
Effects of Feeding 60% Dried Corn Distillers’ Grains or the Equivalent Sulfur as CaSO4 on DNA Integrity and Gene Expression in Yearling Angus Bull Sperm
 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
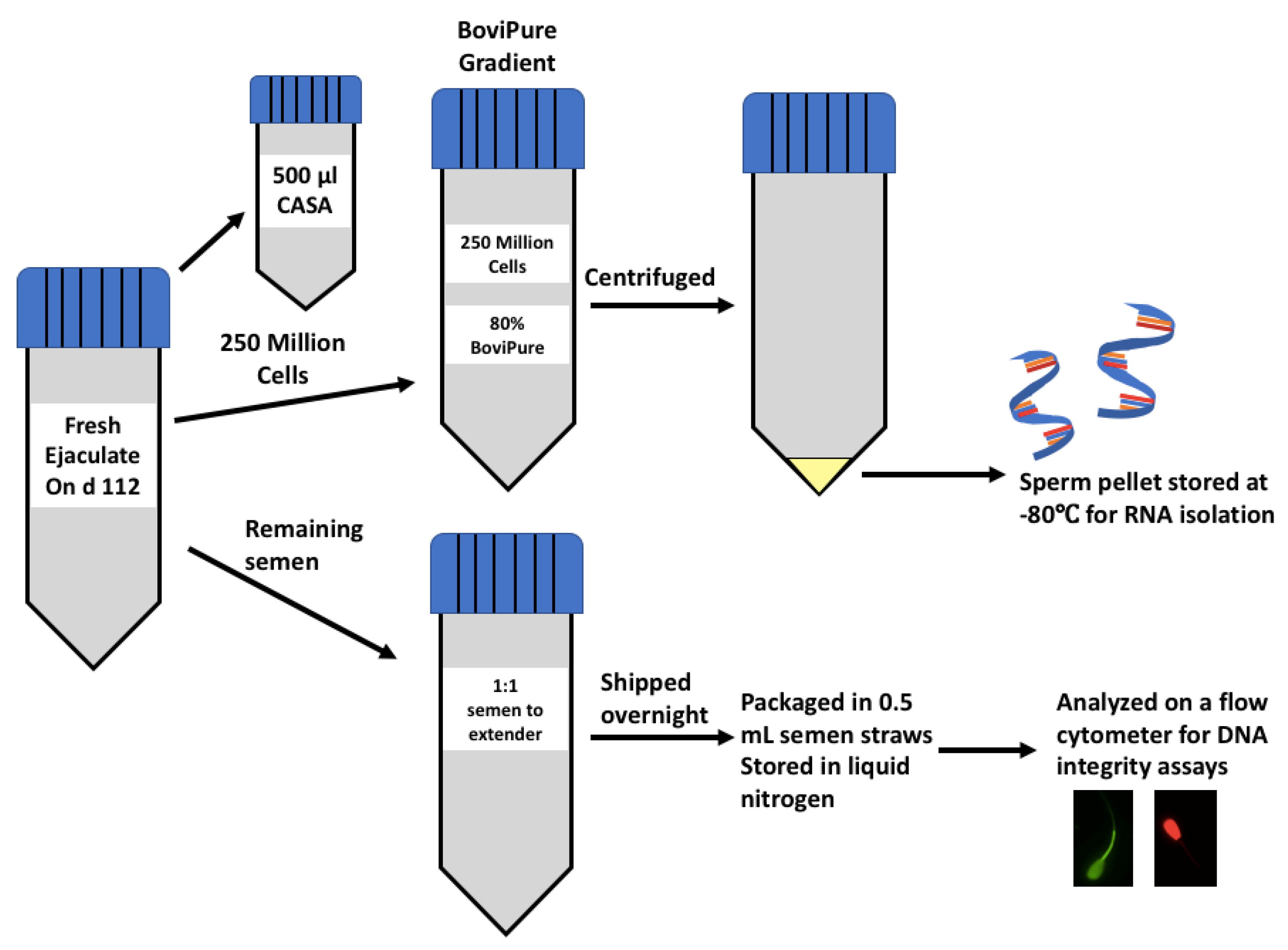
2.2. Sample Preparation for RNA Isolation
2.3. Total RNA Isolation
2.3.1. RNA Sample Quantification and Quality Evaluation
2.3.2. RNA Library Preparation and Sequencing
2.3.3. Data Quality Control and Differential Analysis
2.4. Sample Preparation for Flow Cytometry Assays
2.4.1. Mitochondrial Energy Potential
2.4.2. Oxidation Potential
2.4.3. DNA Integrity
2.4.4. Viability Assay
2.4.5. Acrosome Integrity Assay
2.4.6. Zinc Signature Assay
2.4.7. Statistical Analysis for Flow Cytometry Analysis
3. Results
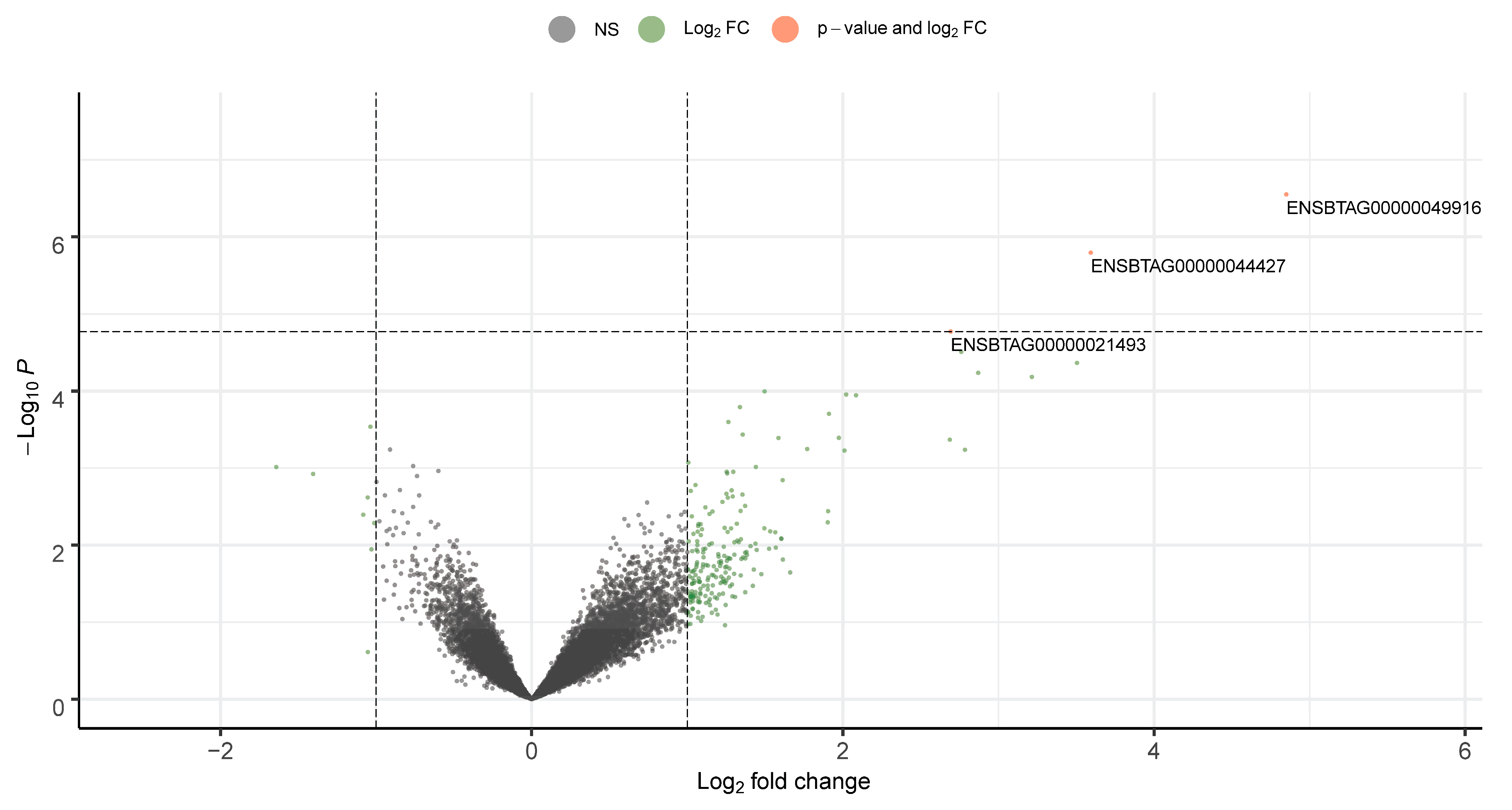
3.1. Sperm mRNA Quality and Transcript Abundance
3.2. Flow Cytometry Analysis
4. Discussion
4.1. RNA Quality and Transcript Abundance
4.2. Mitochondrial Energy Potential and DNA Integrity
4.3. Zinc Signatures in Sperm
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bollwein, H.; Janett, F.; Kaske, M. Impact of nutritional programming on the growth, health, and sexual development of bull calves. Domest. Anim. Endocrinol. 2016, 56, S180–S190. [Google Scholar] [CrossRef]
- Kenny, D.A.; Byrne, C.J. Review: The effect of nutrition on timing of pubertal onset and subsequent fertility in the bull. Animal 2018, 12, s36–s44. [Google Scholar] [CrossRef] [PubMed]
- Barth, A.D.; Brito, L.F.; Kastelic, J.P. The effect of nutrition on sexual development of bulls. Theriogenology 2008, 70, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.F.; Barth, A.D.; Wilde, R.E.; Kastelic, J.P. Effect of growth rate from 6 to 16 months of age on sexual development and reproductive function in beef bulls. Theriogenology 2012, 77, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Dance, A.; Thundathil, J.; Wilde, R.; Blondin, P.; Kastelic, J. Enhanced early-life nutrition promotes hormone production and reproductive development in Holstein bulls. J. Dairy Sci. 2015, 98, 987–998. [Google Scholar] [CrossRef]
- Kassetas, C.J.; Caton, J.S.; Kirsch, J.D.; Dorsam, S.T.; McCarthy, K.L.; Crouse, M.S.; Sedivec, K.K.; Neville, B.W.; Dahlen, C.R. Effects of feeding 60% dried corn distillers grains plus solubles or the equivalent sulfur as CaSO4 on performance and reproductive traits of yearling Angus bulls. Theriogenology 2021, 162, 6–14. [Google Scholar] [CrossRef]
- Kassetas, C.J.; Caton, J.S.; Kirsch, J.D.; Dorsam, S.T.; McCarthy, K.L.; Crouse, M.S.; Sedivec, K.K.; Neville, B.W.; Dahlen, C.R. Effects of feeding bulls dried corn distiller’s grains plus solubles or CaSO4 on mineral and metabolite concentrations in serum and seminal plasma. Anim. Reprod. Sci. 2021, 226, 106703. [Google Scholar] [CrossRef]
- Bansal, A.K.; Bilaspuri, G.S. Impacts of oxidative stress and antioxidants on semen functions. Vet. Med. Int. 2010, 2010, 686137. [Google Scholar] [CrossRef]
- Crane, A.R.; Redden, R.R.; Crouse, M.S.; Kirsch, J.D.; Borowicz, P.P.; Held, J.E.; Swanson, K.C.; Schauer, C.S. Influence of distiller’s dried grains with solubles on ram lamb growth and reproductive traits. J. Anim. Sci. 2018, 96, 1484–1494. [Google Scholar] [CrossRef]
- Van Emon, M.L.; Vonnahme, K.A.; Berg, P.T.; Redden, R.R.; Thompson, M.M.; Kirsch, J.D.; Schauer, C.S. Influence of level of dried distillers grains with solubles on feedlot performance, carcass characteristics, serum testosterone concentrations, and spermatozoa motility and concentration of growing rams. J. Anim. Sci. 2013, 91, 5821–5828. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Z.; Qie, M.; Zheng, R.; Shetty, J.; Wang, J. Sodium fluoride and sulfur dioxide affected male reproduction by disturbing blood-testis barrier in mice. Food Chem. Toxicol. 2016, 94, 103–111. [Google Scholar] [CrossRef]
- Fortes, M.R.S.; Satake, N.; Corbet, D.H.; Corbet, N.J.; Burns, B.M.; Moore, S.S.; Boe-Hansen, G.B. Sperm protamine deficiency correlates with sperm DNA damage in Bos indicus bulls. Andrology 2014, 2, 370–378. [Google Scholar] [CrossRef]
- Evenson, D.P. The Sperm Chromatin Structure Assay (SCSA(®)) and other sperm DNA fragmentation tests for evaluation of sperm nuclear DNA integrity as related to fertility. Anim. Reprod. Sci. 2016, 169, 56–75. [Google Scholar] [CrossRef]
- Johnson, C.; Dance, A.; Kovalchuk, I.; Kastelic, J.; Thundathil, J. Enhanced early-life nutrition upregulates cholesterol biosynthetic gene expression and Sertoli cell maturation in testes of pre-pubertal Holstein bulls. Sci. Rep. 2019, 9, 6448. [Google Scholar] [CrossRef]
- Geary, T.W.; Waterman, R.C.; Van Emon, M.L.; Ratzburg, C.R.; Lake, S.; Eik, B.A.; Armstrong, D.R.; Zezeski, A.L.; Heldt, J.S. Effect of supplemental trace minerals on novel measures of bull fertility1. Transl. Anim. Sci. 2019, 3, 1813–1817. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.; Zigo, M.; Sutovsky, P. Zinc: A Necessary Ion for Mammalian Sperm Fertilization Competency. Int. J. Mol. Sci. 2018, 19, 4097. [Google Scholar] [CrossRef] [PubMed]
- Kerns, K.; Zigo, M.; Drobnis, E.Z.; Sutovsky, M.; Sutovsky, P. Zinc ion flux during mammalian sperm capacitation. Nat. Commun. 2018, 9, 2061. [Google Scholar] [CrossRef] [PubMed]
- Parthipan, S.; Selvaraju, S.; Somashekar, L.; Arangasamy, A.; Sivaram, M.; Ravindra, J.P. Spermatozoal transcripts expression levels are predictive of semen quality and conception rate in bulls (Bos taurus). Theriogenology 2017, 98, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Selvaraju, S.; Swathi, D.; Ramya, L.; Lavanya, M.; Archana, S.S.; Sivaram, M. Orchestrating the expression levels of sperm mRNAs reveals CCDC174 as an important determinant of semen quality and bull fertility. Syst. Biol. Reprod. Med. 2021, 67, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Samardzija, M.; Karadjole, M.; Matkovic, M.; Cergolj, M.; Getz, I.; Dobranic, T.; Tomaskovic, A.; Petric, J.; Surina, J.; Grizelj, J.; et al. A comparison of BoviPure and Percoll on bull sperm separation protocols for IVF. Anim. Reprod. Sci. 2006, 91, 237–247. [Google Scholar] [CrossRef]
- Arias, M.E.; Andara, K.; Briones, E.; Felmer, R. Bovine sperm separation by Swim-up and density gradients (Percoll and BoviPure): Effect on sperm quality, function and gene expression. Reprod. Biol. 2017, 17, 126–132. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P.; Larson, K.L.; Jost, L.K. Sperm Chromatin Structure Assay: Its Clinical Use for Detecting Sperm DNA Fragmentation in Male Infertility and Comparisons With Other Techniques. J. Androl. 2002, 23, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using SYBR-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef]
- Tao, J.; Critser, E.S.; Critser, J.K. Evaluation of mouse sperm acrosomal status and viability by flow cytometry. Mol. Reprod. Dev. 1993, 36, 183–194. [Google Scholar] [CrossRef]
- Zoca, S.M.; Geary, T.W.; Zezeski, A.L.; Kerns, K.C.; Dalton, J.C.; Harstine, B.R.; Utt, M.D.; Cushman, R.A.; Walker, J.A.; Perry, G.A. Bull field fertility differences can be estimated with in vitro sperm capacitation and flow cytometry. Front. Anim. Sci. 2023, 4, 1180975. [Google Scholar] [CrossRef]
- Soubry, A.; Hoyo, C.; Jirtle, R.L.; Murphy, S.K. A paternal environmental legacy: Evidence for epigenetic inheritance through the male germ line. Bioessays 2014, 36, 359–371. [Google Scholar] [CrossRef]
- Stuppia, L.; Franzago, M.; Ballerini, P.; Gatta, V.; Antonucci, I. Epigenetics and male reproduction: The consequences of paternal lifestyle on fertility, embryo development, and children lifetime health. Clin. Epigenetics 2015, 7, 120. [Google Scholar] [CrossRef]
- Jodar, M. Sperm and seminal plasma RNAs: What roles do they play beyond fertilization? Reproduction 2019, 158, R113–R123. [Google Scholar] [CrossRef]
- Raval, N.P.; Shah, T.M.; George, L.B.; Joshi, C.G. Insight into bovine (Bos indicus) spermatozoal whole transcriptome profile. Theriogenology 2019, 129, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, N.; Walter, P. The Signal Recognition Particle (SRP) RNA Links Conformational Changes in the SRP to Protein Targeting. Mol. Biol. Cell 2007, 18, 2728–2734. [Google Scholar] [CrossRef]
- Sellem, E.; Jammes, H.; Schibler, L. Sperm-borne sncRNAs: Potential biomarkers for semen fertility? Reprod. Fertil. Dev. 2022, 34, 160–173. [Google Scholar] [CrossRef]
- Dogan, S.; Vargovic, P.; Oliveira, R.; Belser, L.E.; Kaya, A.; Moura, A.; Sutovsky, P.; Parrish, J.; Topper, E.; Memili, E. Sperm protamine-status correlates to the fertility of breeding bulls. Biol. Reprod. 2015, 92, 92. [Google Scholar] [CrossRef] [PubMed]
- Bissonnette, N.; Lévesque-Sergerie, J.P.; Thibault, C.; Boissonneault, G. Spermatozoal transcriptome profiling for bull sperm motility: A potential tool to evaluate semen quality. Reproduction 2009, 138, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Moura, F.H.; Macias-Franco, A.; Pena-Bello, C.A.; Archilia, E.C.; Batalha, I.M.; Silva, A.E.; Moreira, G.M.; Norris, A.B.; Schütz, L.F.; Fonseca, M.A. Sperm DNA 5-methyl cytosine and RNA N 6-methyladenosine methylation are differently affected during periods of body weight losses and body weight gain of young and mature breeding bulls. J. Anim. Sci. 2022, 100, skab362. [Google Scholar] [CrossRef]
- Sharma, U. Paternal Contributions to Offspring Health: Role of Sperm Small RNAs in Intergenerational Transmission of Epigenetic Information. Front. Cell Dev. Biol. 2019, 7, 215. [Google Scholar] [CrossRef]
- Alves, M.B.R.; Arruda, R.P.; Batissaco, L.; Garcia-Oliveros, L.N.; Gonzaga, V.H.G.; Nogueira, V.J.M.; Almeida, F.D.S.; Pinto, S.C.C.; Andrade, G.M.; Perecin, F.; et al. Changes in miRNA levels of sperm and small extracellular vesicles of seminal plasma are associated with transient scrotal heat stress in bulls. Theriogenology 2021, 161, 26–40. [Google Scholar] [CrossRef]
- Mao, S.; Sendler, E.; Goodrich, R.J.; Hauser, R.; Krawetz, S.A. A comparison of sperm RNA-seq methods. Syst. Biol. Reprod. Med. 2014, 60, 308–315. [Google Scholar] [CrossRef]
- Sellem, E.; Marthey, S.; Rau, A.; Jouneau, L.; Bonnet, A.; Perrier, J.P.; Fritz, S.; Le Danvic, C.; Boussaha, M.; Kiefer, H.; et al. A comprehensive overview of bull sperm-borne small non-coding RNAs and their diversity across breeds. Epigenetics Chromatin 2020, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Sellem, E.; Marthey, S.; Rau, A.; Jouneau, L.; Bonnet, A.; Le Danvic, C.; Guyonnet, B.; Kiefer, H.; Jammes, H.; Schibler, L. Dynamics of cattle sperm sncRNAs during maturation, from testis to ejaculated sperm. Epigenetics Chromatin 2021, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.A.; Paasch, U.; Villegas, J.V. Mitochondrial membrane potential disruption pattern in human sperm. Hum. Reprod. 2009, 24, 2079–2085. [Google Scholar] [CrossRef]
- Starkov, A.A. The role of mitochondria in reactive oxygen species metabolism and signaling. Ann. N. Y. Acad. Sci. 2008, 1147, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Garner, D.L.; Thomas, C.A.; Joerg, H.W.; DeJarnette, J.M.; Marshall, C.E. Fluorometric assessments of mitochondrial function and viability in cryopreserved bovine spermatozoa. Biol. Reprod. 1997, 57, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Qin, G.; Zhang, B. DNA damage in mice treated with sulfur dioxide by inhalation. Environ. Mol. Mutagen. 2005, 46, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Liu, Y. Cell Morphological Ultrastructural Changes in Various Organs from Mice Exposed by Inhalation to Sulfur Dioxide. Inhal. Toxicol. 2007, 19, 543–551. [Google Scholar] [CrossRef]
- Drewnoski, M.E.; Pogge, D.J.; Hansen, S.L. High-sulfur in beef cattle diets: A review. J. Anim. Sci. 2014, 92, 3763–3780. [Google Scholar] [CrossRef]
- Suarez, S.S. The oviductal sperm reservoir in mammals: Mechanisms of formation. Biol. Reprod. 1998, 58, 1105–1107. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment 1 | p-Value | ||||
|---|---|---|---|---|---|
| Item, % | CON | 60DDGS | SULF | SEM | Treatment |
| Viability Assay | |||||
| Live | 35.4 | 26.5 | 33.1 | 3.94 | 0.26 |
| Acrosome Integrity Assay | |||||
| Live intact acrosome | 42.6 | 34.2 | 35.3 | 3.81 | 0.25 |
| Oxidation Potential 2 | |||||
| Live ROS-positive | 38.5 | 28.3 | 35.8 | 4.68 | 0.29 |
| Mitochondrial Potential | |||||
| Polarized | 15.0 x | 11.7 x | 26.6 y | 4.74 | 0.08 |
| DNA Integrity Assay | |||||
| High Green 3 | 0.9 | 0.6 | 0.6 | 0.14 | 0.31 |
| DNA fragmentation index 4 | 5.7 | 4.6 | 4.7 | 0.49 | 0.19 |
| Treatment 2 | p-Value | ||||
|---|---|---|---|---|---|
| Item, % | CON | 60DDGS | SULF | SEM | Treatment |
| Signature 1 | 0.2 a | 0.1 b | 0.1 b | 0.04 | 0.04 |
| Signature 2 | 42.3 | 35.8 | 37.4 | 3.63 | 0.41 |
| Signature 3 | 53.6 | 61.4 | 59.1 | 3.69 | 0.29 |
| Signature 3 (dead) | 2.3 | 1.6 | 2.2 | 0.27 | 0.11 |
| Signature 4 | 0.6 | 0.3 | 0.3 | 0.11 | 0.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kassetas, C.J.; Geary, T.W.; Zezeski, A.L.; Caton, J.S.; Kirsch, J.D.; Dorsam, S.T.; Diniz, W.J.S.; McCarthy, K.L.; Crouse, M.S.; Sedivec, K.K.; et al. Effects of Feeding 60% Dried Corn Distillers’ Grains or the Equivalent Sulfur as CaSO4 on DNA Integrity and Gene Expression in Yearling Angus Bull Sperm. Ruminants 2023, 3, 286-298. https://doi.org/10.3390/ruminants3040026
Kassetas CJ, Geary TW, Zezeski AL, Caton JS, Kirsch JD, Dorsam ST, Diniz WJS, McCarthy KL, Crouse MS, Sedivec KK, et al. Effects of Feeding 60% Dried Corn Distillers’ Grains or the Equivalent Sulfur as CaSO4 on DNA Integrity and Gene Expression in Yearling Angus Bull Sperm. Ruminants. 2023; 3(4):286-298. https://doi.org/10.3390/ruminants3040026
Chicago/Turabian StyleKassetas, Cierrah J., Tom W. Geary, Abby L. Zezeski, Joel S. Caton, James D. Kirsch, Sheri T. Dorsam, Wellison J. S. Diniz, Kacie L. McCarthy, Matthew S. Crouse, Kevin K. Sedivec, and et al. 2023. "Effects of Feeding 60% Dried Corn Distillers’ Grains or the Equivalent Sulfur as CaSO4 on DNA Integrity and Gene Expression in Yearling Angus Bull Sperm" Ruminants 3, no. 4: 286-298. https://doi.org/10.3390/ruminants3040026