

Characterization of an Acidogenic Bacterial Consortium as Probiotic and Its Effect on Rumen Fermentation In Vitro and In Vivo
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Broth and Agar Nutrient Medium Preparation
2.2. Acidogenic Bacterial Consortia Characterization and Culture Conditions
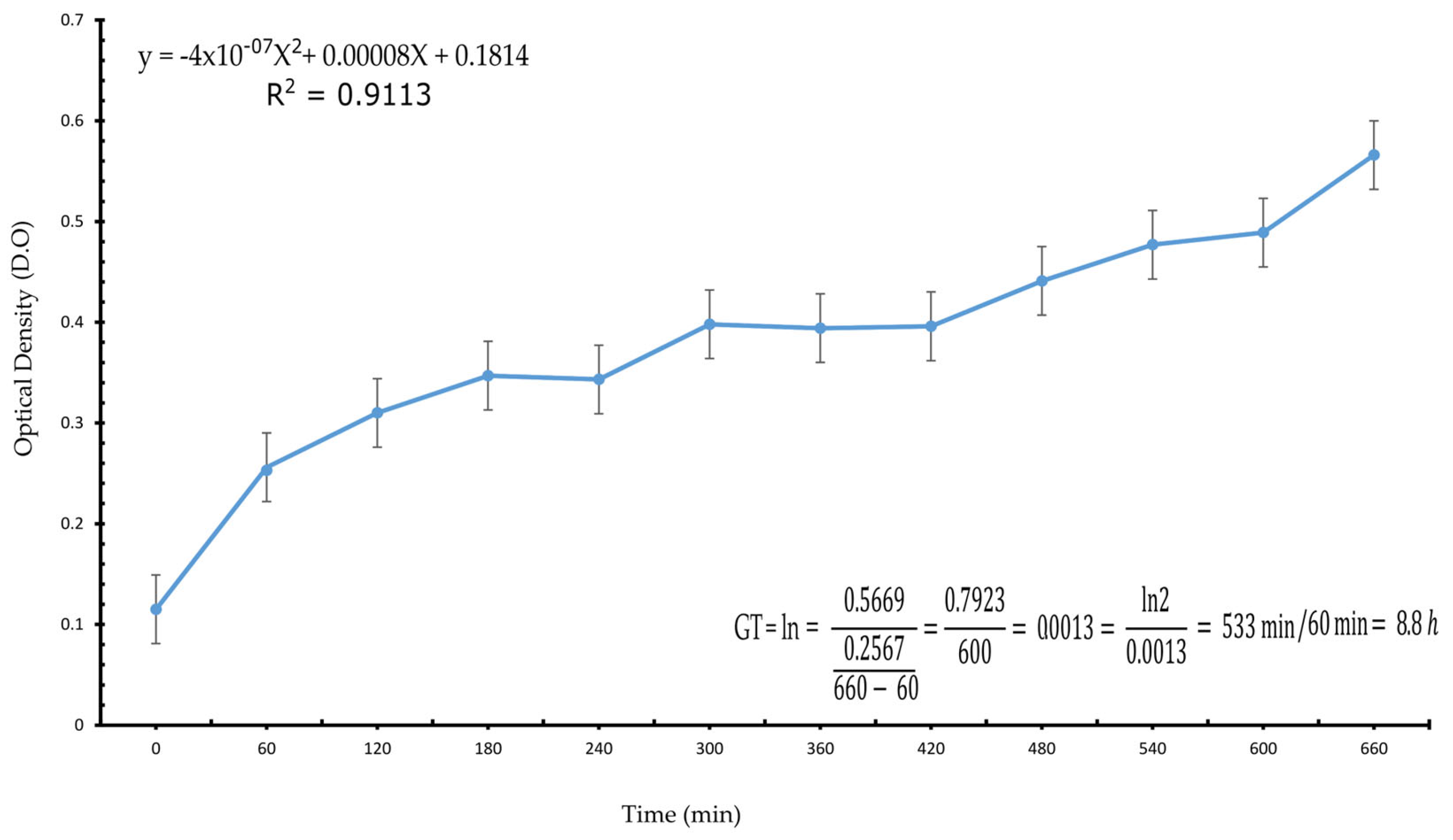
Generation Time of the Bacterial Consortium
2.3. Probiotic Tests
2.3.1. Susceptibility to Antibiotics
2.3.2. Determination of Tetracycline Resistance Genes Tet M, Tet K, and Tet W
2.3.3. Resistance to pH
2.3.4. Gastric Juice Resistance Test
2.3.5. Resistance to Bile Salts
2.3.6. Coaggregation Capacity
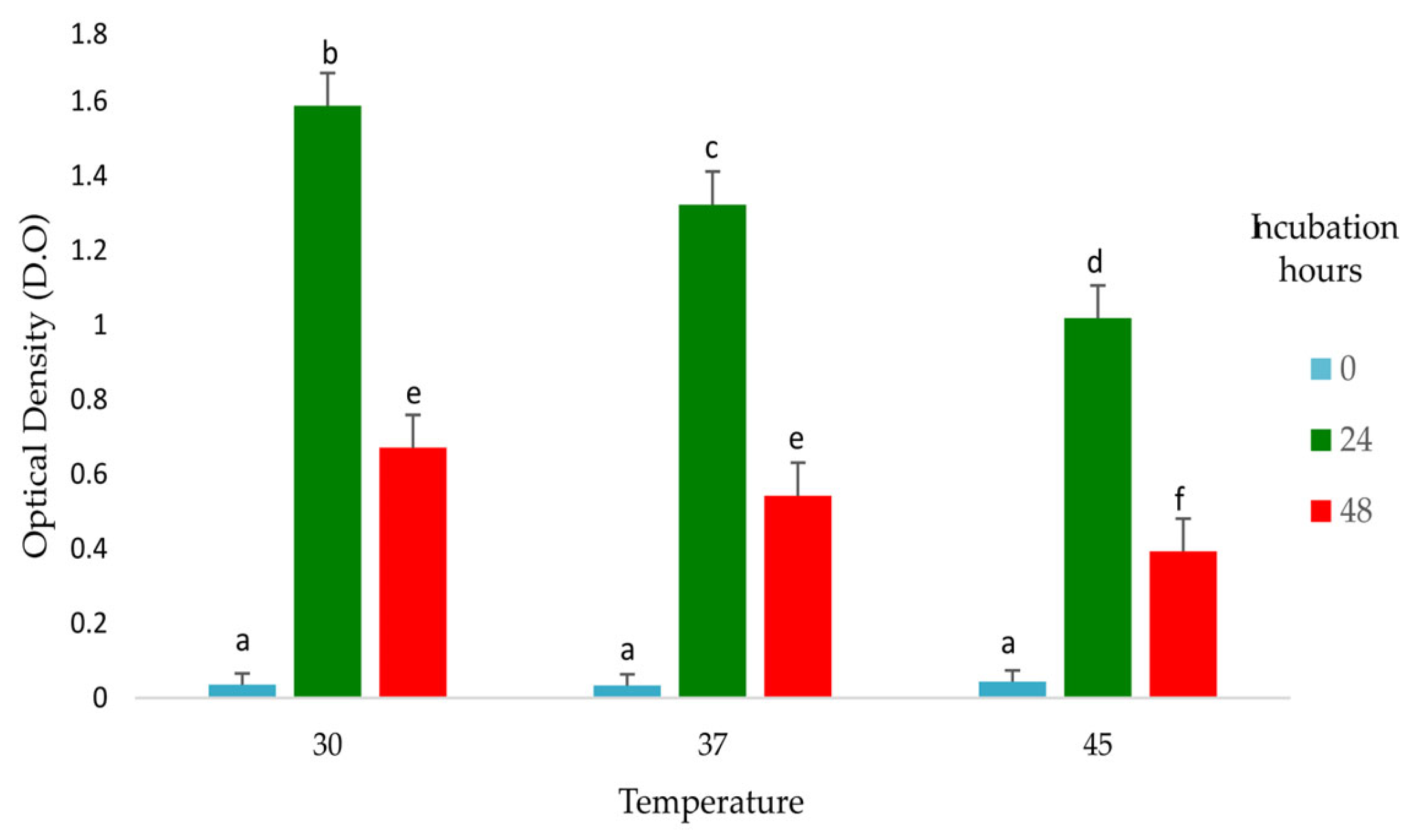
2.3.7. Tolerance to Temperature
2.3.8. Antagonistic Activity
2.3.9. Adhesion to Mucus
2.4. In Vitro Disappearance of Dry Matter and VFA Production
2.5. Effect of the Addition of Fresh ABC to Limousin Heifers
2.5.1. DNA Extraction and Microbiota rRNA 16s Sequencing
2.5.2. Relative Quantification of Lactobacillus spp. and Propionibacterium spp.
2.6. Statistical Analysis
3. Results
3.1. Growth Rate
3.2. Probiotic Tests
3.2.1. Susceptibility to Antibiotics
3.2.2. Tetracycline Resistance Genes Tet R Tet M and Tet W
3.2.3. Resistance to pH
3.2.4. Gastric Juice Resistance Test
3.2.5. Resistance to Bile Salts
3.2.6. Tolerance to Temperature
3.2.7. Coaggregation Capacity
3.2.8. Antagonistic Activity
3.2.9. Adhesion to Mucus
3.3. Disappearance of Dry Matter and VFA Production In Vitro
3.4. Effect of the Addition of Fresh ABC to Limousin Heifers
3.4.1. Serum Glucose Determination
3.4.2. ABC Effects on VFA Production
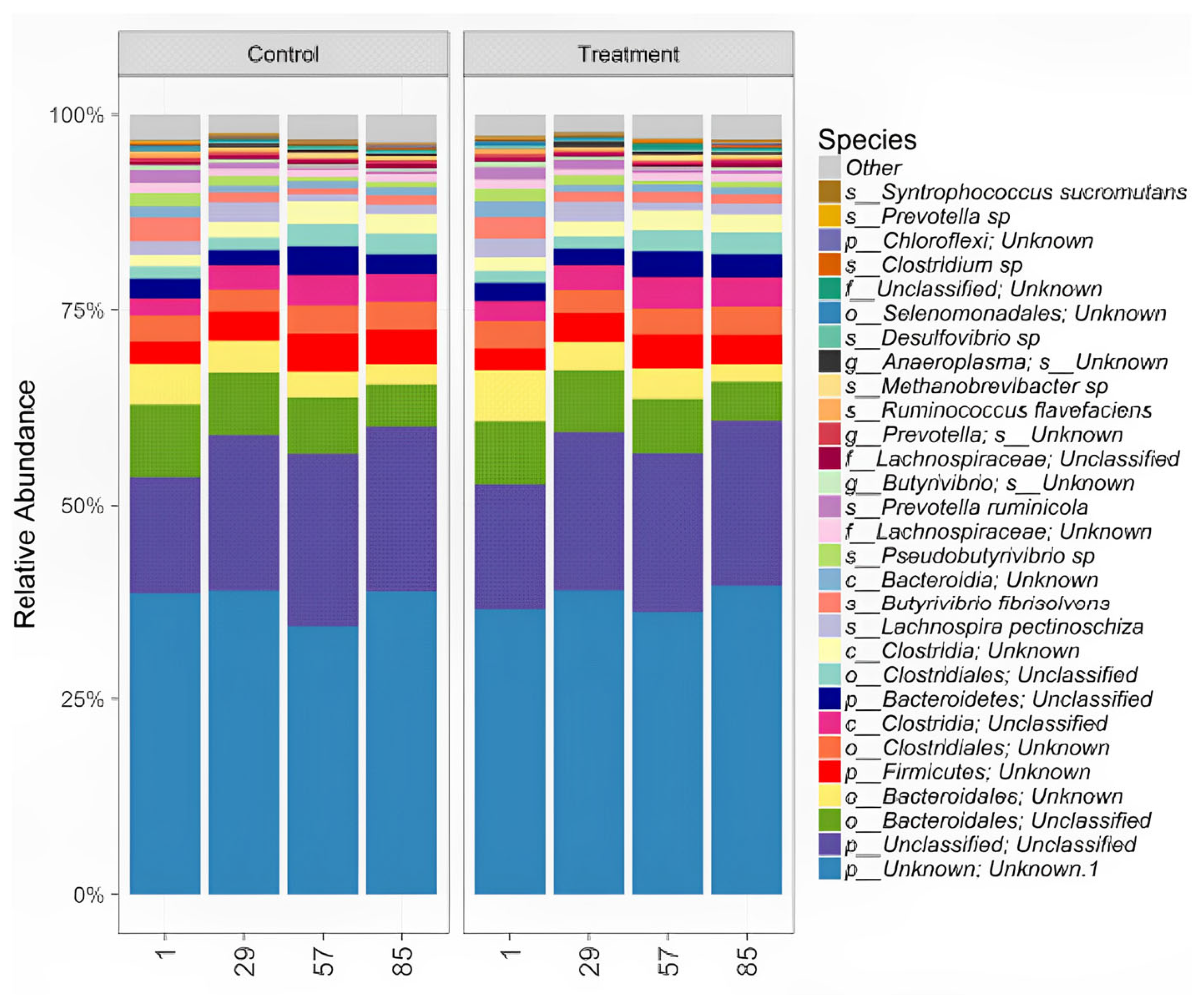
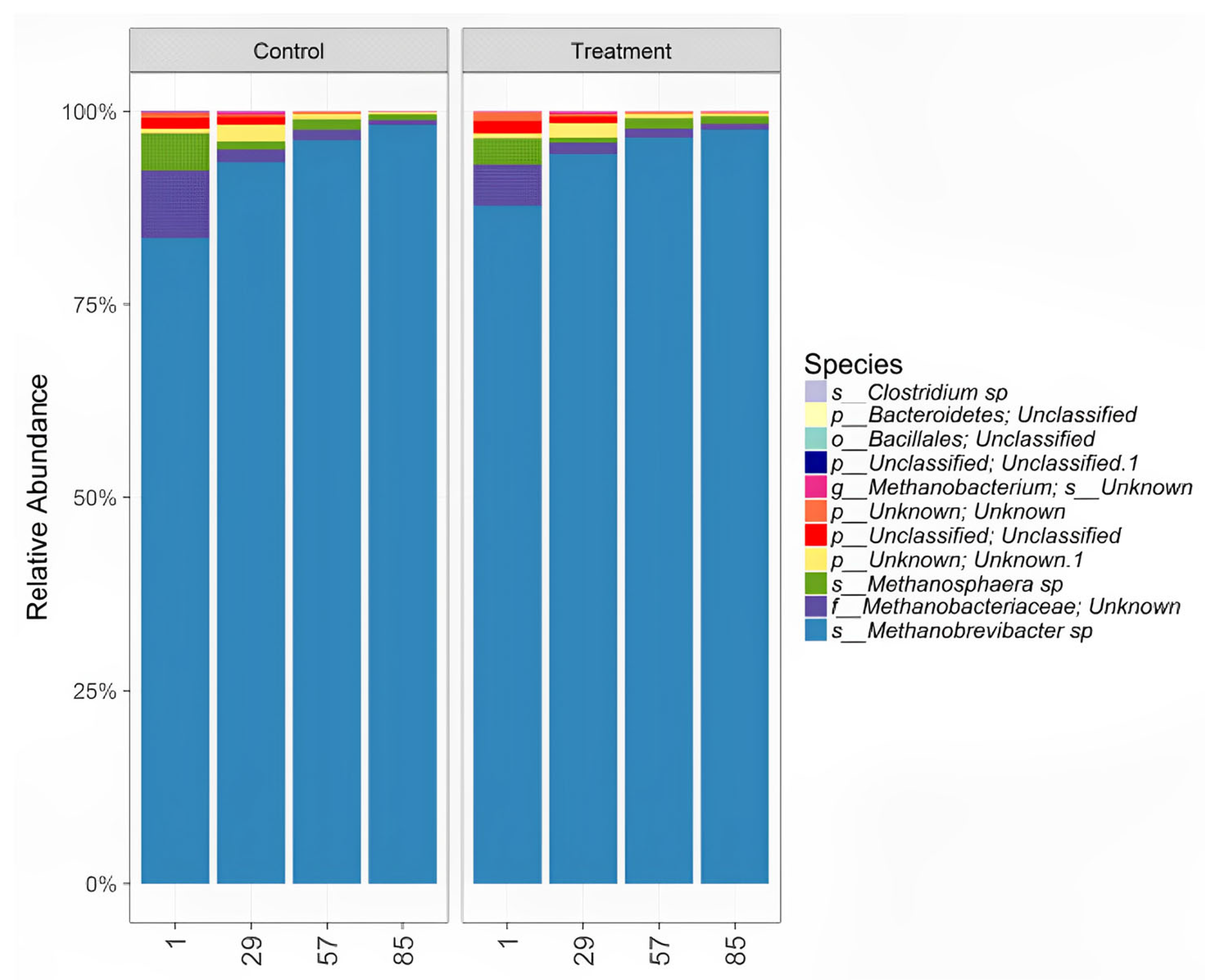
3.4.3. ABC Effects on Ruminal Microbiota
3.4.4. Relative Quantification of Lactobacillus spp. and Propionibacterium spp.
4. Discussion
4.1. Probiotic Tests
4.2. Effect of the Addition of Fresh ABC to Limousin Heifers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arowolo, M.A.; He, J. Use of probiotics and botanical extracts to improve ruminant production in the tropics: A review. Anim. Nutr. 2018, 4, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Krause, D.O.; Denman, S.E.; Mackie, R.I.; Morrison, M.; Rae, A.L.; Attwood, G.T.; McSweeney, C.S. Opportunities to improve fiber degradation in the rumen: Microbiology, ecology, and genomics. FEMS Microbiol. Rev. 2003, 27, 663–693. [Google Scholar] [CrossRef]
- García-Sorrondegui, M. Empleo de Probióticos en los Animales. 2012. Available online: https://www.engormix.com/ganaderia/levaduras-ganado-engorde/empleo-probioticos-animales_a29474/ (accessed on 14 June 2023).
- F.A.O./W.H.O. Joint FAO/WHO (Food and Agriculture Organization/World Health Organization) Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; FAO: Rome, France; WHO: Geneva, Switzerland, 2002. [Google Scholar]
- Garcia, M.; Lopez, Y.; Carcasses, A. Empleo de Probióticos en Animales. Sitio Argentino de Producción Animal. 2012. Available online: http://www.produccion-animal.com.ar/ (accessed on 14 June 2023).
- F.A.O. Biotecnología Agrícola. Available online: http://www.fao.org/3/Y5160s/y5160s07.htm (accessed on 14 June 2023).
- Guarner, F.; Perdigon, G.; Corthier, G.; Salminen, S.; Koletzko, B.; Morelli, L. Should yoghurt cultures be considered probiotic? Br. J. Nutr. 2005, 93, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Pineiro, M.; Stanton, C. Probiotic Bacteria: Legislative Framework—Requirements to Evidence Basis. J. Nutr. 2007, 137, 850S–853S. [Google Scholar] [CrossRef]
- Uyeno, Y.; Shigemori, S.; Shimosato, T. Effect of Probiotics/Prebiotics on Cattle Health and Productivity. Microbes Environ. 2015, 30, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Reuben, R.C.; Elghandour, M.M.M.Y.; Alqaisi, O.; Cone, J.W.; Márquez, O.; Salem, A.Z.M. Influence of microbial probiotics on ruminant health and nutrition: Sources, mode of action and implications. J. Sci. Food Agric. 2022, 102, 1319–1340. [Google Scholar] [CrossRef]
- Ochoa Carreño, C.; Montoya Restrepo, D.A. Microbial Consortia: A Biological Metaphor Applied to Enterprise Association in Agricultural Production Chains. Rev. Fac. Cienc. Econ. 2010, 18, 55–74. [Google Scholar] [CrossRef]
- Timmerman, H.M.; Koning, C.J.M.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics. A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef]
- Li, Z.; Lei, X.; Chen, X.; Yin, Q.; Shen, J.; Yao, J. Long-term and combined effects of N-[2-(nitrooxy)ethyl]-3-pyridinecarboxamide and fumaric acid on methane production, rumen fermentation, and lactation performance in dairy goats. J. Anim. Sci. Biotechnol. 2021, 12, 125. [Google Scholar] [CrossRef]
- Ben Shabat, S.K.; Sasson, G.; Doron-Faigenboim, A.; Durman, T.; Yaacoby, S.; Miller, M.E.B.; White, B.A.; Shterzer, N.; Mizrahi, I. Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants. ISME J. 2016, 10, 2958–2972. [Google Scholar] [CrossRef]
- Timmerman, H.; Mulder, L.; Everts, H.; van Espen, D.; van der Wal, E.; Klaassen, G.; Rouwers, S.; Hartemink, R.; Rombouts, F.; Beynen, A. Health and Growth of Veal Calves Fed Milk Replacers With or Without Probiotics. J. Dairy Sci. 2005, 88, 2154–2165. [Google Scholar] [CrossRef]
- Souza, V.; Lopes, N.; Zacaroni, O.; Silveira, V.; Pereira, R.; Freitas, J.; Almeida, R.; Salvati, G.; Pereira, M. Lactation performance and diet digestibility of dairy cows in response to the supplementation of Bacillus subtilis spores. Livest. Sci. 2017, 200, 35–39. [Google Scholar] [CrossRef]
- Chokeshaiusaha, K.; Sananmuang, T.; Puthier, D.; Nguyen, C. An innovative approach to predict immune-associated genes mutually targeted by cow and human milk microRNAs expression profiles. Vet. World 2018, 11, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141, S15–S28. [Google Scholar] [CrossRef]
- Fuller, R. A review: Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar]
- Robles, C. Adición de un Consorcio Bacteriano Altamente Propiogénico al Líquido Ruminal Ovino y su Efecto Sobre la Relación Molar Acetato: Propionato. Bachelor’s Thesis, Universidad Veracruzana, Veracruz, Mexico, 2017. [Google Scholar]
- Aguilar, M. Desarrollo de una Celda Electroquímica Microbiana con Bacterias Propionogénicas Para Incrementar la Concentración de Propionato y Disminuir la Concentración de Metano en Líquido Ruminal in Vitro. Master’s Thesis, Universidad Autónoma de México, Mexico City, Mexico, 2016. [Google Scholar]
- Ratkowsky, D.A.; Olley, J.; McMeekin, T.A.; Ball, A. Relationship between temperature and growth rate of bacterial cultures. J. Bacteriol. 1982, 149, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, A.K.; Johnson, M.C.; Ray, B. Purification, characterization and antimicrobial spectrum of a bacteriocin produced by Pediococcus acidilactici. J. Appl. Bacteriol. 1988, 65, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Cueto-Vigil, M.; Acuña-Monsalve, Y.; Valenzuela-Riaño, J. In Vitro Evaluation of Probiotic Potential of Lactic Bacteria Acid Isolated From Coastal Serum. Actual Biológicas 2010, 32, 129–138. [Google Scholar] [CrossRef]
- Yen, J.-C.; Chang, F.-J.; Chang, S. A new criterion for automatic multilevel thresholding. IEEE Trans. Image Process. 1995, 4, 370–378. [Google Scholar] [CrossRef]
- Robles, C. Aumento en la Síntesis de Propionato Ruminal Mediante el Diseño de Productos Biotecnológicos. Master’s Thesis, Universidad Autónoma de México, Mexico City, Mexico.
- Chai, Z.; Song, L.; Liu, J.; Wu, Y. Insertion depth of oral stomach tubes may affect the fermentation parameters of ruminal fluid collected in dairy cows. J. Dairy Sci. 2012, 95, 5978–5984. [Google Scholar] [CrossRef]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Ratkowsky, D.A.; Ross, T. Modelling the bacterial growth/no growth interface. Appl. Env. Microbiol. 1998, 20, 29–33. [Google Scholar] [CrossRef]
- E.F.S.A European Food Safety Authority. Introduction of a Qualified Presumption of Safety (QPS) Approach for Assessment of Selected Microorganisms Referred to EFSA Opinion of the Scientific Committee. 2007, pp. 1–16. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/587 (accessed on 8 October 2020).
- EFSA Panel on Additives and Products or Substances Used in Animal Feed (FEEDAP). EFSA J. 2018. Available online: https://www.efsa.europa.eu/en/science/scientific-committee-and-panels/feedap (accessed on 8 October 2020).
- Albuquerque, T.M.R.; Garcia, E.F.; Araújo, A.d.O.; Magnani, M.; Saarela, M.; de Souza, E.L. In Vitro Characterization of Lactobacillus Strains Isolated from Fruit Processing By-Products as Potential Probiotics. Probiotics Antimicrob. Proteins 2018, 10, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Int. J. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic Potential and Safety Evaluation of Enterococcus faecalis OB14 and OB15, Isolated From Traditional Tunisian Testouri Cheese and Rigouta, Using Physiological and Genomic Analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef]
- Kristich, C.J.; Rice, L.B.; Arias, C.A. Enterococcal infection treatment and antibiotic resistance. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Barbosa, J.; Ferreira, V.; Teixeira, P. Antibiotic susceptibility of enterococci isolated from traditional fermented meat products. Int. J. Food Microbiol. 2009, 26, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Gueimonde, M.; Sánchez, B.; de Los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Zhang, W.; Guo, H.; Pan, L.; Zhang, H.; Sun, T. Comparative studies on antibiotic resistance in Lactobacillus casei and Lactobacillus plantarum. Food Control. 2015, 50, 250–258. [Google Scholar] [CrossRef]
- Danielsen, M. Characterization of the tetracycline resistance plasmid pMD5057 from Lactobacillus plantarum 5057 reveals a composite structure. Plasmid 2002, 48, 98–103. [Google Scholar] [CrossRef]
- Krehbiel, C.R.; Rust, S.R.; Zhang, G.; Gilliland, S.E. Bacterial direct-fed microbials in ruminant diets: Performance response and mode of action. J. Anim. Sci. 2003, 81, 120–132. [Google Scholar] [CrossRef]
- Bunešová, V.; Vlková, E.; Geigerová, M.; Rada, V. Effect of rearing systems and diets composition on the survival of probiotic bifidobacteria in the digestive tract of calves. Livest. Sci. 2015, 178, 317–321. [Google Scholar] [CrossRef]
- Gueimonde, M.; Salminen, S. New methods for selecting and evaluating probiotics. Dig. Liver Dis. 2006, 38, S242–S247. [Google Scholar] [CrossRef]
- Lana, R.P.; Russell, J.B.; Van Amburgh, M.E. The role of pH in regulating ruminal methane and ammonia production. J. Anim. Sci. 1998, 76, 2190–2196. [Google Scholar] [CrossRef]
- Kern, D.L.; Slyter, L.L.; Leffel, E.C.; Weaver, J.M.; Oltjen, R.R. Ponies vs. Steers: Microbial and Chemical Characteristics of Intestinal Ingesta. J. Anim. Sci. 1974, 38, 559–564. [Google Scholar] [CrossRef]
- Wheeler, W.E.; Noller, C.H. Gastrointestinal Tract pH and Starch in Feces of Ruminants. J. Anim. Sci. 1977, 44, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, N.C.; Ficoseco, C.A.; Mansilla, F.I.; Melián, C.; Hébert, E.M.; Vignolo, G.M.; Nader-Macías, M.E.F. Identification, characterization and selection of autochthonous lactic acid bacteria as probiotic for feedlot cattle. Livest. Sci. 2018, 212, 99–110. [Google Scholar] [CrossRef]
- De Vuyst, L.; Vrancken, G.; Ravyts, F.; Rimaux, T.; Weckx, S. Biodiversity, ecological determinants, and metabolic exploitation of sourdough microbiota. Int. J. Food Microbiol. 2009, 26, 666–675. [Google Scholar] [CrossRef]
- Merritt, M.E.; Donaldson, J.R. Effect of bile salts on the DNA and membrane integrity of enteric bacteria. J. Med. Microbiol. 2009, 58, 1533–1541. [Google Scholar] [CrossRef]
- Sánchez, L.; Omura, M.; Lucas, A.; Pérez, T.; Llanes, M.; Ferreira, C. Cepas de Lactobacillus spp. con capacidades probióticas aisladas del tracto intestinal de terneros neonatos. Rev. Salud Anim. 2015, 37, 94–104. [Google Scholar]
- Vandevoorde, L.; Christiaens, H.; Verstraete, W. Prevalence of coaggregation reactions among chicken lactobacilli. J. Appl. Bacteriol. 1992, 72, 214–219. [Google Scholar] [CrossRef]
- Kolter, R.; Greenberg, E.P. Microbial sciences: The superficial life of microbes. Nature 2006, 441, 300–302. [Google Scholar] [CrossRef]
- López, A.; Espinoza, E. Evaluación del Efecto Probiótico (Pruebas de viabilidad y antagonismo in vitro) del L. plantarum en Queso Crema de Cabra Itle. Universidad de Guayaquil. 2017. Available online: http://repositorio.ug.edu.ec/handle/redug/22132 (accessed on 12 January 2020).
- Molina, A. Probióticos y su mecanismo de acción en alimentación animal. Agron Mesoam. 2019, 30, 601–611. [Google Scholar] [CrossRef]
- Ghorbani, G.R.; Morgavi, D.P.; Beauchemin, K.A.; Leedle, J.A.Z. Effects of bacterial direct-fed microbials on ruminal fermentation, blood variables, and the microbial populations of feedlot cattle. J. Anim. Sci. 2002, 80, 1977–1985. [Google Scholar] [CrossRef]
- Cagle, C.M.; Fonseca, M.A.; Callaway, T.R.; Runyan, C.A.; Cravey, M.D.; Tedeschi, L.O. Evaluation of the effects of live yeast on rumen parameters and in situ digestibility of dry matter and neutral detergent fiber in beef cattle fed growing and finishing diets. Appl. Anim. Sci. 2020, 36, 36–47. [Google Scholar] [CrossRef]
- Newbold, J. Probiotics for ruminants. Ann Zootec. 1996, 45, 329–335. [Google Scholar] [CrossRef]
- Qiao, G.H.; Shan, A.S.; Ma, N.; Ma, Q.Q.; Sun, Z.W. Effect of supplemental Bacillus cultures on rumen fermentation and milk yield in Chinese Holstein cows. J. Anim. Physiol. Anim. Nutr. 2010, 94, 429–436. [Google Scholar] [CrossRef]
- Bąkowski, M.; Kiczorowska, B. Probiotic microorganisms and herbs in ruminant nutrition as natural modulators of health and production efficiency—A review. Ann. Anim. Sci. 2021, 21, 3–28. [Google Scholar] [CrossRef]
- Arantzamendi, L.; Jiménez, G.; Blanch, A. Efecto del Probiótico Bacillus Toyoi Sobre los Parámetros Productivos de Vacuno de engorde. Rev. Ganad. 2005, 32, 30–32. [Google Scholar]
- Dick, K.; Duff, G.; Limesand, S.; Cuneo, S.; Knudson, D.; McMurphy, C.; Hall, L.; Bernal-Rigoli, J.; Marchello, J. Effects of a direct-fed microbial on digestive-tract morphology of Holstein bull calves and performance and carcass characteristics of Holstein steers. Anim. Sci. 2013, 29, 107–115. [Google Scholar] [CrossRef]
- Vyas, D.; McGeough, J.; McGinn, M.; McAllister, A.; Beauchemin, A. Effect of Propionibacterium spp. on ruminal fermentation, nutrient digestibility, and methane emissions in beef heifers fed a high-forage diet. J. Anim. Sci. 2014, 92, 2192–2201. [Google Scholar] [CrossRef]
- Tripathi, M.; Karim, S. Effect of individual and mixed live yeast culture feeding on growth performance, nutrient utilization and microbial crude protein synthesis in lambs. Anim. Feed. Sci. Technol. 2010, 155, 163–171. [Google Scholar] [CrossRef]
- Lvarez, M. Comportamiento Productivo y Digestibilidad de Nutrientes de Novillos en Engorda Suplementados con un Aditivo a Base de Probióticos y Enzimas Digestivas. Ph.D. Thesis, Universidad Autónoma de Chihuahua, Chihuahua, Mexico, 2017. [Google Scholar]
- Miyasaka, A.S. Nutrición Animal, 2nd ed; Trillas: Mexico City, Mexico, 2009. [Google Scholar]
- Carro, M.; Ranilla, M.; Tejido, M. Utilización de aditivos en la alimentación del ganado ovino y caprino. Sitio Argent. Prod Anim. 2006, 3, 26–37. [Google Scholar]
- Lehloenya, K.V.; Krehbiel, C.; Mertz, K.; Rehberger, T.; Spicer, L. Effects of Propionibacteria and Yeast Culture Fed to Steers on Nutrient Intake and Site and Extent of Digestion. J. Dairy Sci. 2008, 91, 653–662. [Google Scholar] [CrossRef]
- Stein, D.; Allen, D.; Perry, E.; Bruner, J.; Gates, K.; Rehberger, T.; Mertz, K.; Jones, D.; Spicer, L. Effects of Feeding Propionibacteria to Dairy Cows on Milk Yield, Milk Components, and Reproduction. J. Dairy Sci. 2006, 89, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Weiss, W.; Wyatt, D.; McKelvey, T. Effect of Feeding Propionibacteria on Milk Production by Early Lactation Dairy Cows. J. Dairy Sci. 2008, 91, 646–652. [Google Scholar] [CrossRef]
- Van Houtert, M.F.J.; Leng, R.A. Protein, acetate and propionate for roughage-fed lambs. 1. Body and blood composition. Anim. Prod. 1993, 56, 359–368. [Google Scholar] [CrossRef]
- Van Houtert, M.F.J. The production and metabolism of volatile fatty acids by ruminants fed roughages: A review. Anim. Feed. Sci. Technol. 1993, 43, 189–225. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Q.; Yang, W.Z.; Dong, Q.; Yang, X.M.; He, D.; Zhang, P.; Dong, K.; Huang, Y. Effects of selenium yeast on rumen fermentation, lactation performance and feed digestibilities in lactating dairy cows. Livest. Sci. 2009, 126, 239–244. [Google Scholar] [CrossRef]
- Henning, H.; Horn, H.; Leeuw, J.; Meissner, H.; Hagg, M. Effect of ruminal administration of the lactate-utilizing strain Megasphaera elsdenii (Me) NCIMB 41125 on abrupt or gradual transition from forage to concentrate diets. Anim. Feed. Sci. Technol. 2010, 157, 20–29. [Google Scholar] [CrossRef]
- Klieve, A.V.; Hennessy, D.; Ouwerkerk, D.; Forster, R.J.; Mackie, R.I.; Attwood, G.T. Establishing populations of Megasphaera elsdenii YE 34 and Butyrivibrio fibrisolvens YE 44 in the rumen of cattle fed high grain diets. J. Appl. Microbiol. 2003, 95, 621–630. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Family |
|---|---|
| Actinobacteria | Propionibacteriaceae |
| Nocardioidaceae | |
| Bacteroidetes | Rikenellaceae |
| Prophyromonadaceae | |
| Chloriflexi | Anaerolinaeceae |
| Caldilineacea | |
| Firmicutes | Ruminococcaceae |
| Clostridiaceae | |
| Veillonellaceae | |
| Peptococcaceae | |
| Enterococcaceae | |
| Nitrospira | Nitrospiraceae |
| Proteobacteria | Enterobacteriaceae |
| Syntrophobacteraceae | |
| Syntrophaceae | |
| Methyloccystaceae | |
| Desulfovibrionaceae | |
| Vibrionaceae | |
| Spirochaetes | Spirochaetaceae |
| Gram-Positive | ABC | Gram-Negative | ABC |
|---|---|---|---|
| SXT (25 μg) | Susceptible | AM (10 μg) | Susceptible |
| PE (10 u) | Resistant | AK (30 μg) | Susceptible |
| VA (30 μg) | Susceptible | GE (10 μg) | Susceptible |
| TE (30 μg) | Resistant | CF (30 μg) | Susceptible |
| AM (10 μg) | Susceptible | NET (30 μg) | Susceptible |
| CPF (5 μg) | Intermediate | CL (30 μg) | Intermediate |
| GEN (10 μg) | Resistant | SXT (25 μg) | Intermediate |
| NF (300 μg) | Susceptible |
| pH | 2.0 | 3.0 | 4.0 | 5.0 | 5.5 | 6.0 | 6.5 | 7 | SEM |
|---|---|---|---|---|---|---|---|---|---|
| 24 h | 6.6 × 107 a,+ | 3.3 × 108 a,+ | 5.0 × 109 a,b,+ | 6.0 × 109 a,b,+ | 8.7 × 109 b,+ | 1.9 × 1010 c,+ | 3.7 × 1010 e,+ | 3.5 × 1010 d,e,+ | 2.7 × 108 |
| 48 h | 0 a,+ | 0 a,+ | 3.8 × 109 a,b,+ | 9.8 × 109 b,+ | 2.6 × 1010 c,d,* | 3.0 × 1010 d,* | 4.1 × 1010 e,+ | 4.0 × 1010 e,+ | 2.7 × 108 |
| 2.0 | 3.0 | 4.0 | |
|---|---|---|---|
| CFUs | 3.0 × 107 a | 9.0 × 107 a | 5.0 × 108 b |
| SEM | 782,209.60 | 782,209.60 | 782,209.60 |
| Bacteria | 30 min | 60 min | 90 min | 120 min | 150 min |
|---|---|---|---|---|---|
| ABC | 25.0 | 100.0 | 83.3 | 65.0 | 76.7 |
| Bacteria | Incubation Broth | CFUs | SEM |
|---|---|---|---|
| ABC | NM | 1.6 × 106 | 842,120.3 |
| ABC | NM + Bile salts | 1.4 × 106 | 842,120.3 |
| Time (h) | Control | ABC 4 × 108 | ABC 4 × 1010 | ABC 4 × 1012 | SEM |
|---|---|---|---|---|---|
| 24 | 45.43 a,+ | 44.32 a,+ | 43.38 a,+ | 45.17 a,+ | 0.91 |
| 48 | 48.86 b,* | 51.73 c,* | 51.43 c,* | 51.97 c,* | 0.91 |
| Control | ABC | |||
|---|---|---|---|---|
| mMol/L | Mean | SEM | Mean | SEM |
| Acetate | 23.8 a | 5.7 | 41.6 b | 4.7 |
| Propionate | 8.9 a | 1.3 | 13.2 b | 1.09 0.86 |
| Butirate | 2.8 a | 1.0 | 6.5 b | |
| Total VFAs | 35.5 | - | 61.3 | - |
| Acetate: Propionate ratio | 2.7:1 a | 0.11 | 3.2:1 b | 0.09 |
| Productive Parameters (kg) | Control | SEM | ABC | SEM |
|---|---|---|---|---|
| IW | 274.9 a | 11.6 | 283.0 b | 14.3 |
| FW | 348.5 a | 7.1 | 371.8 b | 5.5 |
| DWG | 0.87 a | 0.04 | 1.06 b | 0.03 |
| DMI | 11.9 a | 1.7 | 11.1 b | 0.97 |
| FC | 11.4 a | 2.18 | 9.22 b | 1.80 |
| Days | Control (mg/dL) | SEM | ABC (mg/dL) | SEM |
|---|---|---|---|---|
| Day 1 | 79.07 | 4.14 | 80.46 | 4.43 |
| Day 29 | 78.27 | 4.14 | 79.05 | 4.43 |
| Day 57 | 84.48 | 4.14 | 88.86 | 4.78 |
| Day 85 | 81.69 b | 3.02 | 90.52 a | 2.62 |
| Control | SEM | ABC | SEM | |
|---|---|---|---|---|
| Acetate | 94.87 a | 71.35 | 108.46 b | 65.62 |
| Propionate | 17.60 a | 10.1 | 20.98 b | 9.01 |
| Butyrate | 7.98 a | 14.9 | 9.62 a | 15.8 |
| Total VFAs | 120.45 | 139.06 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robles-Rodríguez, C.; Cardoso-Carmona, D.; González-Dávalos, L.; Lozano-Flores, C.; Páez-Trejo, A.; Shimada, A.; Mora, O. Characterization of an Acidogenic Bacterial Consortium as Probiotic and Its Effect on Rumen Fermentation In Vitro and In Vivo. Ruminants 2023, 3, 324-346. https://doi.org/10.3390/ruminants3040028
Robles-Rodríguez C, Cardoso-Carmona D, González-Dávalos L, Lozano-Flores C, Páez-Trejo A, Shimada A, Mora O. Characterization of an Acidogenic Bacterial Consortium as Probiotic and Its Effect on Rumen Fermentation In Vitro and In Vivo. Ruminants. 2023; 3(4):324-346. https://doi.org/10.3390/ruminants3040028
Chicago/Turabian StyleRobles-Rodríguez, Carolina, Diego Cardoso-Carmona, Laura González-Dávalos, Carlos Lozano-Flores, Allan Páez-Trejo, Armando Shimada, and Ofelia Mora. 2023. "Characterization of an Acidogenic Bacterial Consortium as Probiotic and Its Effect on Rumen Fermentation In Vitro and In Vivo" Ruminants 3, no. 4: 324-346. https://doi.org/10.3390/ruminants3040028