1. Introduction
Vitamin A and its derivatives (retinoids) are known to promote and maintain normal differentiation in various types of cells and to inhibit malignant proliferation, suggesting their potential involvement in the anti-tumor effects of these compounds [
1,
2]. Among them, all-
trans-3,7,11,15-tetramethyl-2,6,10,14-hexadecatetraenoic acid (geranylgeranoic acid; GGA) and its didehydrogenated derivative all-
trans-3,7,11,15-tetramethyl-2,4,6,10,14-hexadecapentaenoic acid (dGGA or E-5166) are referred to as acyclic retinoids due to their binding affinities to cellular retinoic acid-binding protein (CRABP) [
3] and ligand activities to nuclear retinoic acid receptors (RAR and RXR) [
4]. In particular, the acyclic retinoid dGGA has been developed as a prophylactic agent against secondary hepatocellular carcinoma with higher therapeutic efficacy and lower toxicity, which enables the compound to be administered with large doses for the long term [
5]. Indeed, we had observed the effects of dGGA on human hepatoma-derived cell lines and chemically induced liver tumors in rats, reporting them as similar to the differentiation-inducing effects of retinoic acid [
6,
7].
To begin with, dGGA was developed as a substance with a strong affinity for CRABP. CRABP is an intracellular transporter for retinoic acid which appears and increases during carcinogenesis and cell proliferation phases and is abundant in tumor tissues [
6,
7]. Additionally, compounds like dGGA, which have an affinity for CRABP, are observed to be taken up more in hepatocarcinoma tissues than in non-cancerous tissues [
6,
7]. Therefore, we predicted that a compound with anti-tumor activity and strong affinity for CRABP would be preferentially taken up by cancerous tissues via CRABP, leading to the manifestation of anti-tumor effects.
More than 40 years ago, Omori and Chytil [
8] observed that relatively small amounts of retinoids administered orally to vitamin A-deficient rats induced rapid changes in gene expression, revealing that one of the mechanisms of vitamin A action involves gene expression. In that study, a single oral dose of 100µg of retinoid (all-
trans retinoic acid, retinyl acetate, or dGGA) was administered to vitamin A-deficient rats, resulting in significant changes in gene expression profiles appearing within 1 h after administration, many of which persisted for at least 3 months. From this, we obtained three implications for our cancer prevention research using retinoids. First, this study on the tumor-suppressive effects of a single small dose of dGGA was designed and initiated based on the inference that similar changes in gene expression may occur at sites closely related to cell redifferentiation or induction of other mechanisms detrimental to malignant cells. Secondly, and more importantly, some of the changes in gene expression that occurred at that time persisted for as long as 3 months. Therefore, we hypothesized that a single oral administration of a small dose of dGGA to young C3H mice, which spontaneously develop hepatocarcinoma, could suppress the spontaneous development of liver cancer in mice at age two years. Finally, we also decided to examine in the present experimental design whether these changes occur only after a single administration of a small amount (50 µg/mouse) of dGGA and are not observed when large doses of dGGA were continuously administered over a longer period. In other words, in the present study, we decided to compare the hepatoma-suppressive effect of dGGA between a single small dose of dGGA at different times of administration and long-term rearing of mice for more than one year on a diet supplemented with 0.02% dGGA (equivalent to 625 µg per mouse per day).
In this study, we discovered a novel method of administering a hepatoma suppressant that is distinctly different from conventional methods with larger doses for longer periods. It is a low-frequency administration method that demonstrates a very potent anti-tumor effect with relatively small amounts (50 µg per mouse) of dGGA administered orally only 1–2 times to 11-month-old male C3H/HeNCrj mice that naturally develop liver cancer.
2. Materials and Methods
dGGA (E-5166) was prepared by Eisai Co., Ltd. (Tokyo, Japan). Male C3H/HeNCrj mice were purchased from Charles River Japan Inc., Kanagawa, Japan, and fed on a basal pelleted laboratory diet (MF; Oriental Yeast Co., Ltd., Tokyo, Japan) and distilled water ad libitum.
Experiment 1: Inhibition of Spontaneous Hepatocarcinogenesis through Long-term Continuous Administration of dGGA: A total of 20 male CH3/HeNCrj mice at 8 weeks of age were raised on a basal diet. From 32 to 91 weeks of age, the mice were provided with a diet containing 0.02% dGGA. Preparation of dGGA-containing diets: A basal pelleted laboratory diet was soaked in a solution of dGGA dissolved in ethyl alcohol (adjusted to 0.02% of the diet) for 5 min, then the solution was evaporated completely and dried thoroughly. Eisai Co., Ltd. (Gifu, Japan) was responsible for the preparation of the diet.
Experiment 2: Inhibition of Spontaneous Hepatocarcinogenesis through a Single and Small-Dose Administration of dGGA: A total of 120 mice, which were 8 weeks of age, were divided into eight groups of 10 to 25 animals. Using a gastric applicator syringe, 100 μL of cottonseed oil with 50 µg dGGA dissolved in it was orally administered to the animals only once at different time points: 2 (Group 1), 5 (Group 2), 6 (Group 3), 8 (Group 4), 11 (Group 5), 14 (Group 6), and 17 months (Group 7) after birth, respectively. The control animals (Group 8) were not treated with dGGA during the experimental period.
Experiment 3: Inhibition of Spontaneous Hepatocarcinogenesis through Small-Dose Dual Administrations of dGGA: Similarly, to the previous setup, mice under the same conditions were administered 50 µg of dGGA twice at different time points. The administration timings for each group were as follows: Some of the animals in Group 2 of Experiment 2 received a total of two dGGA doses at months 5 and 11, which was designated Group 2*, and some members of Group 4 received a total of two dGGA doses at months 8 and 11, which was designated Group 4*, respectively.
At the age of 23 months, a laparotomy of all mice was performed to evaluate the numbers and sizes of liver tumors, and after careful removal of the tumor portion, liver tumors were individually weighed (separation from the non-cancerous portion is relatively easy). The liver tumor tissues were processed by the routine gross method.
All the animals in the current experiment were kept in the animal room of the Gifu University School of Medicine, which is a hygienically controlled facility, after obtaining permission from the Research Ethics Review Committee of Gifu University School of Medicine (1986-34).
The following methods were used to analyze the statistical data. Number of liver tumors, weight, and tumor-bearing rate: Welch’s t-test. Test for differences in the distribution pattern of liver tumors: Ridit analysis.
3. Results
3.1. Influences of dGGA Administration on the Body Weight of the Mice
The body weights of the mice in each group at the final time point were as follows:
Experiment (1): 33.5 ± 3.1 g (n = 20). Experiment (2): Group 1: 33.6 ± 0.9 g (n = 10), Group 2: 34.4 ± 2.3 g (n = 9), Group 3: 33.4 ± 2.1 g (n = 11), Group 4: 34.1 ± 2.3 g (n = 10), Group 5: 34.5 ± 1.5 g (n = 11), Group 6: 33.8 ± 2.8 g (n = 10), Group 7: 33.4 ± 2.0 g (n = 10), and Group 8 (Control): 34.9 ± 4.7 g (n = 14). Experiment (3): Group 2*: 33.1 ± 1.9 g (n = 12) and Group 4*: 33.9 ± 2.4 g (n = 10). There were no significant differences between groups, including the long-term administration group. dGGA toxicity and side effects have been studied previously [
5,
6], and the doses used in this study, including the long-term administration cases, were all less than the toxicity and side-effect dose.
3.2. Inhibition of Hepatocarcinogenesis through Long-Term Continuous Administration of dGGA: (Experiment 1)
Using feed containing 0.02% dGGA, the administration of dGGA was conducted for 448 days. Based on the intake of feed, the estimated daily dGGA administration was 625 μg per mouse. At the time of laparotomy, the average tumor weight per mouse was 0.52 ± 0.85 g, and the tumor-bearing rate (proportion of animals with liver tumors) was 60.0%, both of which showed significant differences (
p < 0.05) compared to the control group (1.52 ± 1.48 g per mouse, 100%), respectively (
Table 1).
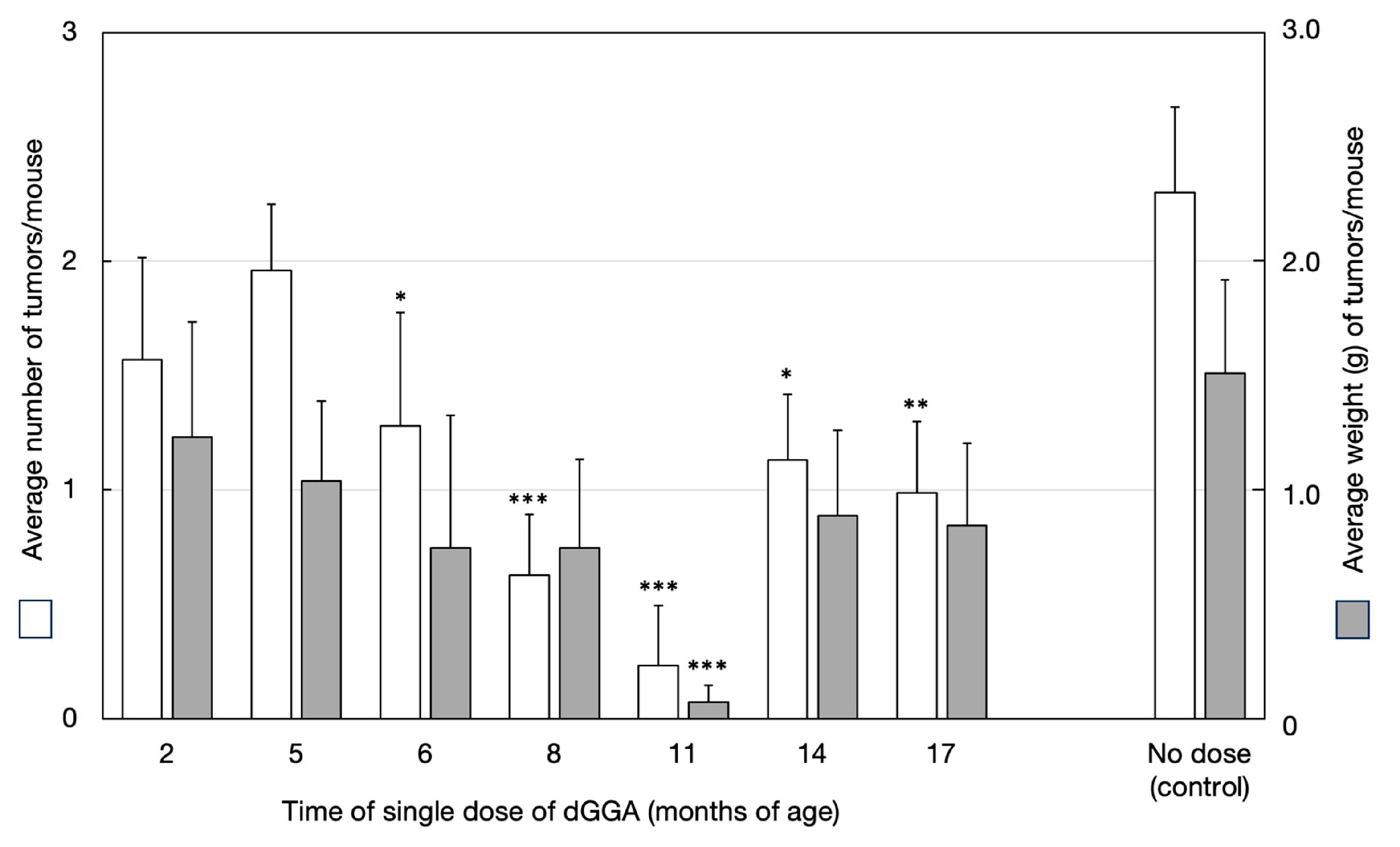
3.3. Inhibition of Spontaneous Hepatocarcinogenesis through Small-Dose Single Administration of dGGA: (Experiment 2) (Figure 1)
The cumulative average number of tumors per mouse and the average tumor weight (g per mouse) for each group of animals are shown in
Figure 1. The average number of tumors per mouse was significantly lower (
p < 0.05) in the dGGA-administered group (Groups 1–7: 0.81 ± 0.91 tumors per mouse) compared to the non-administered control group (2.33 ± 1.23 tumors per mouse). Groups 4–7 showed a significantly lower number of tumors (
p < 0.02–0.05) compared to the control group, with the lowest count observed in Group 5 (0.43 ± 0.79 tumors per mouse).
The average tumor weight in the dGGA-administered group (Groups 1–7: 0.59 ± 1.13 g per mouse) was significantly lower (
p < 0.05) than that of the control group (1.52 ± 1.48 g per mouse). Among the individual groups, all groups (1–7) exhibited lower values compared to the control group, with a significant difference (
p < 0.02) observed especially in Group 5 (administered dGGA only once, at 11 months of age) (
Figure 1).
The tumor-bearing rate was 100% for Group 2 (administered dGGA at 5 months of age) and the non-administered control Group 8, with liver tumors in all cases. In contrast, the administered groups at 2 months and 6–17 months showed lower tumor incidence rates, particularly in the groups dosed at 6, 8, 11, and 17 months, with significant differences (p < 0.02–0.05) compared to the control group. Notably, the lowest rate was 28.6% for Group 5, which received dGGA administration at 11 months.
The distribution of the number of tumors per mouse revealed that, except for Groups 1 and 2, the dGGA-administered groups had significantly fewer tumors per mouse (according to Ridit analysis). Especially in Groups 3, 4, 5, and 7, the most frequent number of tumors per mouse was 0, and the maximum count was 2. In the control group, animals with 2 tumors were the most common, with a maximum count of 4 per mouse.
Regarding the distribution of tumor sizes, tumors with a diameter of 5 mm or less were the most common, followed by those with a diameter of 10–20 mm. In the control group, more than half of the animals had tumors with a diameter of 20 mm or more, showing a significant difference from the dGGA-administered groups. Especially in Groups 4 and 5, no tumors with a diameter of 20 mm or more were observed, with a significant difference (p < 0.01) from the control group.
Figure 1.
Relationship between timing of a single dose of dGGA and inhibitory effect on tumor development in spontaneous hepatoma mice (C3H/HeNCrj). *: p < 0.05, **: p < 0.02, ***: p < 0.01 vs. corresponding value of control.
Figure 1.
Relationship between timing of a single dose of dGGA and inhibitory effect on tumor development in spontaneous hepatoma mice (C3H/HeNCrj). *: p < 0.05, **: p < 0.02, ***: p < 0.01 vs. corresponding value of control.
3.4. Inhibition of Spontaneous Hepatocarcinogenesis through Small-Dose Dual Administration of dGGA: (Experiment 3) (Table 1 and Table 2)
After the first administration, a second administration of small-dose dGGA was conducted at intervals of 3–6 months, resulting in a more remarkable inhibitory effect on liver cancer. A significant difference (p < 0.01~0.001) was observed between the dual administration group and the single administration group. Groups that received dGGA administration once at 5 months and 11 months showed that the average number of tumors per mouse, and average tumor weights per mouse, were reduced to 1/8 and 1/16, respectively, compared to the single administration group. The most significant effect was observed in the group that received dGGA administration once at 8 months and 11 months (a total of two administrations), where tumor occurrence was almost absent, and the average tumor weight per mouse was only 1/760 of the control group’s value.
The differences in administration duration, dosage, and effects between the long-term high-dose administration method and the low-dose infrequent administration method are summarized in
Table 1 and
Table 2.
Table 2.
Inhibitory effects of timely dosing of dGGA on the development of spontaneous hepatoma in C3H/HeNCrj mice.
Table 2.
Inhibitory effects of timely dosing of dGGA on the development of spontaneous hepatoma in C3H/HeNCrj mice.
| Timing 1 | dGGA Amount (µg) | Tumor Incidence (%) | Tumor Number Per Mouse | Tumor Weight (g) Per Mouse |
|---|
| 5 M 1 | 50 | 100 ††† | 2.00 ± 0.71 ††† | 1.05 ± 0.86 ††† |
| 5 M + 11 M | 100 | 12.5 ****,††† | 0.25 ± 0.70 ****,††† | 0.04 ± 0.11***,††† |
| 8 M | 50 | 37.5 *** | 0.85 ± 1.69 *** | 0.75 ± 0.96 |
| 8 M + 11 M | 100 | 16.7 **** | 0.18 ± 0.40 ****,† | 0.002 ± 0.004****,†† |
| 11 M | 50 | 28.6 *** | 0.43 ± 0.79 ***,† | 0.08 ± 0.21***,†† |
| control | 0 | 100 | 2.33 ± 1.23 | 1.52 ± 1.48 |
4. Discussion
Administering 625 μg of dGGA per day for approximately 450 days resulted in significant differences in inhibitory effects on hepatocarcinogenesis, such as reducing the average tumor weight per mouse to approximately one-third of the control group. In contrast, by administering a much smaller amount of dGGA (approximately 1/3000 to 1/6000 of the total dose) only once or twice, a substantially superior anti-tumor effect was achieved compared to the conventional method (
Table 1).
Notably, the administration timing played a crucial role in the anti-tumor effects, with the most effective period being at 11 months of age. The natural incidence of liver tumors in C3H mice rapidly increases around one year of age [
9,
10,
11,
12,
13]. In the present study, groups that received dGGA administration during the low-incidence period of 2 to 5 months of age showed minimal anti-tumor effects. On the other hand, the group that received administration at 11 months of age, just before the expected increase in liver tumor incidence, exhibited the most significant effect. However, the effectiveness declined from 14 months of age onwards. In the group that received dGGA administration at 14 months, where cancer had likely already developed to some extent, although the average number of tumors per mouse was significantly lower (
Figure 1), the number of tumors with a diameter of 10 mm or more was not significantly different from the control group. This suggests that in this case, dGGA might have primarily suppressed the emergence of new tumors.
The group that received dual administration of dGGA at 11 months and 3–6 months earlier exhibited the most potent anti-tumor effect. This remarkable outcome was particularly evident in the group that received dual administration at 8 months and 11 months, where tumor occurrence was almost non-existent. These results may indicate that dGGA administered at the right time and in the right amount effectively prevents the development of cancer. Further investigation is needed to elucidate the most effective dose and time points for the administration of dGGA.
The results of the present study provide highly suggestive insights into the phase III clinical trials of dGGA [
5,
14,
15,
16]. Specifically, notable findings were observed in a randomized controlled trial with a placebo for the prevention of recurrence of secondary hepatocellular carcinoma after curative treatment for primary liver cancer [
5]. In addition to significant inhibitory effects on liver carcinogenesis, it is noteworthy that the preventive effects of second primary liver cancer development continued for more than three years after the end of the one-year dosing period. In these types of studies, it is quite common that the differences between the medication and non-medication (placebo) groups rapidly decreased or disappeared soon after medication cessation, even when significant inhibitory effects were observed during medication. Moreover, it is exceptionally rare for the inhibitory effect to persist for over six years after treatment cessation, as observed in subsequent follow-up observations [
14]. Therefore, the persistence of inhibitory effects after medication cessation is exceptional. These effects cannot be explained by the pharmacological effects of high-dose, long-term administration but can be understood by considering that an appropriate amount of dGGA was administered at the right time during recurrence.
This study was initially conducted based on an entirely novel concept. The observation that administering small amounts of physiologically active chemical substances only once or twice induced significant and long-lasting changes in the carcinogenic process, ultimately leading to the suppression of carcinogenesis through an unknown mechanism, is unprecedented to our knowledge. Apart from the prevention of cervical cancer by human papillomavirus (HPV) vaccines (mentioned later), such observations have scarcely been reported. Therefore, it would be extremely difficult to show the previous studies which support our findings. In fact, we announced the preliminary results of this study around 1990 [
7]. Unfortunately, we were unable to explain the mechanism underlying the preventive action of a single dose of dGGA, so we were unable to publish the results as a widespread academic paper. Recently, at least part of the mechanism has started to become apparent (Tabata, Omori, and Shidoji) [
17], and we are truly pleased that our research has come to light. Thus, in this context, we consider dGGA as an analog of endogenous metabolite GGA rather than an exogenous retinoid and we will discuss it in comparison with the effects of GGA.
As mentioned above, in C3H mice, the spontaneous occurrence of liver cancer begins between 11 and 12 months of age, and at 12 months of age, 60–75% of mice are anatomically observed to have developed liver tumors [
9,
10,
11,
12,
13]. While it is not entirely clear what is happening in the livers of mice at this time, we revealed that GGA is a substance synthesized via the mevalonate pathway in vivo [
18] and that its hepatic levels decrease with aging [
17]. Interestingly, in C3H mice, the period of decreased endogenous GGA levels in the liver almost coincided with the period when spontaneous hepatocarcinogenesis began to occur and a small dose of exogenously administered GGA exerts a high inhibitory effect on liver tumors. Details of our hypothesis regarding the significant inhibitory effect of a single administration of GGA at this age are seen in our recent study [
17]. In the present study, although we used dGGA instead of GGA, the effect is completely reproduced. Furthermore, taking into account the decay curve of liver GGA levels with aging [
17], it is interesting to note that the effect is further enhanced when small doses of dGGA are orally administered twice at 5 and 11 months of age, or 8 and 11 months, respectively. The excellent anti-carcinogenic effect through administering a small amount of dGGA or GGA at 11 months of age is seemingly explained by our hypothesis: “Administered GGA or dGGA at the timing just before cells in a precancerous state become cancerous remove precancerous cells [
19] and prevent the proliferation of cancer cells.” For details on the mechanism of removal of precancerous hepatoma cells by GGA, please refer to the latest review on GGA [
19]. The next challenge should be in discovering the critical period (the critical time point) when GGAs exert their most significant effects on other cancers naturally occurring in humans.
As detailed in the Introduction, the dose of dGGA used in this study, 50 µg/mouse per dose, was determined from our experience with many of our retinoid studies [
3,
4,
6,
8]. Assuming uniform diffusion throughout the 35 mL volume of a 35 g mouse, with the body density assumed to be 1, the concentration of dGGA, administered at 50 μg to the mouse, is calculated to be approximately 5 μM. Interestingly, in in vitro cell culture experiments using human liver cancer cell lines, dGGA has been shown to induce cell death at concentrations ranging from 1 to 10 μM [
20]. While this may be a coincidental correlation, the observed anti-hepatocarcinogenic effects in C3H mice may suggest that 50 μg of dGGA could be an appropriate dosage. It is also worthwhile comparing the single or double dose of 50 µg used in C3H mice with the dosages employed in phase III clinical trials of dGGA. In a randomized clinical trial (RCT) of dGGA using a placebo, 600 mg of dGGA was orally administered daily per person for one year after curative therapy for hepatoma [
5,
14], and in a separate RCT dGGA, 600 mg, 300 mg, or placebo was orally administered daily per person for 96 weeks [
15,
16]. In both RCTs, a significant inhibitory effect on the second primary hepatoma and a life-extension effect was confirmed in the 600 mg dose group. On the other hand, in the liver cancer prevention experiment using C3H mice conducted in this study, a preventive effect against spontaneous hepatoma was observed in the groups that received single or double oral administrations of 50 µg/mouse of dGGA at 11 months of age and in the group that ingested the equivalent of 625 µg/mouse from the diet for 448 days. When converted to the Human Equivalent Dose (HED) considering body surface area from these two dosages, following the 2005 FDA guidance (Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers.
https://www.fda.gov/regulatory-information/search-fda-guidance-documents/estimating-maximum-safe-starting-dose-initial-clinical-trials-therapeutics-adult-healthy-volunteers, accessed on 17 October 2023), they amount to 9.6–9.8 mg/60 kg and 120–122 mg/60 kg, respectively. These HED values are much lower compared to the daily dosage of 600 mg/person used in the two clinical trials described above. Of course, it is currently unclear whether 50 µg dGGA is the optimal dose for the prevention of hepatocarcinoma in male C3H mice. Future experiments should be conducted to determine the optimal dose and frequency of dGGA administration.
The mechanism behind the prolonged retention of changes in gene expression may involve DNA methylation. Differences in DNA methylation were observed between the cancerous and non-cancerous parts of the liver in C3H mice [
21], and in C3HHeB/Fej mice, which were introduced with the methylated repair enzyme MGMT (O6-methylguanine-DNA methyltransferase), spontaneous hepatocellular carcinoma was significantly suppressed [
22]. If we can selectively induce and secure DNA methylation on DNA that is directly or indirectly related to carcinogenesis inhibition, it is likely that our knowledge of cancer prevention and treatment will expand further. In the context of epigenetic changes spanning across cell generations, modifications such as methylation and acetylation of histones also play a crucial role. GGA is known to non-competitively inhibit lysine demethylase 1A (KDM1A or LSD1), one of the epigenetic enzymes [
23]. Additionally, treatment of human hepatoma-derived cell lines with 20 µM GGA has been reported to induce the translocation of KDM1A from the nuclear chromatin to the cytoplasm within 3 h [
24]. While it has not been confirmed whether dGGA exhibits similar effects as GGA, there is a possibility that changes in histone modifications may be involved in the mechanism of irreversible alterations induced by a single administration of GGA or dGGA in the progression of spontaneous hepatoma in C3H mice.
A most popular example of a clear effect of single-dose administration is the significant suppression of human cervical cancer by administering the HPV vaccine in a single dose [
25,
26]. Although this result seems to be based on a completely different mechanism than the effects observed in this study, future progress in the effective prevention of carcinogenesis with compounds like dGGA would emphasize the effectiveness of a single administration. Another suggestive example of the irreversible effects of a single administration appears in the study by Takahashi K. et al. [
27] that reported the effects of a single administration of anti-USAG-1 (uterine sensitization-associated gene-1) antibody-induced tooth regeneration. They hypothesized that the
USAG-1 gene plays key regulatory roles in suppressing tooth development and the administration of anti-USAG-1 relieves congenital tooth agenesis. These effects, similar to our findings, suggest the involvement of the irreversible changes induced by a single administration of signaling substances. The present study might have found the conditions necessary for such an “unlocking” response.
Although the details of the mechanism of action of the remarkable anti-tumor effect of GGAs demonstrated by the special administration method in this study are still largely unknown, we hope that this study will open a new dimension in the prevention and treatment of human cancer.




{kind=link}