
Nutritional and Physiological Constraints Contributing to Limitations in Small Intestinal Starch Digestion and Glucose Absorption in Ruminants


Abstract
:1. Introduction
2. Small Intestinal Digesta Composition and Passage Rate
2.1. Small Intestinal Digesta Composition
2.2. Small Intestinal Passage Rate
3. Small Intestinal Starch Disappearance
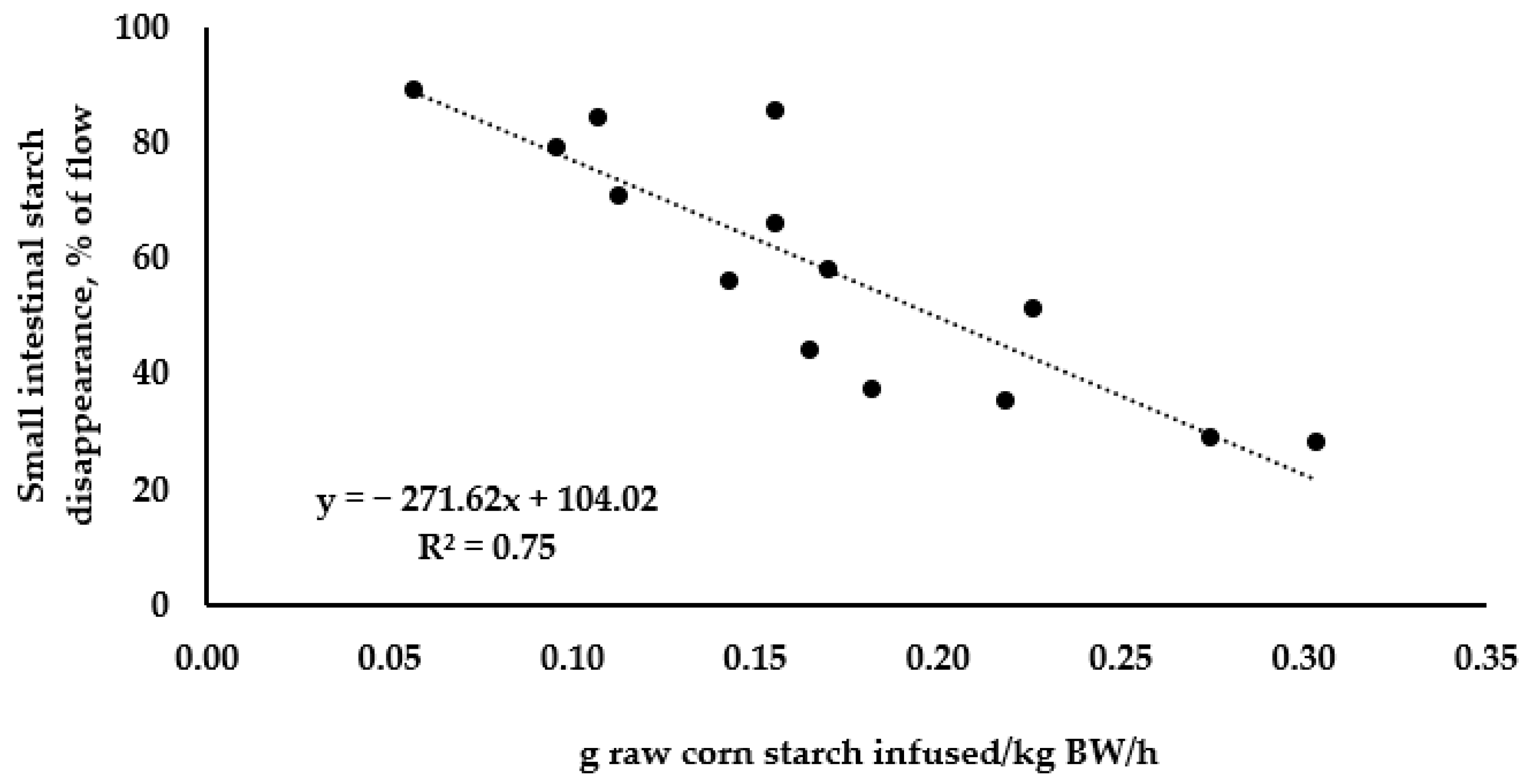
3.1. Small Intestinal Starch Disappearance: Linear Relationships
3.2. Influence of Nitrogenous Compounds on Small Intestinal Starch Disappearance
3.3. Influence of Grain Processing on Small Intestinal Starch Disappearance
4. Pancreatic α-Amylase
4.1. Influence of Dry Matter and Energy Intake on Pancreatic α-Amylase Activity
4.2. Influence of Dietary or Luminal Carbohydrate on Pancreatic α-Amylase Activity
4.3. Influence of Dietary or Luminal Nitrogenous Compounds on Pancreatic α-Amylase Activity
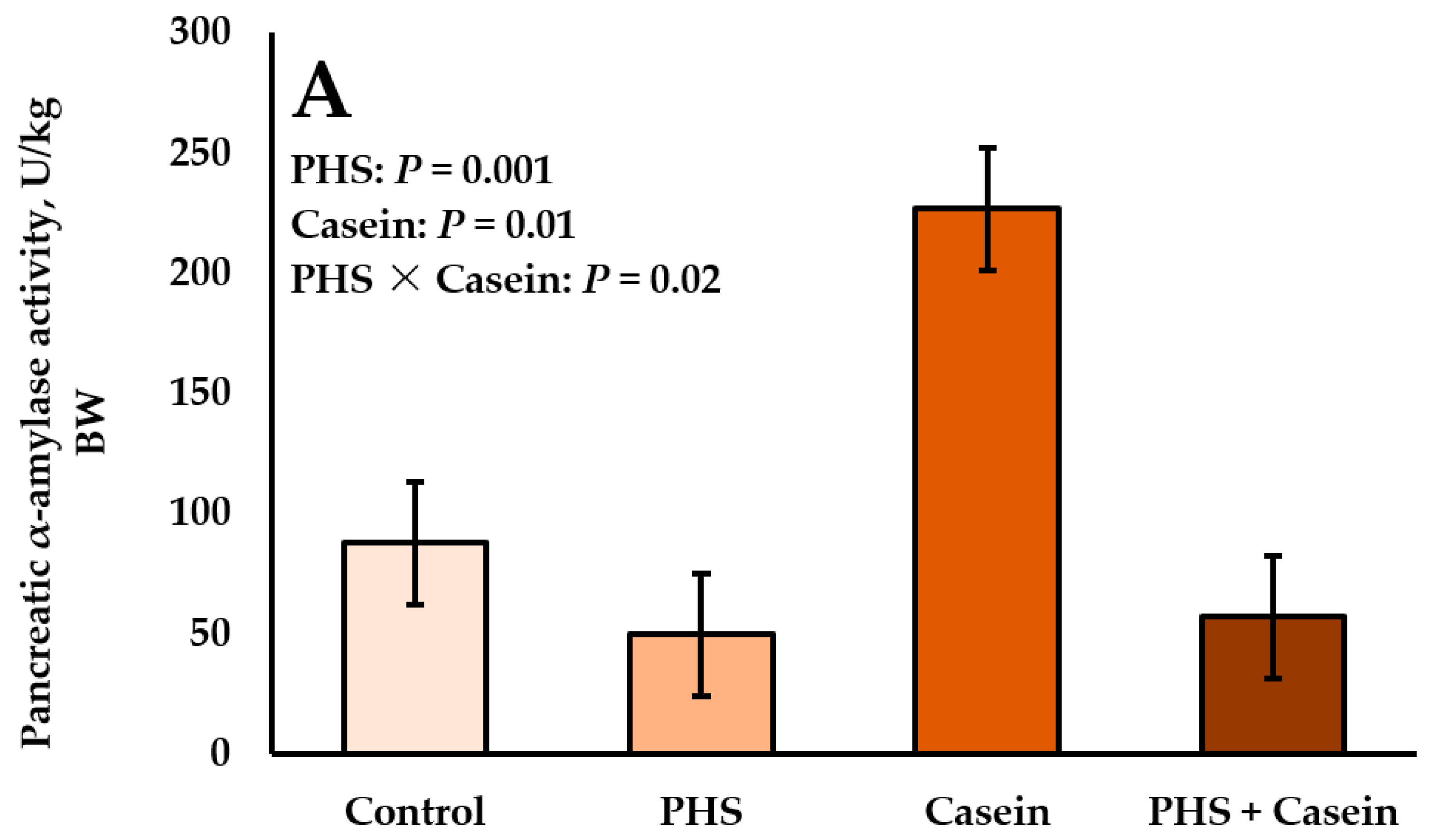
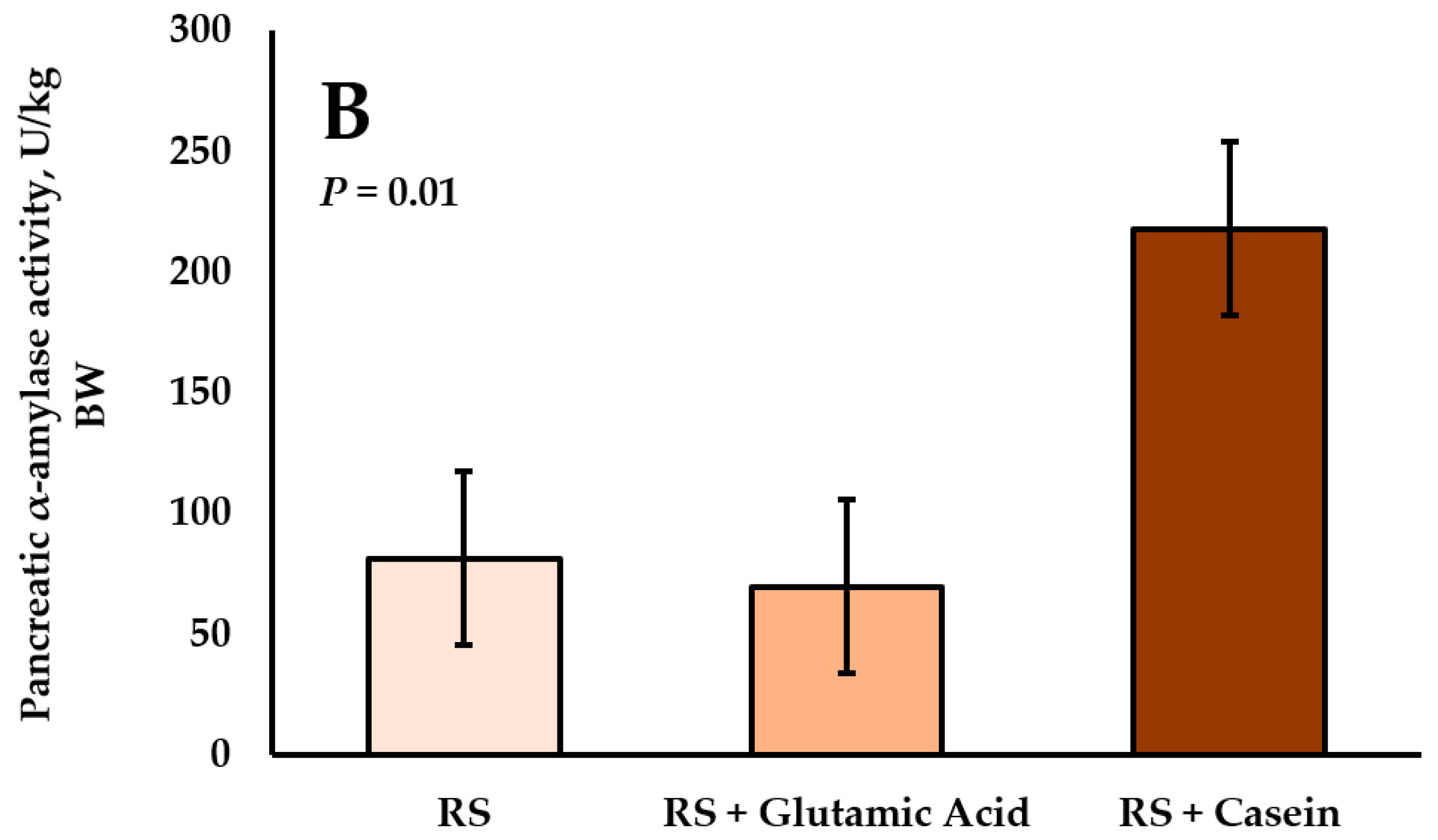
4.4. Interactions between Starch and Protein Supply on Pancreatic α-Amylase Activity
4.5. Pancreatic α-Amylase Activity and Small Intestinal Starch Disappearance
5. Small Intestinal Carbohydrases
5.1. Influence of Energy Intake on Small Intestinal Maltase, Isomaltase, Glucoamylase Activity
5.2. Influence of Macronutrients on Small Intestinal Maltase, Isomaltase, Glucoamylase Activity
6. Sucrase
6.1. Intestinal Sucrase Activity Is Absent in Multiple Ruminant Species
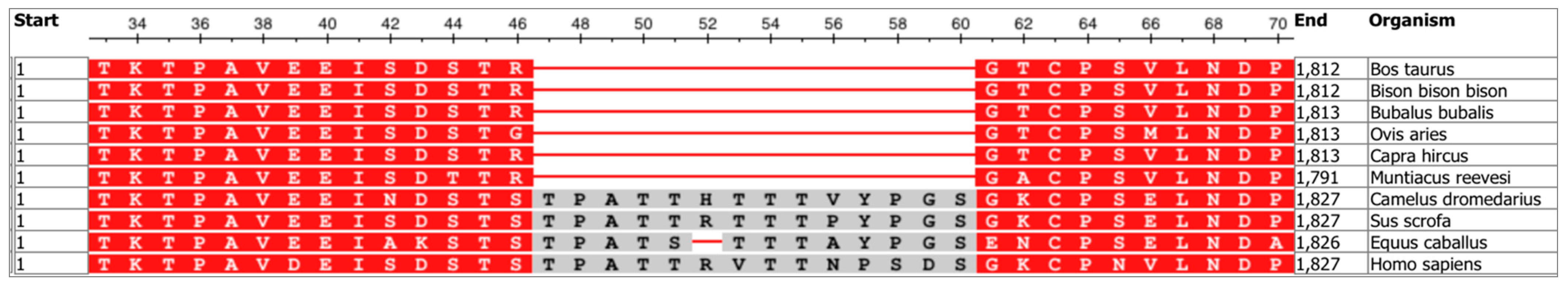
6.2. Congenital Sucrase-Isomaltase Deficiency and Multiple Sequence Alignment
6.3. Nutritional Influences on Sucrase-Isomaltase mRNA Expression and Activity
6.4. Impacts on Carbohydrate Digestion
7. Glucose Absorption
7.1. Sodium/Glucose Cotransporter-1 (SGLT1)
7.2. Glucose Transporter 5 (GLUT5)
7.3. Glucose Transporter 2 (GLUT2)
7.4. Portal Appearance of Glucose
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crouse, M.S.; Greseth, N.P.; McLean, K.J.; Crosswhite, M.R.; Pereira, N.N.; Ward, A.K.; Reynolds, L.P.; Dahlen, C.R.; Neville, B.W.; Borowicz, P.P.; et al. Maternal nutrition and stage of early pregnancy in beef heifers: Impacts on hexose and AA concentrations in maternal and fetal fluids. J. Anim. Sci. 2019, 97, 1296–1316. [Google Scholar] [CrossRef]
- Brockman, R.P.; Laarveld, B. Hormonal regulation of metabolism in ruminants: A review. Livest. Prod. Sci. 1986, 14, 313–334. [Google Scholar] [CrossRef]
- Dollar, A.M.; Porter, J.W. Utilization of carbohydrates by the young calf. Nature 1957, 179, 1299–1300. [Google Scholar] [CrossRef] [PubMed]
- Siddons, R.C.; Smith, R.H.; Henschel, M.J.; Hill, W.B.; Porter, J.W. Carbohydrate utilization in the pre-ruminant calf. Br. J. Nutr. 1969, 23, 333–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ørskov, E.R. Starch digestion and utilization in ruminants. J. Anim. Sci. 1986, 63, 1624–1633. [Google Scholar] [CrossRef] [PubMed]
- Black, J.L. A theoretical consideration of the effect of preventing rumen fermentation on the efficiency of utilization of dietary energy and protein in lambs. Br. J. Nutr. 1971, 25, 31–55. [Google Scholar] [CrossRef] [Green Version]
- Owens, F.N.; Zinn, R.A.; Kim, Y.K. Limits to starch digestion in the ruminant small intestine. J. Anim. Sci. 1986, 63, 1634–1648. [Google Scholar] [CrossRef]
- Harmon, D.L.; McLeod, K.R. Glucose uptake and regulation by intestinal tissues: Implications and whole-body energetics. J. Anim. Sci. 2001, 79, E59–E72. [Google Scholar] [CrossRef]
- Harmon, D.L.; Richards, C.J.; Swanson, K.C.; Howell, J.A.; Matthews, J.C.; True, A.D.; Huntington, G.B.; Gahr, S.A.; Russell, R.W. Influence of ruminal or postruminal starch on visceral glucose metabolism in steers. In Proceedings of the 15th Symposium on Energy Metabolism in Animals, Snekkersten, Denmark, 11–16 September 2001; pp. 273–276. [Google Scholar]
- Brake, D.W.; Swanson, K.C. RUMINANT NUTRITION SYMPOSIUM: Effects of postruminal flows of protein and amino acids on small intestinal starch digestion in beef cattle. J. Anim. Sci. 2018, 96, 739–750. [Google Scholar] [CrossRef]
- Walker, D.M. The development of the digestive system of the young animal III. Carbohydrase enzyme development in the young lamb. J. Agric. Sci. 1959, 53, 374–380. [Google Scholar] [CrossRef]
- Hembry, F.G.; Bell, M.C.; Hall, R.F. Intestinal carbohydrase activity and carbohydrate utilization in mature sheep. J. Nutr. 1967, 93, 175–181. [Google Scholar] [CrossRef]
- Siddons, R.C. Carbohydrase activities in the bovine digestive tract. Biochem. J. 1968, 108, 839–844. [Google Scholar] [CrossRef]
- Coombe, N.B.; Siddons, R.C. Carbohydrases of the bovine small intestine. Br. J. Nutr. 1973, 30, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Sir Elkhatim, M.M.; Osman, A.M. The level and distribution of disaccharidases in the camel (C. dromedarius) intestine. Comp. Biochem. Physiol. A Comp. Physiol. 1982, 71, 199–204. [Google Scholar] [CrossRef]
- Harmon, D.L. Nutritional regulation of postruminal digestive enzymes in ruminants. J. Dairy Sci. 1993, 76, 2102–2111. [Google Scholar] [CrossRef]
- Bauer, M.L.; Harmon, D.L.; Bohnert, D.W.; Branco, A.F.; Huntington, G.B. Influence of α-linked glucose on sodium-glucose cotransport activity along the small intestine in cattle. J. Anim. Sci. 2001, 79, 1917–1924. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.M.; Guimaraes, K.C.; Matthews, J.C.; McLeod, K.R.; Baldwin, R.L.; Harmon, D.L. Influence of abomasal carbohydrates on small intestinal sodium-dependent glucose cotransporter activity and abundance in steers. J. Anim. Sci. 2004, 82, 3015–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreikemeier, K.K.; Harmon, D.L.; Brandt Jr, R.T.; Avery, T.B.; Johnson, D.E. Small intestinal starch digestion in steers: Effect of various levels of abomasal glucose, corn starch and corn dextrin infusion on small intestinal disappearance and net glucose absorption. J. Anim. Sci. 1991, 69, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Huntington, G.B.; Harmon, D.L.; Richards, C.J. Sites, rates, and limits of starch digestion and glucose metabolism in growing cattle. J. Anim. Sci. 2006, 84, E14–E24. [Google Scholar] [CrossRef] [PubMed]
- Merchen, N.R. Digestion, absorption, and excretion in ruminants. In The Ruminant Animal—Digestive Physiology and Nutrition; Church, D.C., Ed.; Prentice Hall: Englewood Cliffs, NJ, USA, 1988; p. 172. [Google Scholar]
- Swanson, K.C. Small Intestinal Anatomy, Physiology, and Digestion in Ruminants. In Reference Module in Food Science; Smithers, G., Knoerzer, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- McAllan, A.B.; Smith, R.H. Carbohydrate metabolism in the ruminant. Bacterial carbohydrates formed in the rumen and their contribution to digesta entering the duodenum. Br. J. Nutr. 1974, 31, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Elsden, S.R.; Hitchcock, M.W.; Marshall, R.A.; Phillipson, A.T. Volatile acid in the digesta of ruminants and other animals. J. Exp. Biol. 1946, 22, 191–202. [Google Scholar] [CrossRef]
- Jenkins, T.C. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Doreau, M.; Ferlay, A. Digestion and utilisation of fatty acids by ruminants. Anim. Feed Sci. Technol. 1994, 45, 379–396. [Google Scholar] [CrossRef]
- Theurer, C.B. Grain processing effects on starch utilization by ruminants. J. Anim. Sci. 1986, 63, 1649–1662. [Google Scholar] [CrossRef]
- Ferraris, R.P.; Diamond, J.M. Specific regulation of intestinal nutrient transporters by their dietary substrates. Annu. Rev. Physiol. 1989, 51, 125–141. [Google Scholar] [CrossRef]
- Kreikemeier, K.K.; Harmon, D.L.; Brandt Jr, R.T.; Nagaraja, T.G.; Cochran, R.C. Steam-rolled wheat diets for finishing cattle: Effects of dietary roughage and feed intake on finishing steer performance and ruminal metabolism. J. Anim. Sci. 1990, 68, 2130–2141. [Google Scholar] [CrossRef] [Green Version]
- Staples, C.R.; Fernando, R.L.; Fahey Jr, G.C.; Berger, L.L.; Jaster, E.H. Effects of intake of a mixed diet by dairy steers on digestion events. J. Dairy Sci. 1984, 67, 995–1006. [Google Scholar] [CrossRef]
- Pearce, G.R.; Moir, R.J. Rumination in sheep. I. The influence of rumination and grinding upon the passage and digestion of food. Aust. J. Agric. Res. 1964, 15, 635–644. [Google Scholar] [CrossRef]
- desBordes, C.K.; Welch, J.G. Influence of specific gravity on rumination and passage of indigestible particles. J. Anim. Sci. 1984, 59, 470–475. [Google Scholar] [CrossRef]
- Cherney, D.J.R.; Mertens, D.R.; Moore, J.E. Fluid and particulate retention times in sheep as influenced by intake level and forage morphological composition. J. Anim. Sci. 1991, 69, 413–422. [Google Scholar] [CrossRef]
- Berger, L.L.; Klopfenstein, T.J.; Britton, R.A. Effect of sodium hydroxide treatment on rate of passage and rate of ruminal fiber digestion. J. Anim. Sci. 1980, 50, 745–749. [Google Scholar] [CrossRef]
- Zorrilla-Rios, J.; Owens, F.N.; Horn, G.W.; McNew, R.W. Effect of ammoniation of wheat straw on performance and digestion kinetics in cattle. J. Anim. Sci. 1985, 60, 814–821. [Google Scholar] [CrossRef]
- Colucci, P.E.; MacLeod, G.K.; Grovum, W.L.; McMillan, I.; Barney, D.J. Digesta kinetics in sheep and cattle fed diets with different forage to concentrate ratios at high and low intakes. J. Dairy Sci. 1990, 73, 2143–2156. [Google Scholar] [CrossRef]
- McCollum, F.T.; Galyean, M.L. Influence of cottonseed meal supplementation on voluntary intake, rumen fermentation and rate of passage of prairie hay in beef steers. J. Anim. Sci. 1985, 60, 570–577. [Google Scholar] [CrossRef]
- Prigge, E.C.; Baker, M.J.; Varga, G.A. Comparative digestion, rumen fermentation and kinetics of forage diets by steers and wethers. J. Anim. Sci. 1984, 59, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Reid, R.L.; Jung, G.A.; Cox-Ganser, J.M.; Rybeck, B.F.; Townsend, E.C. Comparative utilization of warm-and cool-season forages by cattle, sheep and goats. J. Anim. Sci. 1990, 68, 2986–2994. [Google Scholar] [CrossRef]
- Mann, D.L.; Goode, L.; Pond, K.R. Voluntary intake, gain, digestibility, rate of passage and gastrointestinal tract fill in tropical and temperate breeds of sheep. J. Anim. Sci. 1987, 64, 880–886. [Google Scholar] [CrossRef]
- Gunter, S.A.; Judkins, M.B.; Krysl, L.J.; Broesder, J.T.; Barton, R.K.; Rueda, B.R.; Hallford, D.M.; Holcombe, D.W. Digesta kinetics, ruminal fermentation characteristics and serum metabolites of pregnant and lactating ewes fed chopped alfalfa hay. J. Anim. Sci. 1990, 68, 3821–3831. [Google Scholar] [CrossRef] [PubMed]
- Coffey, K.P.; Paterson, J.A.; Saul, C.S.; Coffey, L.S.; Turner, K.E.; Bowman, J.G. The influence of pregnancy and source of supplemental protein on intake, digestive kinetics and amino acid absorption by ewes. J. Anim. Sci. 1989, 67, 1805–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westra, R.; Christopherson, R.J. Effects of cold on digestibility, retention time of digesta, reticulum motility and thyroid hormones in sheep. Can. J. Anim. Sci. 1976, 56, 699–708. [Google Scholar] [CrossRef]
- Sissons, J.W.; Thurston, S.M.; Smith, R.H. Reticular myoelectric activity and turnover of rumen digesta in the growing steer. Can. J. Anim. Sci. 1984, 64, 70–71. [Google Scholar] [CrossRef]
- Okine, E.K.; Mathison, G.W.; Hardin, R.T. Effects of changes in frequency of reticular contractions on fluid and particulate passage rates in cattle. J. Anim. Sci. 1989, 67, 3388–3396. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, H.R.; Hinds, F.C.; Bryant, M.P.; Owens, F.N. Efficiency of energy utilization by mixed rumen bacteria in continuous culture. J. Dairy Sci. 1975, 58, 1645–1659. [Google Scholar] [CrossRef]
- Prigge, E.C.; Galyean, M.L.; Owens, F.N.; Wagner, D.G.; Johnson, R.R. Microbial protein synthesis in steers fed processed corn rations. J. Anim. Sci. 1978, 46, 249–254. [Google Scholar] [CrossRef]
- Goodall, E.D.; Kay, R.N.B. Digestion and absorption in the large intestine of the sheep. J. Physiol. 1965, 176, 12–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgstaller, J.; Wittek, T.; Smith, G.W. Invited review: Abomasal emptying in calves and its potential influence on gastrointestinal disease. J. Dairy Sci. 2017, 100, 17–35. [Google Scholar] [CrossRef]
- Walker, J.A.; Harmon, D.L. Influence of ruminal or abomasal starch hydrolysate infusion on pancreatic exocrine secretion and blood glucose and insulin concentrations in steers. J. Anim. Sci. 1995, 73, 3766–3774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Ghedalia, D.; Tagari, H.; Bondi, A. Protein digestion in the intestine of sheep. Br. J. Nutr. 1974, 31, 125–142. [Google Scholar] [CrossRef]
- Zinn, R.A.; Owens, F.N. Expanding our concept of starch digestion in the ruminant. In Proceedings of the Minnesota Nutrition Conference, Bloomington, IN, USA, 22–23 September 1980; 1980; p. 69. [Google Scholar]
- Wilfart, A.; Montagne, L.; Simmins, H.; Noblet, J.; van Milgen, J. Effect of fibre content in the diet on the mean retention time in different segments of the digestive tract in growing pigs. Livest. Sci. 2007, 109, 27–29. [Google Scholar] [CrossRef]
- Croom, W.J., Jr.; Bull, L.S.; Taylor, I.L. Regulation of pancreatic exocrine secretion in ruminants: A review. J. Nutr. 1992, 122, 191–202. [Google Scholar] [CrossRef]
- Brake, D.W.; Titgemeyer, E.C.; Bailey, E.A.; Anderson, D.E. Small intestinal digestion of raw cornstarch in cattle consuming a soybean hull-based diet is improved by duodenal casein infusion. J. Anim. Sci. 2014, 92, 4047–4056. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.H. Passage of digesta through the calf abomasum and small intestine. J. Physiol. 1964, 172, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karr, M.R.; Little, C.O.; Mitchell, G.E., Jr. Starch disappearance from different segments of the digestive tract of steers. J. Anim. Sci. 1966, 25, 652–654. [Google Scholar] [CrossRef] [PubMed]
- Tucker, R.E.; Mitchell, G.E., Jr.; Little, C.O. Ruminal and postruminal starch digestion in sheep. J. Anim. Sci. 1968, 27, 824–826. [Google Scholar] [CrossRef] [PubMed]
- Little, C.O.; Mitchell, G.E.; Reitnour, C.M. Postruminal digestion of corn starch in steers. J. Anim. Sci. 1968, 27, 790–792. [Google Scholar] [CrossRef]
- Moharrery, A.; Larsen, M.; Weisbjerg, M.R. Starch digestion in the rumen, small intestine, and hind gut of dairy cows–A meta-analysis. Anim. Feed Sci. Technol. 2014, 192, 1–14. [Google Scholar] [CrossRef]
- Ørskov, E.R. The effect of processing on digestion and utilization of cereals by ruminants. Proc. Nutr. Soc. 1976, 35, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Ørskov, E.R.; Fraser, C.; Kay, R.N.B. Dietary factors influencing the digestion of starch in the rumen and small and large intestine of early weaned lambs. Br. J. Nutr. 1969, 23, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Harmon, D.L.; Yamka, R.M.; Elam, N.A. Factors affecting intestinal starch digestion in ruminants: A review. Can. J. Anim. Sci. 2004, 84, 309–318. [Google Scholar] [CrossRef]
- Kreikemeier, K.K.; Harmon, D.L. Abomasal glucose, maize starch and maize dextrin infusions in cattle: Small-intestinal disappearance, net portal glucose flux and ileal oligosaccharide flow. Br. J. Nutr. 1995, 73, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Richards, C.J.; Branco, A.F.; Bohnert, D.W.; Huntington, G.B.; Macari, M.; Harmon, D.L. Intestinal starch disappearance increased in steers abomasally infused with starch and protein. J. Anim. Sci. 2002, 80, 3361–3368. [Google Scholar] [CrossRef] [Green Version]
- Brake, D.W.; Titgemeyer, E.C.; Anderson, D.E. Duodenal supply of glutamate and casein both improve intestinal starch digestion in cattle but by apparently different mechanisms. J. Anim. Sci. 2014, 92, 4057–4067. [Google Scholar] [CrossRef] [Green Version]
- Blom, E.J.; Anderson, D.E.; Brake, D.W. Increases in duodenal glutamic acid supply linearly increase small intestinal starch digestion but not nitrogen balance in cattle. J. Anim. Sci. 2016, 94, 5332–5340. [Google Scholar] [CrossRef]
- Westreicher-Kristen, E.; Robbers, K.; Blank, R.; Tröscher, A.; Dickhoefer, U.; Wolffram, S.; Susenbeth, A. Postruminal digestion of starch infused into the abomasum of heifers with or without exogenous amylase administration. J. Anim. Sci. 2018, 96, 1939–1951. [Google Scholar] [CrossRef]
- Trotta, R.J.; Loe, E.R.; Swanson, K.C.; Bauer, M.L. Duodenal infusion of exogenous glucoamylase increases small intestinal starch disappearance in cattle. In Proceedings of the Plains Nutrition Conference, San Antonio, TX, USA, 1–3 April 2020. [Google Scholar]
- Acharya, S.; Petzel, E.A.; Bailey, E.A.; Brake, D.W. 182 Effects of duodenal casein and glutamic acid infusion on small intestinal starch digestion and energy and nitrogen balance in cattle. J. Dairy Sci. 2020, 103, 70. [Google Scholar]
- Wang, X.; Taniguchi, K. Activity of pancreatic digestive enzyme in sheep given abomasal infusion of starch and casein. Anim. Feed Sci. Technol. 1998, 69, 870–874. [Google Scholar]
- Taniguchi, K.; Huntington, G.B.; Glenn, B.P. Net nutrient flux by visceral tissues of beef steers given abomasal and ruminal infusions of casein and starch. J. Anim. Sci. 1995, 73, 236–249. [Google Scholar] [CrossRef]
- Mendoza, G.D.; Britton, R.A. Response of intestinal starch digestion to duodenal infusion of casein. J. Appl. Anim. Res. 2003, 24, 123–128. [Google Scholar] [CrossRef]
- Mabjeesh, S.J.; Guy, D.; Sklan, D. Na+/glucose co-transporter abundance and activity in the small intestine of lambs: Enhancement by abomasal infusion of casein. Br. J. Nutr. 2003, 89, 573–580. [Google Scholar] [CrossRef]
- Yu, Z.P.; Xu, M.; Wang, F.; Liu, K.; Yao, J.H.; Wu, Z.; Qin, D.K.; Sun, F.F. Effect of duodenal infusion of leucine and phenylalanine on intestinal enzyme activities and starch digestibility in goats. Livest. Sci. 2014, 162, 134–140. [Google Scholar] [CrossRef]
- Huntington, G.B. Starch utilization by ruminants: From basics to the bunk. J. Anim. Sci. 1997, 75, 852–867. [Google Scholar] [CrossRef] [PubMed]
- Owens, C.E.; Zinn, R.A.; Hassen, A.; Owens, F.N. Mathematical linkage of total-tract digestion of starch and neutral detergent fiber to their fecal concentrations and the effect of site of starch digestion on extent of digestion and energetic efficiency of cattle. Prof. Anim. Sci. 2016, 32, 531–549. [Google Scholar] [CrossRef] [Green Version]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle: Eighth Revised Edition; The National Academies Press: Washington, DC, USA, 2016; p. 494. [Google Scholar]
- Sniffen, C.J.; O’Connor, J.D.; Van Soest, P.J.; Fox, D.G.; Russell, J.B. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 1992, 70, 3562–3577. [Google Scholar] [CrossRef] [PubMed]
- Owens, F.N.; Zinn, R.A. Corn grain for cattle: Influence of processing on site and extent of digestion. In Proceedings of the 20th Southwest Nutrition and Management Conference, Tempe, AZ, USA, 24–25 February 2005; pp. 86–112. [Google Scholar]
- Zinn, R.A.; Owens, F.N.; Ware, R.A. Flaking corn: Processing mechanics, quality standards, and impacts on energy availability and performance of feedlot cattle. J. Anim. Sci. 2002, 80, 1145–1156. [Google Scholar] [CrossRef]
- Harmon, D.L. Impact of nutrition on pancreatic exocrine and endocrine secretion in ruminants: A review. J. Anim. Sci. 1992, 70, 1290–1301. [Google Scholar] [CrossRef]
- Harmon, D.L. Dietary influences on carbohydrases and small intestinal starch hydrolysis capacity in ruminants. J. Nutr. 1992, 122, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Harmon, D.L. Understanding starch utilization in the small intestine of cattle. Asian-Australas. J. Anim. Sci. 2009, 22, 915–922. [Google Scholar] [CrossRef]
- Harmon, D.L.; Swanson, K.C. Review: Nutritional regulation of intestinal starch and protein assimilation in ruminants. Animal 2020, 14, s17–s28. [Google Scholar] [CrossRef] [Green Version]
- Trotta, R.J.; Swanson, K.C. Prenatal and Postnatal Nutrition Influence Pancreatic and Intestinal Carbohydrase Activities of Ruminants. Animals 2021, 11. [Google Scholar] [CrossRef]
- Guo, L.; Yao, J.; Cao, Y. Regulation of pancreatic exocrine in ruminants and the related mechanism: The signal transduction and more. Anim. Nutr. 2021. [Google Scholar] [CrossRef]
- Brannon, P.M. Adaptation of the exocrine pancreas to diet. Annu. Rev. Nutr. 1990, 10, 85–105. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.R.; Young, A.W.; Jorgensen, N.A. Effect of dietary corn starch intake on pancreatic amylase and intestinal maltase and pH in cattle. J. Anim. Sci. 1981, 52, 1177–1182. [Google Scholar] [CrossRef] [Green Version]
- Kreikemeier, K.K.; Harmon, D.L.; Peters, J.P.; Gross, K.L.; Armendariz, C.K.; Krehbiel, C.R. Influence of dietary forage and feed intake on carbohydrase activities and small intestinal morphology of calves. J. Anim. Sci. 1990, 68, 2916–2929. [Google Scholar] [CrossRef]
- Clary, J.J.; Mitchell, G.E., Jr.; Little, C.O. Action of bovine and ovine alpha-amylases on various starches. J. Nutr. 1968, 95, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Janes, A.N.; Weekes, T.E.C.; Armstrong, D.G. Carbohydrase activity in the pancreatic tissue and small intestine mucosa of sheep fed dried-grass or ground maize-based diets. J. Agric. Sci. 1985, 104, 435–443. [Google Scholar] [CrossRef]
- Keomanivong, F.E.; Grazul-Bilska, A.T.; Redmer, D.A.; Bass, C.S.; Kaminski, S.L.; Borowicz, P.P.; Kirsch, J.D.; Swanson, K.C. The impact of diet and arginine supplementation on pancreatic mass, digestive enzyme activity, and insulin-containing cell cluster morphology during the estrous cycle in sheep. Domest. Anim. Endocrinol. 2017, 59, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keomanivong, F.E.; Lemley, C.O.; Camacho, L.E.; Yunusova, R.; Borowicz, P.P.; Caton, J.S.; Meyer, A.M.; Vonnahme, K.A.; Swanson, K.C. Influence of nutrient restriction and melatonin supplementation of pregnant ewes on maternal and fetal pancreatic digestive enzymes and insulin-containing clusters. Animal 2016, 10, 440–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trotta, R.J.; Vasquez-Hidalgo, M.A.; Vonnahme, K.A.; Swanson, K.C. Effects of nutrient restriction during midgestation to late gestation on maternal and fetal postruminal carbohydrase activities in sheep. J. Anim. Sci. 2020, 98, skz393. [Google Scholar] [CrossRef]
- Awda, B.J.; Wood, K.M.; Keomanivong, F.E.; Borowicz, P.P.; McBride, B.W.; Miller, S.P.; Fitzsimmons, C.J.; Swanson, K.C. The influence of pregnancy and plane of nutrition during pregnancy on pancreatic digestive enzymes and insulin-containing cell cluster morphology in beef cows. Can. J. Anim. Sci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Keomanivong, F.E.; Camacho, L.E.; Lemley, C.O.; Kuemper, E.A.; Yunusova, R.D.; Borowicz, P.P.; Kirsch, J.D.; Vonnahme, K.A.; Caton, J.S.; Swanson, K.C. Effects of realimentation after nutrient restriction during mid-to late gestation on pancreatic digestive enzymes, serum insulin and glucose levels, and insulin-containing cell cluster morphology. J. Anim. Physiol. Anim. Nutr. 2017, 101, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Ko, M.; Holligan, S.; McBride, B.W.; Fan, M.Z.; Swanson, K.C. Effect of dry matter intake on visceral organ mass, cellularity, and the protein expression of ATP synthase, Na+/K+-ATPase, proliferating cell nuclear antigen and ubiquitin in feedlot steers. Can. J. Anim. Sci. 2009, 89, 253–262. [Google Scholar] [CrossRef]
- Wood, K.M.; Awda, B.J.; Fitzsimmons, C.; Miller, S.P.; McBride, B.W.; Swanson, K.C. Effect of moderate dietary restriction on visceral organ weight, hepatic oxygen consumption, and metabolic proteins associated with energy balance in mature pregnant beef cows. J. Anim. Sci. 2013, 91, 4245–4255. [Google Scholar] [CrossRef] [PubMed]
- Holligan, S.; Wang, J.; Cant, J.P.; Swanson, K.C. A proteomics approach to detect tissue-wide adaptive changes in the pancreas associated with increased pancreatic α-amylase activity in domestic cattle (Bos taurus). Comp. Biochem. Physiol. Part D Genom. Proteom. 2013, 8, 65–71. [Google Scholar] [CrossRef]
- Swanson, K.C.; Matthews, J.C.; Woods, C.A.; Harmon, D.L. Postruminal administration of partially hydrolyzed starch and casein influences pancreatic α-amylase expression in calves. J. Nutr. 2002, 132, 376–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, K.C.; Richards, C.J.; Harmon, D.L. Influence of abomasal infusion of glucose or partially hydrolyzed starch on pancreatic exocrine secretion in beef steers. J. Anim. Sci. 2002, 80, 1112–1116. [Google Scholar] [CrossRef]
- Krehbiel, C.R.; Britton, R.A.; Harmon, D.L.; Wester, T.J.; Stock, R.A. The effects of ruminal acidosis on volatile fatty acid absorption and plasma activities of pancreatic enzymes in lambs. J. Anim. Sci. 1995, 73, 3111–3121. [Google Scholar] [CrossRef]
- Trotta, R.J.; Ward, A.K.; Swanson, K.C. Influence of dietary fructose supplementation on visceral organ mass, carbohydrase activity, and mRNA expression of genes involved in small intestinal carbohydrate assimilation in neonatal calves. J. Dairy Sci. 2020, 103, 10060–10073. [Google Scholar] [CrossRef]
- Richards, C.J.; Swanson, K.C.; Paton, S.J.; Harmon, D.L.; Huntington, G.B. Pancreatic exocrine secretion in steers infused postruminally with casein and cornstarch. J. Anim. Sci. 2003, 81, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Choi, C.W.; Jin, Y.C.; Wang, T.; Lee, K.H.; Ku, M.B.; Hwang, J.H.; Kim, K.H.; Vega, R.S.A.; Lee, H.G. Effect of oral administration of intact casein on gastrointestinal hormone secretion and pancreatic α-amylase activity in Korean native steer. Asian-Australas. J. Anim. Sci. 2013, 26, 654. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.B.; Lee, K.W.; Wang, T.; Lee, J.S.; Jung, U.S.; Nejad, J.G.; Oh, Y.K.; Baek, Y.C.; Kim, K.H.; Lee, H.G. Administration of encapsulated L-tryptophan improves duodenal starch digestion and increases gastrointestinal hormones secretions in beef cattle. Asian-Australas. J. Anim. Sci. 2020, 33, 91–99. [Google Scholar] [CrossRef]
- Trotta, R.J.; Lemley, C.O.; Vonnahme, K.A.; Swanson, K.C. Effects of nutrient restriction and melatonin supplementation from mid-to-late gestation on maternal and fetal small intestinal carbohydrase activities in sheep. Domest. Anim. Endocrinol. 2021, 74, 106555. [Google Scholar] [CrossRef] [PubMed]
- Trotta, R.J.; Keomanivong, F.E.; Peine, J.L.; Caton, J.S.; Swanson, K.C. Influence of maternal nutrient restriction and rumen-protected arginine supplementation on post-ruminal digestive enzyme activity of lamb offspring. Livest. Sci. 2020, 241. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.P.; Xu, M.; Yao, J.H.; Liu, K.; Li, F.; Liu, Y.; Wang, F.; Sun, F.F.; Liu, N.N. Regulation of pancreatic exocrine secretion in goats: Differential effects of short- and long-term duodenal phenylalanine treatment. J. Anim. Physiol. Anim. Nutr. 2013, 97, 431–438. [Google Scholar] [CrossRef]
- Yu, Z.P.; Xu, M.; Liu, K.; Yao, J.H.; Yu, H.X.; Wang, F. Leucine markedly regulates pancreatic exocrine secretion in goats. J. Anim. Physiol. Anim. Nutr. 2014, 98, 169–177. [Google Scholar] [CrossRef]
- Liu, K.; Liu, Y.; Liu, S.M.; Xu, M.; Yu, Z.P.; Wang, X.; Cao, Y.C.; Yao, J.H. Relationships between leucine and the pancreatic exocrine function for improving starch digestibility in ruminants. J. Dairy Sci. 2015, 98, 2576–2582. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.C.; Yang, X.J.; Guo, L.; Zheng, C.; Wang, D.D.; Cai, C.J.; Yao, J.H. Regulation of pancreas development and enzymatic gene expression by duodenal infusion of leucine and phenylalanine in dairy goats. Livest. Sci. 2018, 216, 9–15. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, S.; Yang, X.; Guo, L.; Cai, C.; Yao, J. Effects of dietary leucine and phenylalanine on gastrointestinal development and small intestinal enzyme activities in milk-fed Holstein dairy calves. Biosci. Rep. 2019, 39, BSR20181733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiners, J.N. Supplemental Leucine in Milk Replacer Fed to Neonatal Calves and Lambs: Effects on Growth, Development, and Metabolism. Ph.D. Thesis, North Dakota State University, Fargo, North Dakota, 2021. [Google Scholar]
- Liu, K.; Shen, J.; Cao, Y.; Cai, C.; Yao, J. Duodenal infusions of isoleucine influence pancreatic exocrine function in dairy heifers. Arch Anim. Nutr. 2018, 72, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Tian, H.; Shen, J.; Zheng, C.; Liu, S.; Cao, Y.; Cai, C.; Yao, J. Phenylalanine regulates initiation of digestive enzyme mRNA translation in pancreatic acinar cells and tissue segments in dairy calves. Biosci. Rep. 2018, 38, BSR20171189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Liang, Z.; Zheng, C.; Liu, B.; Yin, Q.; Cao, Y.; Yao, J. Leucine affects α-amylase synthesis through PI3K/Akt-mTOR signaling pathways in pancreatic acinar cells of dairy calves. J. Agric. Food. Chem. 2018, 66, 5149–5156. [Google Scholar] [CrossRef]
- Guo, L.; Yao, J.H.; Zheng, C.; Tian, H.B.; Liu, Y.L.; Liu, S.M.; Cai, C.J.; Xu, X.R.; Cao, Y.C. Leucine regulates α-amylase and trypsin synthesis in dairy calf pancreatic tissue in vitro via the mammalian target of rapamycin signalling pathway. Animal 2019, 13, 1899–1906. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liu, K.; Liu, S.; Guo, L.; Yao, J.; Cai, C. Leucine regulates the exocrine function in pancreatic tissue of dairy goats in vitro. BioMed Res. Int. 2019, 2019, 7521715. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, K.; Liu, S.; Guo, L.; Cai, C.; Yao, J. Isoleucine regulates the synthesis of pancreatic enzymes via the activation of mRNA expression and phosphorylation in the mammalian target of rapamycin signalling pathways in pancreatic tissues. BioMed Res. Int. 2019, 2019, 6302950. [Google Scholar] [CrossRef]
- Trotta, R.J.; Sitorski, L.G.; Acharya, S.; Brake, D.W.; Swanson, K.C. Duodenal infusions of starch with casein or glutamic acid influence pancreatic and small intestinal carbohydrase activities in cattle. J. Nutr. 2020, 150, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Tian, H.; Yao, J.; Ren, H.; Yin, Q.; Cao, Y. Leucine improves alpha-amylase secretion through the general secretory signaling pathway in pancreatic acinar cells of dairy calves. Am. J. Physiol. Cell Physiol. 2020, 318, C1284–C1293. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.C.; Matthews, J.C.; Woods, C.A.; Harmon, D.L. Influence of substrate and/or neurohormonal mimic on in vitro pancreatic enzyme release from calves postruminally infused with partially hydrolyzed starch and/or casein. J. Anim. Sci. 2003, 81, 1323–1331. [Google Scholar] [CrossRef]
- Swanson, K.C.; Benson, J.A.; Matthews, J.C.; Harmon, D.L. Pancreatic exocrine secretion and plasma concentration of some gastrointestinal hormones in response to abomasal infusion of starch hydrolyzate and/or casein. J. Anim. Sci. 2004, 82, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Branco, A.F.; Harmon, D.L.; Bohnert, D.W.; Larson, B.T.; Bauer, M.L. Estimating true digestibility of nonstructural carbohydrates in the small intestine of steers. J. Anim. Sci. 1999, 77, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.C.; Kelly, N.; Salim, H.; Wang, Y.J.; Holligan, S.; Fan, M.Z.; McBride, B.W. Pancreatic mass, cellularity, and α-amylase and trypsin activity in feedlot steers fed diets differing in crude protein concentration. J. Anim. Sci. 2008, 86, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.H.; Klusmeyer, T.H.; Cameron, M.R. Microbial protein synthesis and flows of nitrogen fractions to the duodenum of dairy cows. J. Dairy Sci. 1992, 75, 2304–2323. [Google Scholar] [CrossRef]
- Alman, M.S. Effect of site of starch infusion on nutrient utilization by growing beef steers consuming cubed alfalfa hay. Master’s Thesis, University of Kentucky, Lexington, KY, USA, 2009. [Google Scholar]
- Remillard, R.L.; Johnson, D.E.; Lewis, L.D.; Nockels, C.F. Starch digestion and digesta kinetics in the small intestine of steers fed on a maize grain and maize silage mixture. Anim. Feed Sci. Technol. 1990, 30, 79–89. [Google Scholar] [CrossRef]
- Robbers, K.; Westreicher-Kristen, E.; Troescher, A.; Susenbeth, A. Effects of abomasally infused amylase and increasing amounts of corn starch on fecal excretion of starch, total and microbial nitrogen, and volatile fatty acids in heifers. J. Anim. Sci. 2019, 97, 3984–3993. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.R.R.; Schoonmaker, J.P.; Chizzotti, M.L.; Teixeira, P.D.; Dias, J.C.O.; Gionbelli, T.R.S.; Rodrigues, A.C.; Costa, S.F.; Ladeira, M.M. Total nutrient digestibility and small intestine starch digestion in Nellore and Angus young bulls fed a whole shelled corn diet. J. Anim. Physiol. Anim. Nutr. 2019, 103, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, C.; Gerbert, C.; Frieten, D.; Dusel, G.; Eder, K.; Zitnan, R.; Hammon, H. Effects of ad libitum milk replacer feeding and butyrate supplementation on the epithelial growth and development of the gastrointestinal tract in Holstein calves. J. Dairy Sci. 2019, 102, 8513–8526. [Google Scholar] [CrossRef]
- Bauer, M.L.; Harmon, D.L.; McLeod, K.R.; Huntington, G.B. Influence of α-linked glucose on jejunal sodium–glucose co-transport activity in ruminants. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 129, 577–583. [Google Scholar] [CrossRef]
- Gilbert, M.S.; Pantophlet, A.J.; Berends, H.; Pluschke, A.M.; van den Borne, J.J.G.C.; Hendriks, W.H.; Schols, H.A.; Gerrits, W.J.J. Fermentation in the small intestine contributes substantially to intestinal starch disappearance in calves. J. Nutr. 2015, 145, 1147–1155. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, K.C.; Hazelton, S.R.; Matthews, J.C.; Swanson, K.C.; Harmon, D.L.; Branco, A.F. Influence of starch and casein administered postruminally on small intestinal sodium-glucose cotransport activity and expression. Braz. Arch. Biol. Technol. 2007, 50, 963–970. [Google Scholar] [CrossRef] [Green Version]
- Galand, G. Brush border membrane sucrase-isomaltase, maltase-glucoamylase and trehalase in mammals. Comparative development, effects of glucocorticoids, molecular mechanisms, and phylogenetic implications. Comp. Biochem. Physiol. B 1989, 94, 1–11. [Google Scholar] [CrossRef]
- Lin, A.H.M.; Nichols, B.L.; Quezada-Calvillo, R.; Avery, S.E.; Sim, L.; Rose, D.R.; Naim, H.Y.; Hamaker, B.R. Unexpected high digestion rate of cooked starch by the Ct-maltase-glucoamylase small intestine mucosal α-glucosidase subunit. PLoS ONE 2012, 7, e35473. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.W.; Chegeni, M.; Kim, K.H.; Zhang, G.; Benmoussa, M.; Quezada-Calvillo, R.; Nichols, B.L.; Hamaker, B.R. Different sucrose-isomaltase response of Caco-2 cells to glucose and maltose suggests dietary maltose sensing. J. Clin. Biochem. Nutr. 2014, 54, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Chegeni, M.; Amiri, M.; Nichols, B.L.; Naim, H.Y.; Hamaker, B.R. Dietary starch breakdown product sensing mobilizes and apically activates alpha-glucosidases in small intestinal enterocytes. FASEB J. 2018, 32, 3903–3911. [Google Scholar] [CrossRef] [Green Version]
- Huber, J.T.; Jacobson, N.L.; Allen, R.S.; Hartman, P.A. Digestive enzyme activities in the young calf. J. Dairy Sci. 1961, 44, 1494–1501. [Google Scholar] [CrossRef]
- Shirazi-Beechey, S.P.; Kemp, R.B.; Dyer, J.; Beechey, R.B. Changes in the functions of the intestinal brush border membrane during the development of the ruminant habit in lambs. Comp. Biochem. Physiol. B 1989, 94, 801–806. [Google Scholar] [CrossRef]
- Ridgway, S. A mini review of dolphin carbohydrate metabolism and suggestions for future research using exhaled air. Front. Endocrinol. 2013, 4, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretchmer, N.; Sunshine, P. Intestinal disaccharidase deficiency in the sea lion. Gastroenterology 1967, 53, 123–129. [Google Scholar] [CrossRef]
- Rowell-Schäfer, A.; Lechner-Doll, M.; Hofmann, R.R.; Streich, W.J.; Güven, B.; Meyer, H.H.D. Metabolic evidence of a ‘rumen bypass’ or a ‘ruminal escape’of nutrients in roe deer (Capreolus capreolus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 128, 289–298. [Google Scholar] [CrossRef]
- Kerry, K.R. Intestinal disaccharidase activity in a monotreme and eight species of marsupials (with an added note on the disaccharidases of five species of sea birds). Comp. Biochem. Physiol. 1969, 29, 1015–1022. [Google Scholar] [CrossRef]
- Mohamed, S.A.; Fahmy, A.S.; Salah, H.A. Disaccharidase activities in camel small intestine: Biochemical investigations of maltase–glucoamylase activity. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 124–130. [Google Scholar] [CrossRef]
- Naim, H.Y.; Heine, M.; Zimmer, K.P. Congenital sucrase-isomaltase deficiency: Heterogeneity of inheritance, trafficking, and function of an intestinal enzyme complex. J. Pediatr. Gastroenterol. Nutr. 2012, 55 (Suppl. 2), S13–S20. [Google Scholar] [CrossRef] [Green Version]
- Hunziker, W.; Spiess, M.; Semenza, G.; Lodish, H.F. The sucrase-isomaltase complex: Primary structure, membrane-orientation, and evolution of a stalked, intrinsic brush border protein. Cell 1986, 46, 227–234. [Google Scholar] [CrossRef]
- Fransen, J.A.; Hauri, H.P.; Ginsel, L.A.; Naim, H.Y. Naturally occurring mutations in intestinal sucrase-isomaltase provide evidence for the existence of an intracellular sorting signal in the isomaltase subunit. J. Cell Biol. 1991, 115, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfalah, M.; Jacob, R.; Preuss, U.; Zimmer, K.P.; Naim, H.; Naim, H.Y. O-linked glycans mediate apical sorting of human intestinal sucrase-isomaltase through association with lipid rafts. Curr. Biol. 1999, 9, 593–596. [Google Scholar] [CrossRef] [Green Version]
- Jacob, R.; Alfalah, M.; Grünberg, J.; Obendorf, M.; Naim, H.Y. Structural determinants required for apical sorting of an intestinal brush-border membrane protein. J. Biol. Chem. 2000, 275, 6566–6572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riby, J.E.; Kretchmer, N. Participation of pancreatic enzymes in the degradation of intestinal sucrase-isomaltase. J. Pediatr. Gastroenterol. Nutr. 1985, 4, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Naim, H.Y.; Joberty, G.; Alfalah, M.; Jacob, R. Temporal association of the N- and O-linked glycosylation events and their implication in the polarized sorting of intestinal brush border sucrase-isomaltase, aminopeptidase N, and dipeptidyl peptidase IV. J. Biol. Chem. 1999, 274, 17961–17967. [Google Scholar] [CrossRef] [Green Version]
- Dyer, J.; Merediz, E.F.-C.; Salmon, K.S.H.; Proudman, C.J.; Edwards, G.B.; Shirazi-Beechey, S.P. Molecular characterisation of carbohydrate digestion and absorption in equine small intestine. Equine Vet. J. 2002, 34, 349–358. [Google Scholar] [CrossRef]
- Ørskov, E.R.; Mayes, R.W.; Mann, S.O. Postruminal digestion of sucrose in sheep. Br. J. Nutr. 1972, 28, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.; Harmon, D. 505 Influence of abomasal sucrose infusion on small intestinal disaccharidase activity in lambs. J. Anim. Sci. 1997, 75, 263. [Google Scholar]
- Greene, H.L.; Stifel, F.B.; Herman, R.H. Dietary stimulation of sucrase in a patient with sucrase-isomaltase deficiency. Biochem. Med. 1972, 6, 409–418. [Google Scholar] [CrossRef]
- Nichols, B.L.; Avery, S.E.; Quezada-Calvillo, R.; Kilani, S.B.; Lin, A.H.; Burrin, D.G.; Hodges, B.E.; Chacko, S.K.; Opekun, A.R.; Hindawy, M.E.; et al. Improved starch digestion of sucrase-deficient shrews treated with oral glucoamylase enzyme supplements. J. Pediatr. Gastroenterol. Nutr. 2017, 65, e35–e42. [Google Scholar] [CrossRef]
- Lee, B.H.; Rose, D.R.; Lin, A.H.; Quezada-Calvillo, R.; Nichols, B.L.; Hamaker, B.R. Contribution of the individual small intestinal α-glucosidases to digestion of unusual α-linked glycemic disaccharides. J. Agric. Food. Chem. 2016, 64, 6487–6494. [Google Scholar] [CrossRef] [PubMed]
- Scharrer, E.; Liebich, H.G.; Raab, W.; Promberger, N. Influence of age and rumen development on intestinal absorption of galactose and glucose in lambs. A functional and morphological study. Zentralbl. Veterinarmed. A 1979, 26, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Scharrer, E.; Peter, W.; Raab, W. Reciprocal relationship between rumen development and intestinal sugar transport capacity in sheep. Zentralbl. Veterinarmed. A 1979, 26, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Shirazi-Beechey, S.P.; Hirayama, B.A.; Wang, Y.; Scott, D.; Smith, M.W.; Wright, E.M. Ontogenic development of lamb intestinal sodium-glucose co-transporter is regulated by diet. J. Physiol. 1991, 437, 699–708. [Google Scholar] [CrossRef]
- Dyer, J.; Scott, D.; Beechey, R.B.; Care, A.D.; Abbas, K.S.; Shirazi-Beechey, S.P. Dietary regulation of intestinal glucose transport. In Mammalian Brush Border Membrane Proteins, Part II; Lentze, M.J., Grand, R.J., Naim, H.Y., Eds.; Thieme Verlag: Stuttgart, Germany, 1994; pp. 65–72. [Google Scholar]
- Shirazi-Beechey, S.P.; Wood, I.S.; Dyer, J.; Scott, D.; King, T.P. Intestinal sugar transport in ruminants. In Ruminant Physiology: Digestion, Metabolism, Growth and Reproduction; Engelhardt, W.V., Leonhard, S., Breves, G., Giesecke, D., Eds.; Ferdinand Enke Verlag: Stuttgart, Germany, 1995; p. 117. [Google Scholar]
- Moran, A.W.; Al-Rammahi, M.; Zhang, C.; Bravo, D.; Calsamiglia, S.; Shirazi-Beechey, S.P. Sweet taste receptor expression in ruminant intestine and its activation by artificial sweeteners to regulate glucose absorption. J. Dairy Sci. 2014, 97, 4955–4972. [Google Scholar] [CrossRef] [Green Version]
- Moran, A.W.; Al-Rammahi, M.A.; Batchelor, D.J.; Bravo, D.M.; Shirazi-Beechey, S.P. Glucagon-like peptide-2 and the enteric nervous system are components of cell-cell communication pathway regulating intestinal Na +/glucose co-transport. Front. Nutr. 2018, 5, 101. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.F.; Harmon, D.L.; Vanzant, E.S.; McLeod, K.R.; Boling, J.A.; Matthews, J.C. The small intestinal epithelia of beef steers differentially express sugar transporter messenger ribonucleic acid in response to abomasal versus ruminal infusion of starch hydrolysate. J. Anim. Sci. 2010, 88, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Lohrenz, A.-K.; Duske, K.; Schönhusen, U.; Losand, B.; Seyfert, H.M.; Metges, C.C.; Hammon, H.M. Glucose transporters and enzymes related to glucose synthesis in small intestinal mucosa of mid-lactation dairy cows fed 2 levels of starch. J. Dairy Sci. 2011, 94, 4546–4555. [Google Scholar] [CrossRef]
- Klinger, S.; Zurich, M.; Schröder, B.; Breves, G. Effects of dietary starch source on electrophysiological intestinal epithelial properties and intestinal glucose uptake in growing goats. Arch. Anim. Nutr. 2013, 67, 289–300. [Google Scholar] [CrossRef]
- Ferraris, R.P.; Choe, J.Y.; Patel, C.R. Intestinal absorption of fructose. Annu. Rev. Nutr. 2018, 38, 41–67. [Google Scholar] [CrossRef]
- Shu, R.; David, E.S.; Ferraris, R.P. Dietary fructose enhances intestinal fructose transport and GLUT5 expression in weaning rats. Am. J. Physiol. 1997, 272, G446–G453. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Q.; Glimm, D.R.; Kennelly, J.J. Distribution of mammalian facilitative glucose transporter messenger RNA in bovine tissues. Int. J. Biochem. 1993, 25, 1897–1903. [Google Scholar] [CrossRef]
- Douard, V.; Ferraris, R.P. Regulation of the fructose transporter GLUT5 in health and disease. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E227–E237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, R.; David, E.S.; Ferraris, R.P. Luminal fructose modulates fructose transport and GLUT-5 expression in small intestine of weaning rats. Am. J. Physiol. 1998, 274, G232–G239. [Google Scholar] [CrossRef] [PubMed]
- David, E.S.; Cingari, D.S.; Ferraris, R.P. Dietary induction of intestinal fructose absorption in weaning rats. Pediatr. Res. 1995, 37, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Ferraris, R.P. Developmental reprogramming of rat GLUT-5 requires de novo mRNA and protein synthesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G113–G120. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; David, E.S.; Espina, N.; Ferraris, R.P. GLUT-5 expression in neonatal rats: Crypt-villus location and age-dependent regulation. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G666–G674. [Google Scholar] [CrossRef]
- Cui, X.L.; Soteropoulos, P.; Tolias, P.; Ferraris, R.P. Fructose-responsive genes in the small intestine of neonatal rats. Physiol. Genom. 2004, 18, 206–217. [Google Scholar] [CrossRef] [Green Version]
- Douard, V.; Choi, H.I.; Elshenawy, S.; Lagunoff, D.; Ferraris, R.P. Developmental reprogramming of rat GLUT5 requires glucocorticoid receptor translocation to the nucleus. J. Physiol. 2008, 586, 3657–3673. [Google Scholar] [CrossRef]
- Douard, V.; Cui, X.L.; Soteropoulos, P.; Ferraris, R.P. Dexamethasone sensitizes the neonatal intestine to fructose induction of intestinal fructose transporter (Slc2A5) function. Endocrinology 2008, 149, 409–423. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Douard, V.; Mochizuki, K.; Goda, T.; Ferraris, R.P. Diet-induced epigenetic regulation in vivo of the intestinal fructose transporter Glut5 during development of rat small intestine. Biochem. J. 2011, 435, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Kellett, G.L.; Helliwell, P.A. The diffusive component of intestinal glucose absorption is mediated by the glucose-induced recruitment of GLUT2 to the brush-border membrane. Biochem. J. 2000, 350, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff-Wagner, J.; Zitnan, R.; Schönhusen, U.; Pfannkuche, H.; Hudakova, M.; Metges, C.C.; Hammon, H.M. Diet effects on glucose absorption in the small intestine of neonatal calves: Importance of intestinal mucosal growth, lactase activity, and glucose transporters. J. Dairy Sci. 2014, 97, 6358–6369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinger, S.; Schroder, B.; Gemmer, A.; Reimers, J.; Breves, G.; Herrmann, J.; Wilkens, M.R. Gastrointestinal transport of calcium and glucose in lactating ewes. Physiol. Rep. 2016, 4, e12817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinhoff-Wagner, J.; Schönhusen, U.; Zitnan, R.; Hudakova, M.; Pfannkuche, H.; Hammon, H.M. Ontogenic changes of villus growth, lactase activity, and intestinal glucose transporters in preterm and term born calves with or without prolonged colostrum feeding. PLoS ONE 2015, 10, e0128154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röder, P.V.; Geillinger, K.E.; Zietek, T.S.; Thorens, B.; Koepsell, H.; Daniel, H. The role of SGLT1 and GLUT2 in intestinal glucose transport and sensing. PLoS ONE 2014, 9, e89977. [Google Scholar] [CrossRef] [PubMed]
- Pappenheimer, J.R.; Reiss, K.Z. Contribution of solvent drag through intercellular junctions to absorption of nutrients by the small intestine of the rat. J. Membr. Biol. 1987, 100, 123–136. [Google Scholar] [CrossRef]
- Krehbiel, C.R.; Britton, R.A.; Harmon, D.L.; Peters, J.P.; Stock, R.A.; Grotjan, H.E. Effects of varying levels of duodenal or midjejunal glucose and 2-deoxyglucose infusion on small intestinal disappearance and net portal glucose flux in steers. J. Anim. Sci. 1996, 74, 693–700. [Google Scholar] [CrossRef]
- Schambye, P. Volatile acids and glucose in portal blood of sheep. 2. Sheep fed hay and hay plus crushed oats. Nord. Vet. 1951, 3, 748–762. [Google Scholar]
- Mayes, R.W.; Ørskov, E.R. The utilization of gelled maize starch in the small intestine of sheep. Br. J. Nutr. 1974, 32, 143–153. [Google Scholar] [CrossRef]
- Franklin, M.A.; Mathew, A.G.; Vickers, J.R.; Clift, R.A. Characterization of microbial populations and volatile fatty acid concentrations in the jejunum, ileum, and cecum of pigs weaned at 17 vs 24 days of age. J. Anim. Sci. 2002, 80, 2904–2910. [Google Scholar] [CrossRef] [PubMed]
- Huntington, G.B.; Reynolds, P.J. Net absorption of glucose, L-lactate, volatile fatty acids, and nitrogenous compounds by bovine given abomasal infusions of starch or glucose. J. Dairy Sci. 1986, 69, 2428–2436. [Google Scholar] [CrossRef]
- Lapierre, H.; Lobley, G.E.; Doepel, L.; Raggio, G.; Rulquin, H.; Lemosquet, S. Triennial Lactation Symposium: Mammary metabolism of amino acids in dairy cows. J. Anim. Sci. 2012, 90, 1708–1721. [Google Scholar] [CrossRef] [PubMed]
- El-Kadi, S.W.; Baldwin, R.L.; McLeod, K.R.; Sunny, N.E.; Bequette, B.J. Glutamate is the major anaplerotic substrate in the tricarboxylic acid cycle of isolated rumen epithelial and duodenal mucosal cells from beef cattle. J. Nutr. 2009, 139, 869–875. [Google Scholar] [CrossRef] [Green Version]
- Oba, M.; Baldwin, R.L.; Bequette, B.J. Oxidation of glucose, glutamate, and glutamine by isolated ovine enterocytes in vitro is decreased by the presence of other metabolic fuels. J. Anim. Sci. 2004, 82, 479–486. [Google Scholar] [CrossRef]
- Kristensen, N.B.; Huntington, G.B.; Harmon, D.L. Splanchnic carbohydrate and energy metabolism in growing ruminants. In Biology of Metabolism in Growing Animals; Burrin, D.G., Mersmann, H., Eds.; Elsevier Ltd.: Edinburgh, UK, 2005; Volume 3, pp. 405–432. [Google Scholar]
- MacRae, J.C.; Armstrong, D.G. Studies on intestinal digestion in the sheep. 2. Digestion of some carbohydrate constituents in hay, cereal and hay-cereal rations. Br. J. Nutr. 1969, 23, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Beever, D.E.; Coleho da Silva, J.F.; Prescott, J.H.D.; Armstrong, D.G. The effect in sheep of physical form and stage of growth on the sites of digestion of a dried grass. 1. Sites of digestion of organic matter, energy and carbohydrate. Br. J. Nutr. 1972, 28, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB Life 2010, 62, 869–877. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Small Ruminants | ||||
| [163] | [164] | [134] | [74] | [170] | |
| Species | Lambs | Sheep | Sheep | Lambs | Goats |
| Diet | Pelleted | Roughage | 85% fescue hay | Wheat hay + cracked corn | Hay/Corn/Wheat |
| Source | Duodenal | Duodenal | Ruminal/abomasal | Abomasal | Dietary |
| Nutrient | Glucose | Glucose/Galactose/Fructose | PHS | Casein | - |
| Length | 4 d | 4 d | 7 d | 10 d | 21 d |
| Amount | 8.1 g/d | 8.1 g/d | 6 g/h | 35 g/d | 600 g/d |
| Transporter | SGLT1 | SGLT1 | SGLT1 | SGLT1 | SGLT1 |
| Parameter | Abundance, activity | Abundance, activity | Activity | Abundance, activity | Abundance, activity |
| Duodenum 1 | ↑Abundance and activity | ↑Abundance and activity | - | ↑Abundance, →activity | - |
| Jejunum 1 | - | - | ↑Activity | ↓Abundance, ↑activity | →Abundance, ↑activity |
| Ileum 1 | - | - | - | →Abundance, activity | - |
| Item | Cattle | ||||
| [134] | [18] | [136] | [168] | [104] | |
| Species | Steers | Steers | Steers | Steers | Calves |
| Diet | 85% fescue hay | 90% orchardgrass | 90% alfalfa cubes | Alfalfa cubes | Milk replacer |
| Source | Ruminal/abomasal | Abomasal | Abomasal | Ruminal/abomasal | Dietary |
| Nutrient | PHS | PHS or Glucose | PHS + Casein | PHS | Fructose |
| Length | 7 d | 35 d | 10 d | 14 d | 28 d |
| Amount | 40 g/h | 776 g PHS/d, 886 g glucose/d | 352 g/d starch, 52.8 g/d casein | 800 g/d | 92 g/d |
| Transporter | SGLT1 | SGLT1 | SGLT1 | SGLT1/GLUT2/GLUT5 | SGLT1/GLUT2/GLUT5 |
| Parameter | Activity | Abundance, activity | Abundance, activity | mRNA expression | mRNA expression |
| Duodenum 1 | - | → | → | ↑SGLT1 (RS) | → |
| Jejunum 1 | ↑Activity | → | → | → | → |
| Ileum 1 | - | → | → | ↑SGLT1, GLUT2 (AS) | → |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trotta, R.J.; Harmon, D.L.; Matthews, J.C.; Swanson, K.C. Nutritional and Physiological Constraints Contributing to Limitations in Small Intestinal Starch Digestion and Glucose Absorption in Ruminants. Ruminants 2022, 2, 1-26. https://doi.org/10.3390/ruminants2010001
Trotta RJ, Harmon DL, Matthews JC, Swanson KC. Nutritional and Physiological Constraints Contributing to Limitations in Small Intestinal Starch Digestion and Glucose Absorption in Ruminants. Ruminants. 2022; 2(1):1-26. https://doi.org/10.3390/ruminants2010001
Chicago/Turabian StyleTrotta, Ronald J., David L. Harmon, James C. Matthews, and Kendall C. Swanson. 2022. "Nutritional and Physiological Constraints Contributing to Limitations in Small Intestinal Starch Digestion and Glucose Absorption in Ruminants" Ruminants 2, no. 1: 1-26. https://doi.org/10.3390/ruminants2010001