
Nutritive Value of Cistus ladanifer L. for Ruminants: Seasonal Variation of Chemical Composition, Digestibility, and Antioxidant Activity of Its Morphological Fractions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Sampling
2.2. Chemical Composition
2.3. Extract Preparation and Determination of Phenolic Compounds and Antioxidant Activity
2.3.1. Determination of Total Phenolic and Condensed Tannins Contents
2.3.2. Evaluation of Antioxidant Activity
2.4. Determination of Tocopherol Content
2.5. Statistical Analysis
3. Results
3.1. Seasonal Variation in the Morphological Composition of Cistus Ladanifer Plants
3.2. Seasonal Variation in the Proximate Composition and In Vitro Digestibility of the Morphological Fractions of Cistus Ladanifer
3.3. Seasonal Variation in the Phenolic and Tocopherol Content and Antioxidant Activity of the Morphological Fractions of Cistus Ladanifer
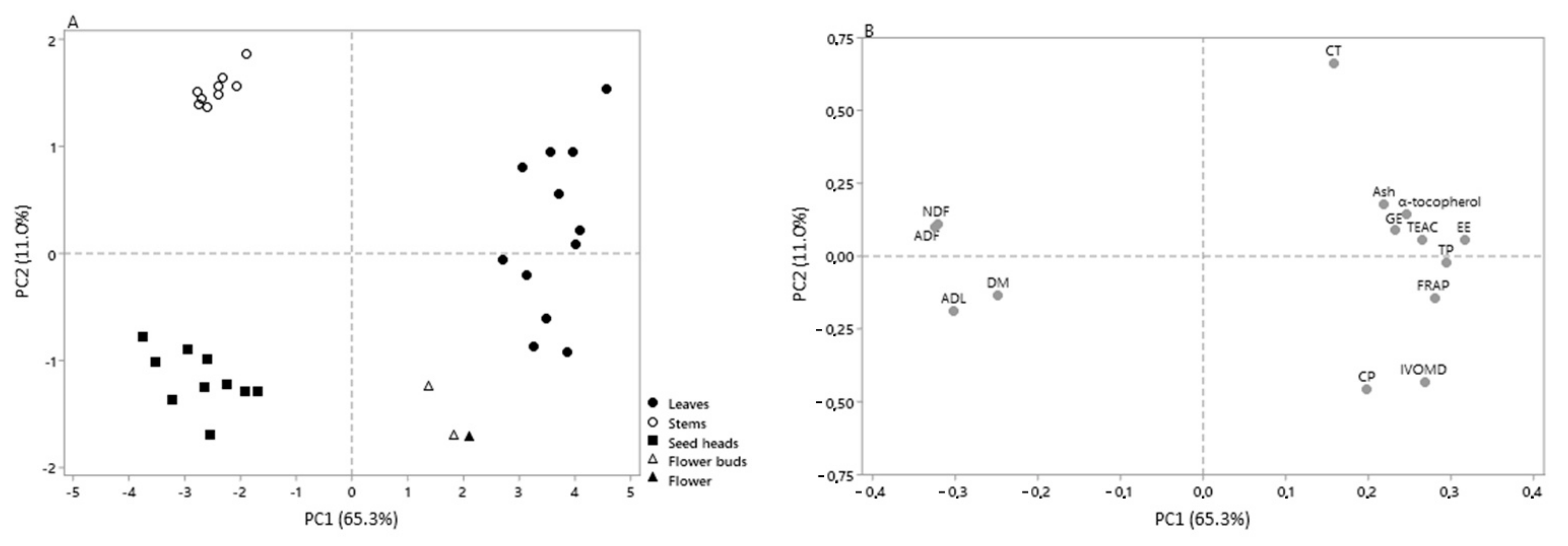
3.4. Chemical Composition, In Vitro Digestibility, Phenolic Content and Antioxidant Activity in Relation to the Morphological Fractions of Cistus Ladanifer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruno-Soares, A.M.; Matos, T.J.S.; Cadima, J. Nutritive Value of Cistus salvifolius Shrubs for Small Ruminants. Anim. Feed. Sci. Technol. 2011, 165, 167–175. [Google Scholar] [CrossRef]
- Papachristou, T.G.; Papanastasis, V.P. Forage Value of Mediterranean Deciduous Woody Fodder Species and Its Implication to Management of Silvo-Pastoral Systems for Goats. Agrofor. Syst. 1994, 27, 269–282. [Google Scholar] [CrossRef]
- Vasta, V.; Luciano, G. The Effects of Dietary Consumption of Plants Secondary Compounds on Small Ruminants’ Products Quality. Small Rumin. Res. 2011, 101, 150–159. [Google Scholar] [CrossRef]
- Vandermeulen, S.; Ramírez-Restrepo, C.A.; Beckers, Y.; Claessens, H.; Bindelle, J. Agroforestry for Ruminants: A Review of Trees and Shrubs as Fodder in Silvopastoral Temperate and Tropical Production Systems. Anim. Prod. Sci. 2018, 58, 767–777. [Google Scholar] [CrossRef]
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The Effect of Condensed Tannins on the Nutrition and Health of Ruminants Fed Fresh Temperate Forages: A Review. Anim. Feed. Sci. Technol. 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Francis, G.; Becker, K. Bioactivity of Phytochemicals in Some Lesser-Known Plants and Their Effects and Potential Applications in Livestock and Aquaculture Production Systems. Animal 2007, 1, 1371–1391. [Google Scholar] [CrossRef] [Green Version]
- Frazão, D.F.; Raimundo, J.R.; Domingues, J.L.; Quintela-Sabarís, C.; Gonçalves, J.C.; Delgado, F. Cistus Ladanifer (Cistaceae): A Natural Resource in Mediterranean-Type Ecosystems. Planta 2017, 247, 1–12. [Google Scholar] [CrossRef]
- Dentinho, M.T.P.; Moreira, O.C.; Pereira, M.S.; Bessa, R.J.B. The Use of a Tannin Crude Extract from Cistus ladanifer L. to Protect Soya-Bean Protein from Degradation in the Rumen. Animal 2007, 1, 645–650. [Google Scholar] [CrossRef] [Green Version]
- Guerreiro, O.; Dentinho, M.T.P.; Moreira, O.C.; Guerra, A.R.; Ramos, P.A.B.; Bessa, R.J.B.; Duarte, M.F.; Jerónimo, E. Potential of Cistus ladanifer L. (Rockrose) in Small Ruminant Diets—Effect of Season and Plant Age on Chemical Composition, in Vitro Digestibility and Antioxidant Activity. Grass Forage Sci. 2016, 71, 437–447. [Google Scholar] [CrossRef]
- Dentinho, M.T.P.; Navas, D.; Potes, J. Chemical and Nutritional Evaluation of Food Complements for Large Cattle Breeding, in Montado de Azinho Area. Pastagens E Forrag. 2005, 26/27, 41–46. [Google Scholar]
- Castro, M.; Teixeira, A.; Fernández-Núñez, E. The Nutritive Value of Different Mediterranean Browse Species Used as Animal Feeds under Oak Silvopastoral Systems in Northern Portugal. Agrofor. Syst. 2021, 95, 269–278. [Google Scholar] [CrossRef]
- Barrajon-Catalan, E.; Fernandez-Arroyo, S.; Saura, D.; Guillen, E.; Fernandez-Gutierrez, A.; Segura-Carretero, A.; Micol, V. Cistaceae Aqueous Extracts Containing Ellagitannins Show Antioxidant and Antimicrobial Capacity, and Cytotoxic Activity against Human Cancer Cells. Food Chem. Toxicol. 2010, 48, 2273–2282. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Duenas, M.; Alves, C.T.; Silva, S.; Henriques, M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Antifungal Activity and Detailed Chemical Characterization of Cistus ladanifer Phenolic Extracts. Ind. Crops Prod. 2013, 41, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Chaves, N.; Escudero, J.C.; Gutiérrez-Merino, C. Role of Ecological Variables in the Seasonal Variation of Flavonoid Content of Cistus ladanifer Exudate. J. Chem. Ecol. 1997, 23, 579–603. [Google Scholar] [CrossRef]
- Jerónimo, E.; Alves, S.P.; Dentinho, M.T.; Martins, S.V.; Prates, J.A.; Vasta, V.; Santos-Silva, J.; Bessa, R.J. Effect of Grape Seed Extract, Cistus ladanifer L., and Vegetable Oil Supplementation on Fatty Acid Composition of Abomasal Digesta and Intramuscular Fat of Lambs. J. Agric. Food Chem. 2010, 58, 10710–10721. [Google Scholar] [CrossRef]
- Guerreiro, O.; Alves, S.P.; Soldado, D.; Cachucho, L.; Almeida, J.M.; Francisco, A.; Santos-Silva, J.; Bessa, R.J.B.; Jeronimo, E. Inclusion of the Aerial Part and Condensed Tannin Extract from Cistus ladanifer L. in Lamb Diets–Effects on Growth Performance, Carcass and Meat Quality and Fatty Acid Composition of Intramuscular and Subcutaneous Fat. Meat Sci. 2020, 160, 107945. [Google Scholar] [CrossRef]
- Francisco, A.; Dentinho, M.T.; Alves, S.P.; Portugal, P.V.; Fernandes, F.; Sengo, S.; Jerónimo, E.; Oliveira, M.A.; Costa, P.; Sequeira, A.; et al. Growth Performance, Carcass and Meat Quality of Lambs Supplemented with Increasing Levels of a Tanniferous Bush (Cistus ladanifer L.) and Vegetable Oils. Meat Sci. 2015, 100, 275–282. [Google Scholar] [CrossRef]
- Francisco, A.; Alves, S.P.; Portugal, P.V.; Dentinho, M.T.; Jerónimo, E.; Sengo, S.; Almeida, J.; Bressan, M.C.; Pires, V.M.R.; Alfaia, C.M.; et al. Effects of Dietary Inclusion of Citrus Pulp and Rockrose Soft Stems and Leaves on Lamb Meat Quality and Fatty Acid Composition. Animal 2018, 12, 872–881. [Google Scholar] [CrossRef]
- Dentinho, M.T.P.; Belo, A.T.; Bessa, R.J.B. Digestion, Ruminal Fermentation and Microbial Nitrogen Supply in Sheep Fed Soybean Meal Treated with Cistus ladanifer L. Tannins. Small Rumin. Res. 2014, 119, 57–64. [Google Scholar] [CrossRef]
- Jerónimo, E.; Alfaia, C.M.M.; Alves, S.P.; Dentinho, M.T.P.; Prates, J.A.M.; Vasta, V.; Santos-Silva, J.; Bessa, R.J.B. Effect of Dietary Grape Seed Extract and Cistus ladanifer L. in Combination with Vegetable Oil Supplementation on Lamb Meat Quality. Meat Sci. 2012, 92, 841–847. [Google Scholar] [CrossRef]
- Jerónimo, E.; Soldado, D.; Sengo, S.; Francisco, A.; Fernandes, F.; Portugal, A.P.V.; Alves, S.P.; Santos-Silva, J.; Bessa, R.J.B. Increasing the α-Tocopherol Content and Lipid Oxidative Stability of Meat through Dietary Cistus ladanifer L. in Lamb Fed Increasing Levels of Polyunsaturated Fatty Acid Rich Vegetable Oils. Meat Sci. 2020, 164, 108092. [Google Scholar] [CrossRef] [PubMed]
- Dentinho, M.T.P.; Paulos, K.; Francisco, A.; Belo, A.T.; Jerónimo, E.; Almeida, J.; Bessa, R.J.B.; Santos-Silva, J. Effect of Soybean Meal Treatment with Cistus ladanifer Condensed Tannins in Growth Performance, Carcass and Meat Quality of Lambs. Livest. Sci. 2020, 236, 104021. [Google Scholar] [CrossRef]
- Jerónimo, E.; Cachucho, L.; Soldado, D.; Guerreiro, O.; Bessa, R.J.B.; Alves, S.P. Fatty Acid Content and Composition of the Morphological Fractions of Cistus ladanifer L. And Its Seasonal Variation. Molecules 2020, 25, 1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marta-Pedroso, C.; Freitas, H.; Domingos, T. A Estepe Cerealifera de Castro Verde. In Ecossistemas e Bem-estar Humano: Avaliação para Portugal do Millennium Ecosystem Assessment; Escolar Editora, Ed.; Escolar Editora: Lisbon, Portugal, 2009; pp. 559–583. [Google Scholar]
- International Organization for Standardization ISO 6496; Animal Feeding Stuffs–Determination of Moisture and the Other Volatile Matter Content. International Organization for Standardization: Geneva, Switzerland, 1999.
- Association of Official Analytical Chemists. Official Methods of Analysis, 15th ed.; Helrich, K., Ed.; AOAC: Arlington, VA, USA, 1990. [Google Scholar]
- Goering, H.K.; van Soest, P.J. Forage Fiber Analyses (Apparatus, Reagents, Procedures, and Some Applications). In Agricultural Research Service Handbook no 379; ARS-USDA: Washington, DC USA, 1970. [Google Scholar]
- International Organization for Standardization ISO 9831; Animal Feeding Stuffs, Animal Products and Faeces or Urine–Determination of Gross Calorimetric Value–Bomb Calorimeter Method. International Organization for Standardization: Geneva, Switzerland, 1998.
- International Organization for Standardization ISO 6492; Animal Feeding Stuffs–Determination of Fat Content. International Organization for Standardization: Geneva, Switzerland, 1999.
- Alexander, R.; Mcgowan, M. A Filtration Procedure for the in Vitro Determination of Digestibility of Herbage. J. Br. Grassl. Soc. 1966, 16, 140–147. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R. Phenolic Constituents in the Leaves of Northern Willows: Methods for the Analysis of Certain Phenolics. J. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Quantification of Tannins in Tree and Shrub Foliage: A Laboratory Manual; Kluwer Academic Publishers: Boston, MA, USA; London, UK, 2003; ISBN 1402016328. [Google Scholar]
- Fialho, L.; Ramôa, S.; Parenzan, S.; Guerreiro, I.; Catronga, H.; Soldado, D.; Guerreiro, O.; García, V.G.; e Silva, P.O.; Jerónimo, E. Effect of Regulated Deficit Irrigation on Pomegranate Fruit Quality at Harvest and during Cold Storage. Agric. Water Manag. 2021, 251, 106869. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The Conversion of Procyanidins and Prodelphinidins to Cyanidin and Delphinidin. Phytochem. 1986, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Strumeyer, D.H.; Malin, M.J. Condensed Tannins in Grain-–Isolation, Fractionation, and Characterization. J. Agric. Food Chem. 1975, 23, 909–914. [Google Scholar] [CrossRef]
- Sickel, H.; Bilger, W.; Ohlson, M. High Levels of α-Tocopherol in Norwegian Alpine Grazing Plants. J. Agric. Food Chem. 2012, 60, 7573–7580. [Google Scholar] [CrossRef]
- Talavera, S.; Gibbs, P.E.; Herrera, J. Reproductive Biology of Cistus ladanifer (Cistaceae). Plant. Syst. Evol. 1993, 186, 123–134. [Google Scholar] [CrossRef]
- Bastida, F.; Talavera, S. Temporal and Spatial Patterns of Seed Dispersal in Two Cistus Species (Cistaceae). Ann. Bot. 2002, 89, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demoly, J.P.; Montserrat, P. Cistus. Flora Iber. 1993, 3, 319–337. [Google Scholar]
- Cabezudo, B.; Navarro, T.; Pérez Latorre, A.; Nieto Caldera, J.M.; Orshan, G. Estudios Fenomorfológicos En La Vegetación Del Sur de España. I. Cistus L. (Phenomorphologic Studies in Vegetation of South of Spain. I. Cistus L.). Acta Bot. Malacit. 1992, 17, 229–237. [Google Scholar] [CrossRef]
- Ammar, H.; Lopez, S.; Gonzalez, J.S.; Ranilla, M.J. Chemical Composition and in Vitro Digestibility of Some Spanish Browse Plant Species. J. Sci. Food Agric. 2004, 84, 197–204. [Google Scholar] [CrossRef]
- Sfougaris, A.I.; Nastis, A.S.; Papageorgiou, N.K. Food Resources and Quality for the Introduced Cretan Wild Coat or Agrimi Capra Aegagrus Cretica on Atalandi Island, Greece, and Implications for Ecosystem Management. Biol. Conserv. 1996, 78, 239–245. [Google Scholar] [CrossRef]
- Piluzza, G.; Sulas, L.; Bullitta, S. Tannins in Forage Plants and Their Role in Animal Husbandry and Environmental Sustainability: A Review. Grass Forage Sci. 2014, 69, 32–48. [Google Scholar] [CrossRef]
- Jonker, A.; Yu, P. The Occurrence, Biosynthesis, and Molecular Structure of Proanthocyanidins and Their Effects on Legume Forage Protein Precipitation, Digestion and Absorption in the Ruminant Digestive Tract. Int. J. Mol. Sci. 2017, 18, 1105. [Google Scholar] [CrossRef] [Green Version]
- Treutter, D. Significance of Flavonoids in Plant Resistance: A Review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Siddhuraju, P.; Becker, K. Plant. Secondary Metabolites; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Waghorn, G. Beneficial and Detrimental Effects of Dietary Condensed Tannins for Sustainable Sheep and Goat Production-Progress and Challenges. Anim. Feed. Sci. Technol. 2008, 147, 116–139. [Google Scholar] [CrossRef]
- Frutos, P.; Hervás, G.; Natalello, A.; Luciano, G.; Fondevila, M.; Priolo, A.; Toral, P.G. Ability of Tannins to Modulate Ruminal Lipid Metabolism and Milk and Meat Fatty Acid Profiles. Anim. Feed. Sci. Technol. 2020, 269, 114623. [Google Scholar] [CrossRef]
- Jerónimo, E.; Pinheiro, C.; Lamy, E.; Dentinho, M.T.; Sales-Baptista, E.; Lopes, O.; Silva, F. Tannins in Ruminant Nutrition: Impact on Animal Performance and Quality of Edible Products. In Tannins: Biochemistry, Food Sources and Nutritional Properties; Combs, C.A., Ed.; Nova Science Publisher Inc.: Hauppauge, NY, USA, 2016; pp. 121–168. [Google Scholar]
- Caprarulo, V.; Giromini, C.; Rossi, L. Review: Chestnut and Quebracho Tannins in Pig Nutrition: The Effects on Performance and Intestinal Health. Animal 2021, 15, 100064. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and Challenges of Tannins as an Alternative to In-Feed Antibiotics for Farm Animal Production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Francisco, A.; Alves, S.P.; Portugal, P.V.; Pires, V.M.R.; Dentinho, M.T.; Alfaia, C.M.; Jeronimo, E.; Prates, J.A.M.; Santos-Silva, J.; Bessa, R.J.B. Effect of Feeding Lambs with a Tanniferous Shrub (Rockrose) and a Vegetable Oil Blend on Fatty Acid Composition of Meat Lipids. Animal 2016, 10, 2061–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, R.; Barros, L.; Carvalho, A.M.; Sousa, M.J.; Morais, J.S.; Ferreira, I.C.F.R. Aromatic Plants as a Source of Important Phytochemicals: Vitamins, Sugars and Fatty Acids in Cistus ladanifer, Cupressus lusitanica and Eucalyptus Gunnii Leaves. Ind. Crops Prod. 2009, 30, 427–430. [Google Scholar] [CrossRef]
- Goffman, F.D.; Böhme, T. Relationship between Fatty Acid Profile and Vitamin e Content in Maize Hybrids (Zea Mays l.). J. Agric. Food Chem. 2001, 49, 4990–4994. [Google Scholar] [CrossRef]
- Frank, J.; Eliasson, C.; Leroy-Nivard, D.; Budek, A.; Lundh, T.; Vessby, B.; Åman, P.; Kamal-Eldin, A. Dietary Secoisolariciresinol Diglucoside and Its Oligomers with 3-Hydroxy-3-Methyl Glutaric Acid Decrease Vitamin E Levels in Rats. Br. J. Nutr. 2004, 92, 169–176. [Google Scholar] [CrossRef]
- Ammar, H.; Lopez, S.; Gonzalez, J.S. Assessment of the Digestibility of Some Mediterranean Shrubs by in Vitro Techniques. Anim. Feed. Sci. Technol. 2005, 119, 323–331. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Leaves | Stems | Seed Heads | Flower Buds | |
|---|---|---|---|---|
| Winter | 269 a | 706 b | 15.9 | 8.84 |
| Spring | 266 a | 679 b | 19.2 | 36.1 |
| Summer | 214 b | 753 a | 33.0 | - |
| Autumn | 276 a | 692 b | 32.1 | - |
| SEM | 7.7 | 9.6 | 9.62 | 14.19 |
| p values | 0.002 | 0.003 | 0.512 | 0.246 |
| DM 1 | Ash 2 | CP 2 | EE 2 | NDF 2 | ADF 2 | ADL 2 | GE 3 | IVOMD 4 | |
|---|---|---|---|---|---|---|---|---|---|
| Leaves | |||||||||
| Winter | 417 | 54.3 a | 114 a | 73.4 | 267 | 210 ab | 63.9 | 20.2 a | 30.4 |
| Spring | 433 | 51.7 a | 73.8 ab | 59.6 | 263 | 223 a | 63.0 | 19.3 b | 34.3 |
| Summer | 567 | 35.9 b | 61.9 b | 81.3 | 249 | 186 c | 50.6 | 20.2 a | 30.1 |
| Autumn | 457 | 40.8 b | 48.5 b | 88.1 | 240 | 191 bc | 54.2 | 20.4 a | 29.3 |
| SEM | 36.0 | 3.35 | 12.81 | 6.51 | 6.3 | 7.0 | 3.64 | 2.06 | 1.24 |
| p values | 0.069 | 0.013 | 0.034 | 0.068 | 0.056 | 0.021 | 0.081 | 0.026 | 0.078 |
| Stems | |||||||||
| Winter | 605 b | 32.2 | 21.9 | 11.3 | 755 | 563 | 137 bc | 19.1 | 9.0 |
| Spring | 597 b | 31.3 | 13.8 | 8.50 | 776 | 572 | 129 c | 19.1 | 7.9 |
| Summer | 644 a | 33.5 | 18.1 | 9.07 | 740 | 562 | 145 ab | 19.3 | 11.8 |
| Autumn | 613 ab | 34.5 | 15.2 | 11.1 | 735 | 573 | 149 a | 19.3 | 11.0 |
| SEM | 10.2 | 1.14 | 2.66 | 1.581 | 14.6 | 12.7 | 31.2 | 1.75 | 0.15 |
| p values | 0.049 | 0.267 | 0.222 | 0.526 | 0.266 | 0.892 | 0.009 | 0.779 | 0.300 |
| Flower buds | |||||||||
| Winter | 371 | 25.0 | 94.7 | 57.8 | 444 | 308 | 83.5 | 20.5 | 34.4 |
| Spring | 356 | 26.8 | 48.8 | 33.5 | 524 | 386 | 122 | 19.1 | 33.5 |
| SEM | 13.0 | 0.60 | 7.98 | 14.57 | 41.4 | 45.2 | 20.9 | 3.11 | 3.04 |
| p values | 0.472 | 0.162 | 0.055 | 0.403 | 0.303 | 0.347 | 0.322 | 0.091 | 0.857 |
| Seed heads | |||||||||
| Winter | 782 | 14.7 | 40.4 | 7.43 | 708 | 574 | 181 | 19.1 | 18.0 b |
| Spring | 818 | 21.3 | 42.8 | 10.3 | 649 | 536 | 176 | 18.9 | 23.0 a |
| Summer | 843 | 29.4 | 61.3 | 10.8 | 638 | 490 | 206 | 19.3 | 23.0 a |
| Autumn | 729 | 24.1 | 51.7 | 9.60 | 679 | 522 | 210 | 19.1 | 22.6 a |
| SEM | 41.7 | 3.07 | 4.45 | 2.16 | 17.8 | 20.8 | 11.8 | 2.04 | 0.66 |
| p values | 0.327 | 0.060 | 0.051 | 0.593 | 0.106 | 0.120 | 0.225 | 0.650 | 0.006 |
| Flowers | |||||||||
| Spring | 458 | 28.4 | 53.8 | 29.6 | 286 | 177 | 37.0 | 18.6 | 34.7 |
| TP 1 | CT 2 | α-Tocopherol 3 | γ-Tocopherol 3 | TEAC 4 | FRAP 5 | |
|---|---|---|---|---|---|---|
| Leaves | ||||||
| Winter | 65.1 | 168 b | 59.5 | 26.7 b | 494 | 334 |
| Spring | 85.2 | 131 b | 143 | 57.1 a | 807 | 257 |
| Summer | 101 | 211 a | 155 | 20.8 b | 940 | 298 |
| Autumn | 90.6 | 246 a | 96 | 15.9 b | 879 | 243 |
| SEM | 7.96 | 12.2 | 37.31 | 6.43 | 120.9 | 26.4 |
| p values | 0.064 | <0.001 | 0.318 | 0.008 | 0.117 | 0.144 |
| Stems | ||||||
| Winter | 40.9 | 179 | 12.2 | n.d. | 412 | 144 |
| Spring | 34.0 | 145 | 10.4 | n.d. | 343 | 131 |
| Summer | 40.6 | 189 | 9.86 | n.d. | 461 | 163 |
| Autumn | 36.1 | 155 | 8.70 | n.d. | 352 | 152 |
| SEM | 3.91 | 16.6 | 1.201 | 40.8 | 16.7 | |
| p values | 0.571 | 0.281 | 0.105 | 0.216 | 0.620 | |
| Flower buds | ||||||
| Winter | 61.9 | 12.7 | 20.1 | 9.50 b | 609 | 280 |
| Spring | 67.8 | 35.4 | 22.4 | 9.99 a | 631 | 297 |
| SEM | 3.56 | 6.50 | 0.68 | 0.032 | 76.5 | 42.7 |
| p values | 0.363 | 0.132 | 0.139 | 0.008 | 0.854 | 0.808 |
| Seed heads | ||||||
| Winter | 34.5 | 24.0 | 9.96 | 8.22 b | 252 | 139 |
| Spring | 42.6 | 26.8 | 10.6 | 8.89 a | 350 | 191 |
| Summer | 50.0 | 24.2 | 10.8 | 8.69 a | 439 | 155 |
| Autumn | 39.9 | 20.3 | 10.7 | n.d. | 367 | 119 |
| SEM | 7.70 | 3.19 | 0.88 | 0.138 | 82.8 | 17.1 |
| p values | 0.431 | 0.641 | 0.828 | 0.018 | 0.350 | 0.068 |
| Flower | ||||||
| Spring | 94.2 | 2.04 | 29.8 | 9.03 | 1034 | 293 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerreiro, O.; Soldado, D.; Dentinho, M.T.P.; Jerónimo, E. Nutritive Value of Cistus ladanifer L. for Ruminants: Seasonal Variation of Chemical Composition, Digestibility, and Antioxidant Activity of Its Morphological Fractions. Ruminants 2022, 2, 308-320. https://doi.org/10.3390/ruminants2030021
Guerreiro O, Soldado D, Dentinho MTP, Jerónimo E. Nutritive Value of Cistus ladanifer L. for Ruminants: Seasonal Variation of Chemical Composition, Digestibility, and Antioxidant Activity of Its Morphological Fractions. Ruminants. 2022; 2(3):308-320. https://doi.org/10.3390/ruminants2030021
Chicago/Turabian StyleGuerreiro, Olinda, David Soldado, Maria T. P. Dentinho, and Eliana Jerónimo. 2022. "Nutritive Value of Cistus ladanifer L. for Ruminants: Seasonal Variation of Chemical Composition, Digestibility, and Antioxidant Activity of Its Morphological Fractions" Ruminants 2, no. 3: 308-320. https://doi.org/10.3390/ruminants2030021