation of Their Effect on Ruminal Gas Production In Vitro. PLoS ONE 2018, 13, e0206241. [Google Scholar] [CrossRef] [PubMed]
- Koenig, K.M.; Beauchemin, K.A. Effect of Feeding Condensed Tannins in High Protein Finishing Diets Containing Corn Distillers Grains on Ruminal Fermentation, Nutrient Digestibility, and Route of Nitrogen Excretion in Beef Cattle. J. Anim. Sci 2018, 96, 4398–4413. [Google Scholar] [CrossRef] [PubMed]
- Barros, F.; Awika, J.M.; Rooney, L.W. Interaction of Tannins and Other Sorghum Phenolic Compounds with Starch and Effects on In Vitro Starch Digestibility. J. Agric. Food Chem. 2012, 60, 11609–11617. [Google Scholar] [CrossRef] [PubMed]
- Salami, S.A.; Luciano, G.; O’Grady, M.N.; Biondi, L.; Newbold, C.J.; Kerry, J.P.; Priolo, A. Sustainability of Feeding Plant By-Products: A Review of the Implications for Ruminant Meat Production. Anim. Feed Sci. Technol 2019, 251, 37–55. [Google Scholar] [CrossRef]
- Chamadia, B.; Grewal, R.S.; Lamba, J.S.; Kaur, J.; Kashyap, N. Effect of Varying Levels of Tannins Treatment on In Vitro Degradability of Soybean Meal. Int. J. Curr. Microbiol.App. Sci 2020, 9, 3991–4000. [Google Scholar] [CrossRef]
- Lavrenčič, A.; Levart, A. In Vitro Dry Matter and Crude Protein Rumen Degradation and Abomasal Digestibility of Soybean Meal Treated with Chestnut and Quebracho Wood Extracts. Food Sci. Nutr. 2021, 9, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Frutos, P.; Hervás, G.; Giráldez, F.J.; Mantecón, A.R. Review. Tannins and Ruminant Nutrition. Pan. J. Agric. Res. 2004, 2, 191. [Google Scholar] [CrossRef]
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The Effect of Condensed Tannins on the Nutrition and Health of Ruminants Fed Fresh Temperate Forages: A Review. Anim. Feed Sci. Technol 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Avila, A.S.; Zambom, M.A.; Faccenda, A.; Fischer, M.L.; Anschau, F.A.; Venturini, T.; Tinini, R.C.R.; Dessbesell, J.G.; Faciola, A.P. Effects of Black Wattle (Acacia Mearnsii) Condensed Tannins on Intake, Protozoa Population, Ruminal Fermentation, and Nutrient Digestibility in Jersey Steers. Animals 2020, 10, 1011. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Composition | Cottonseed Meal | Peanut Meal | Soybean Meal |
|---|---|---|---|
| Dry matter, g/kg | 910 | 915 | 910 |
| Crude protein, g/kg DM | 543 | 628 | 524 |
| Ether extract, g/kg DM | 16.4 | 12.8 | 19.4 |
| Neutral detergent fiber, g/kg DM | 174 | 164 | 157 |
| Ash, g/kg DM | 63.5 | 53.9 | 62.3 |
| Item 1 | Tannin Inclusion, g/kg | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | Linear | Quadratic | Cubic | ||
| Dry matter | ||||||||
| A, g/kg | 194 | 113 | 104 | 112 | 34.7 | <0.01 | <0.01 | 0.13 |
| B, g/kg | 595 | 629 | 644 | 599 | 7.93 | 0.82 | 0.10 | 0.71 |
| kd, h−1 | 0.059 | 0.049 | 0.041 | 0.051 | 0.002 | 0.10 | 0.03 | 0.45 |
| Crude protein | ||||||||
| A, g/kg | 345 | 315 | 331 | 293 | 5.76 | <0.01 | 0.71 | 0.03 |
| B, g/kg | 581 | 572 | 561 | 569 | 10.4 | 0.72 | 0.77 | 0.87 |
| kd, h−1 | 0.084 | 0.060 | 0.046 | 0.044 | 0.001 | <0.01 | 0.08 | 0.92 |
| Item 1 | Tannin Inclusion, g/kg | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | Linear | Quadratic | Cubic | ||
| Dry matter | ||||||||
| A, g/kg | 250 | 228 | 212 | 231 | 398 | 0.04 | <0.01 | 0.40 |
| B, g/kg | 757 | 832 | 749 | 791 | 23.5 | 0.86 | 0.48 | <0.01 |
| kd, h−1 | 0.043 | 0.040 | 0.047 | 0.036 | 0.003 | 0.56 | 0.38 | 0.15 |
| Crude protein | ||||||||
| A, g/kg | 195 | 219 | 226 | 128 | 3.50 | <0.01 | <0.01 | 0.07 |
| B, g/kg | 829 | 772 | 838 | 885 | 13.9 | 0.08 | 0.08 | 0.29 |
| kd, h−1 | 0.041 | 0.048 | 0.037 | 0.040 | 0.001 | 0.61 | 0.72 | 0.25 |
| Item 1 | Tannin Inclusion, g/kg | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | Linear | Quadratic | Cubic | ||
| Dry matter | ||||||||
| A, g/kg | 287 | 182 | 208 | 183 | 3.19 | <0.01 | <0.01 | <0.01 |
| B, g/kg | 731 | 698 | 785 | 667 | 6.43 | 0.32 | 0.08 | <0.01 |
| kd, h−1 | 0.049 | 0.042 | 0.049 | 0.050 | 0.002 | 0.68 | 0.41 | 0.34 |
| Crude protein | ||||||||
| A, g/kg | 128 | 147 | 197 | 117 | 5.02 | 0.70 | <0.01 | <0.01 |
| B, g/kg | 912 | 811 | 926 | 903 | 17.4 | <0.01 | <0.01 | <0.01 |
| kd, h−1 | 0.038 | 0.025 | 0.030 | 0.062 | 0.002 | <0.01 | <0.01 | 0.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loregian, K.E.; Pereira, D.A.B.; Rigon, F.; Magnani, E.; Marcondes, M.I.; Baumel, E.A.; Branco, R.H.; Del Bianco Benedeti, P.; Paula, E.M. Effect of Tannin Inclusion on the Enhancement of Rumen Undegradable Protein of Different Protein Sources. Ruminants 2023, 3, 413-424. https://doi.org/10.3390/ruminants3040034
Loregian KE, Pereira DAB, Rigon F, Magnani E, Marcondes MI, Baumel EA, Branco RH, Del Bianco Benedeti P, Paula EM. Effect of Tannin Inclusion on the Enhancement of Rumen Undegradable Protein of Different Protein Sources. Ruminants. 2023; 3(4):413-424. https://doi.org/10.3390/ruminants3040034
Chicago/Turabian StyleLoregian, Kalista E., David A. B. Pereira, Fernanda Rigon, Elaine Magnani, Marcos I. Marcondes, Eduardo A. Baumel, Renata H. Branco, Pedro Del Bianco Benedeti, and Eduardo M. Paula. 2023. "Effect of Tannin Inclusion on the Enhancement of Rumen Undegradable Protein of Different Protein Sources" Ruminants 3, no. 4: 413-424. https://doi.org/10.3390/ruminants3040034