
Transcriptomic Analysis of Insulin-Secreting Murine Hepatocytes Transduced with an Integrating Adeno-Associated Viral Vector



Abstract
:1. Introduction
2. Materials and Methods
2.1. Liver Samples
2.2. RNA Isolation and Sample Quality Analysis
2.3. Total RNA Sequencing (RNA-Seq) Analysis
2.4. Differential Gene Expression Analysis
2.5. Pathway Analysis
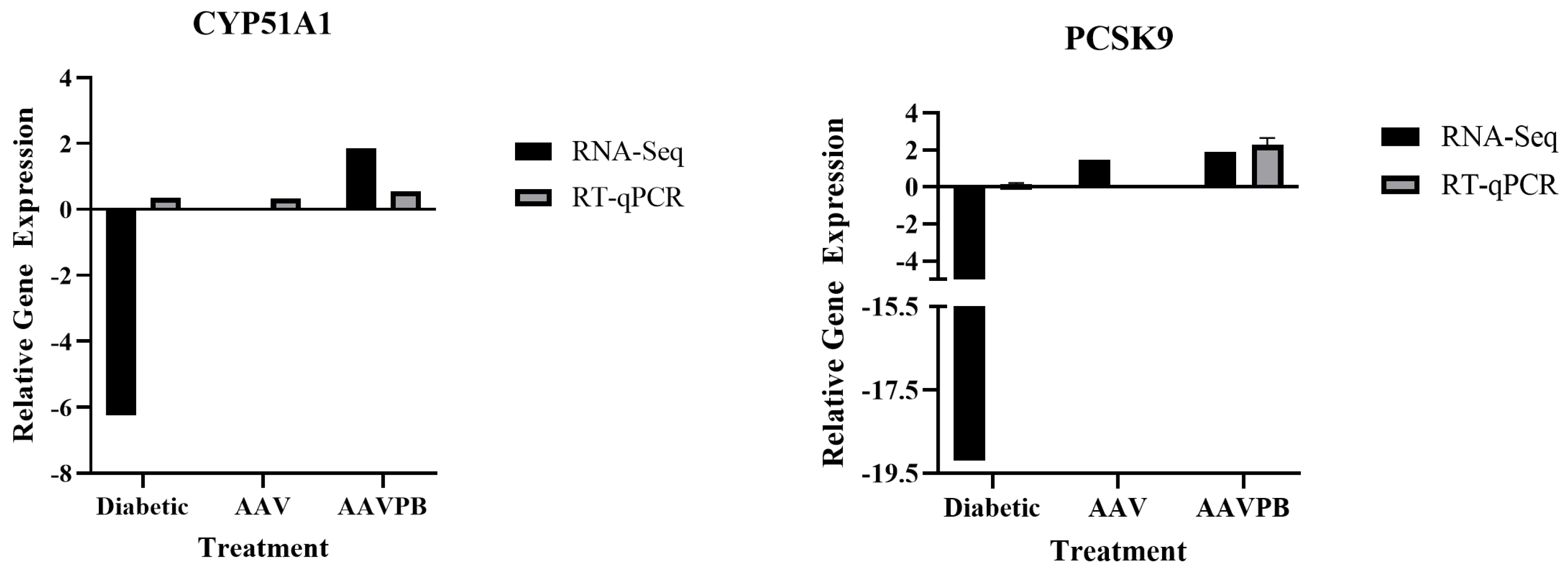
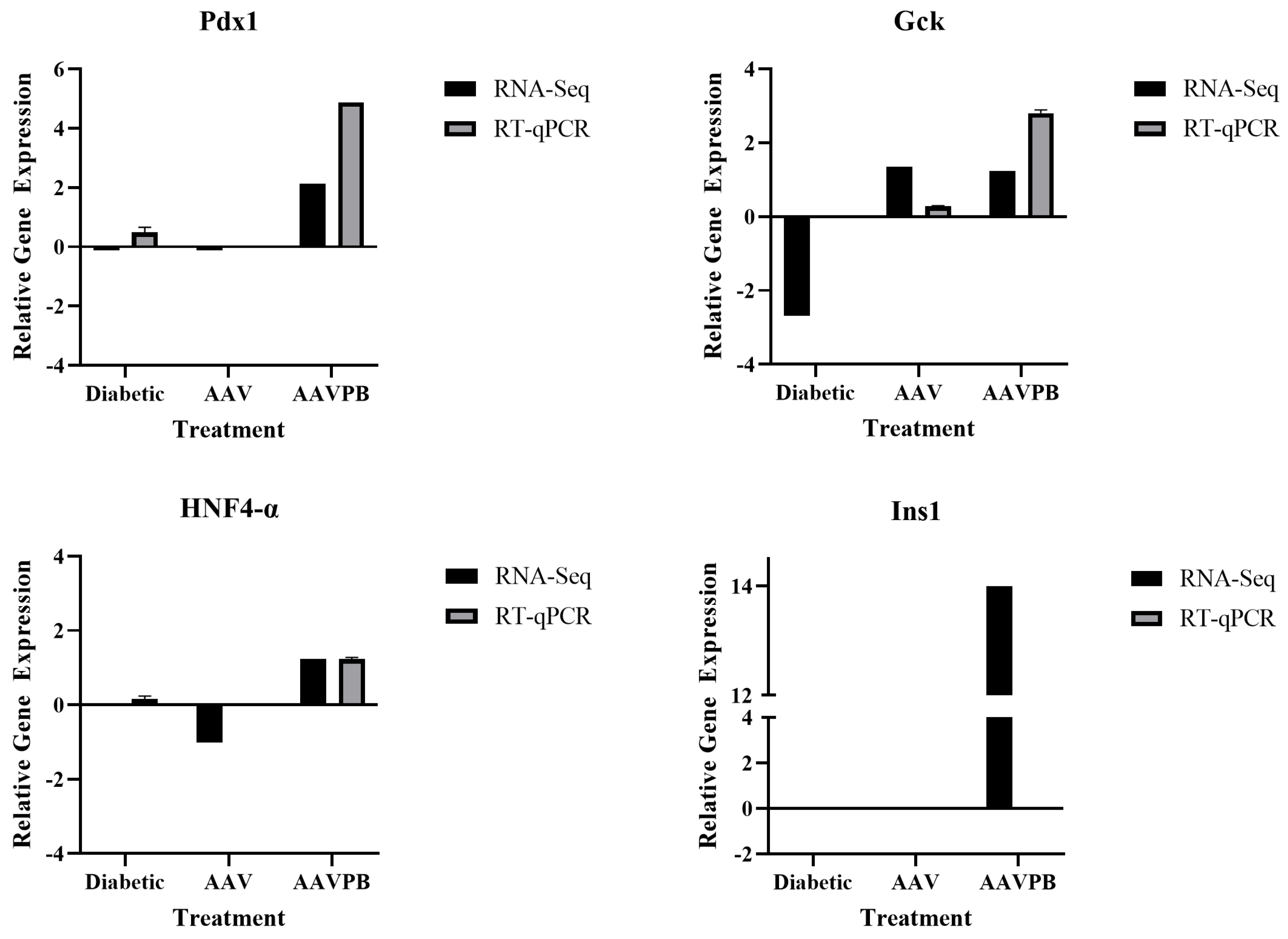
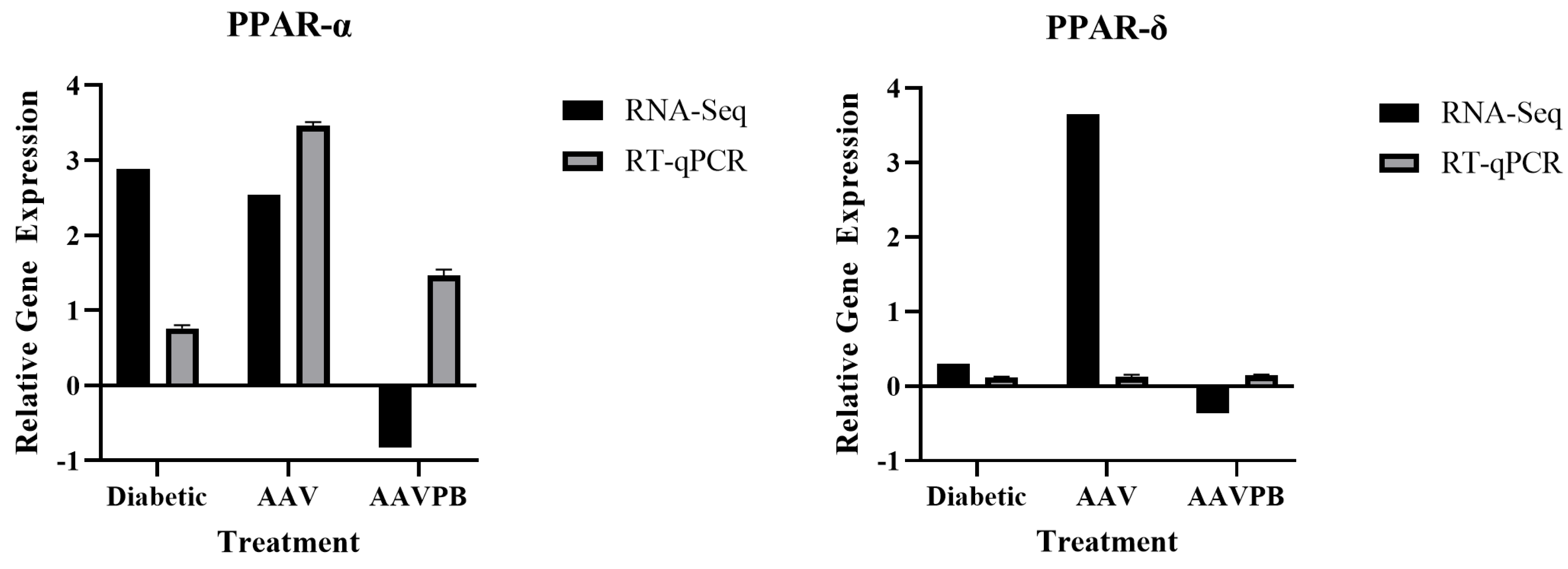
2.6. Validation of Differential Gene Expression by RT-qPCR
2.7. Statistical Analysis
3. Results
3.1. RNA Extraction and RNA-Seq Quality Analysis
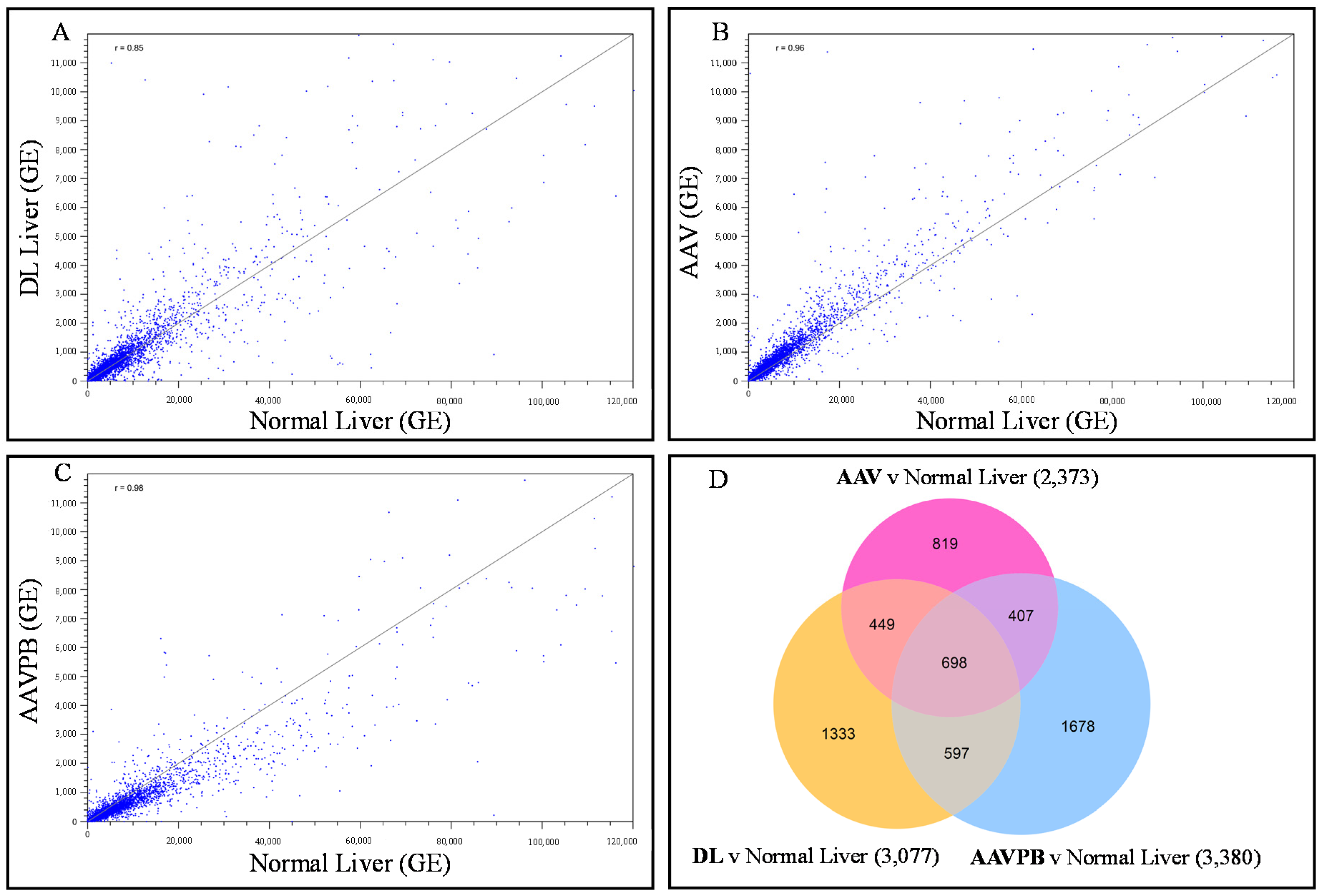
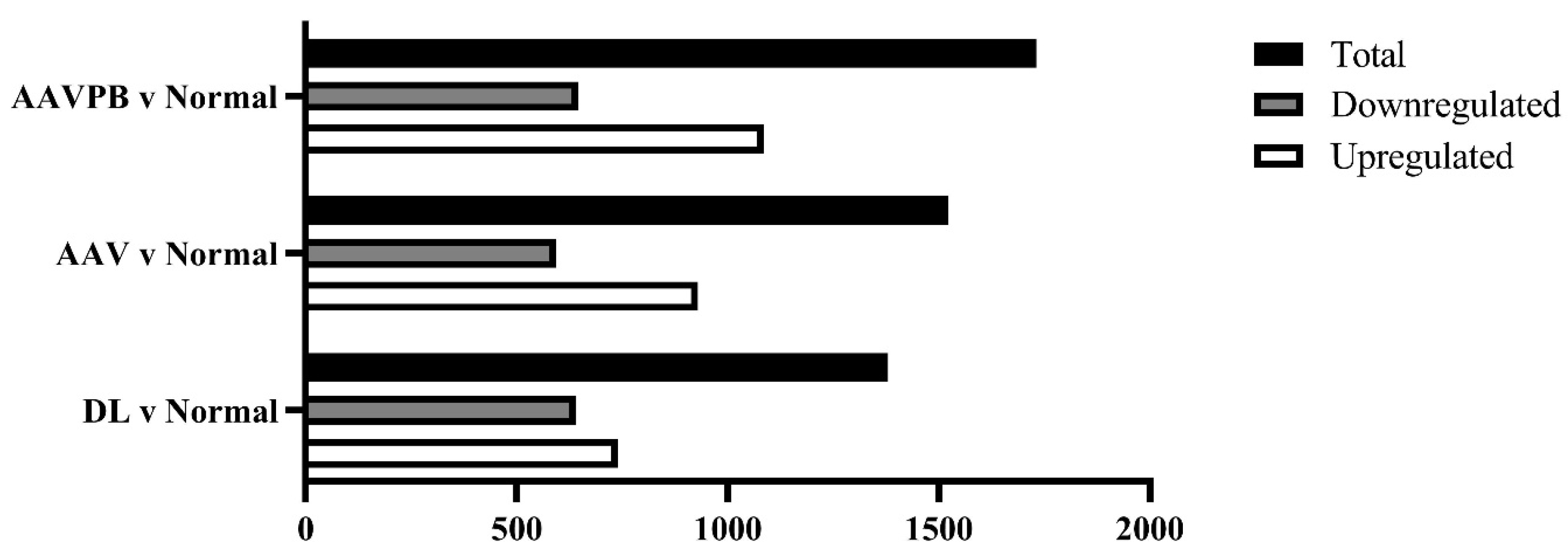
3.2. Identification and Visualization of Differentially Expressed Genes across Treatments
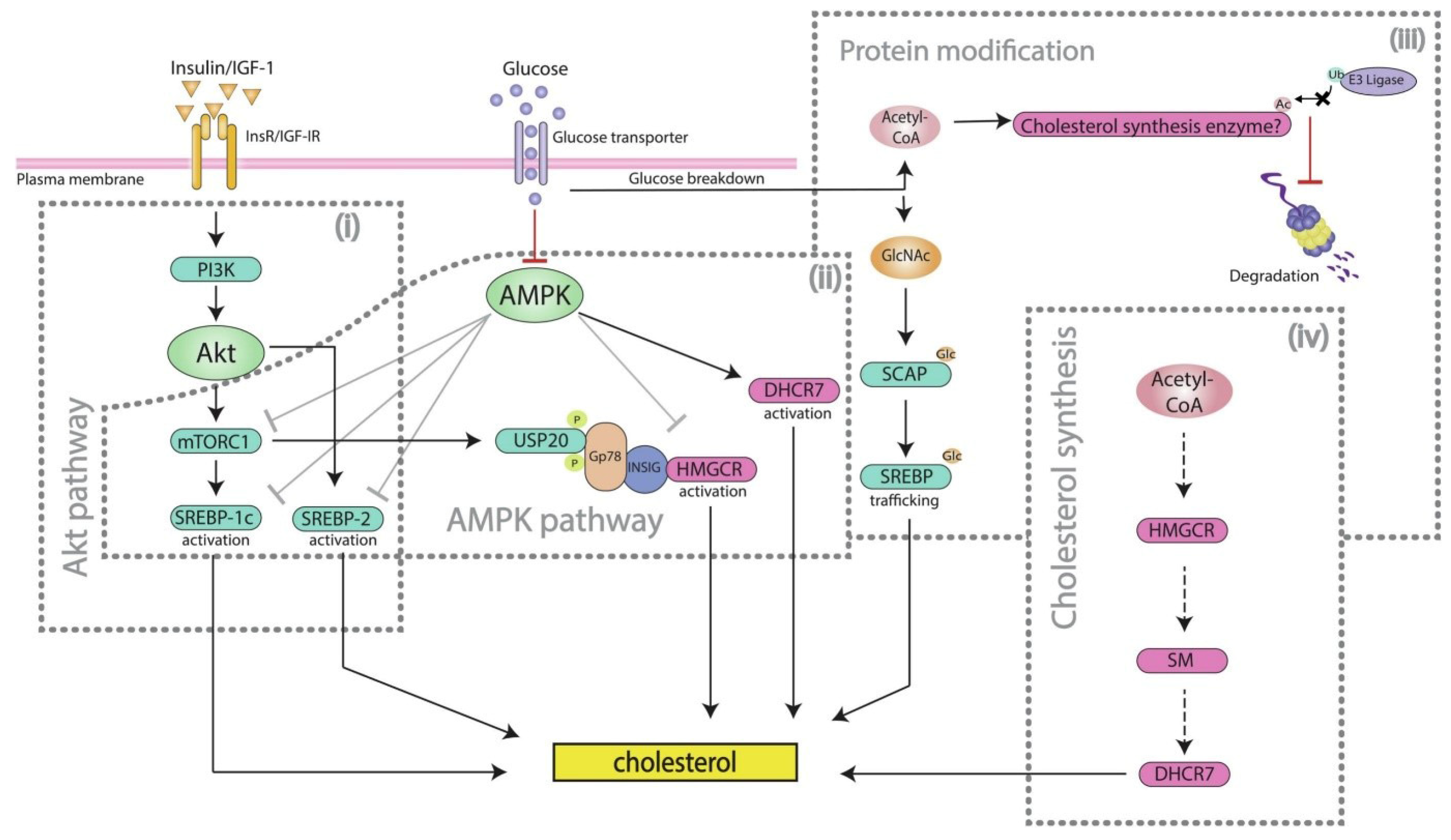
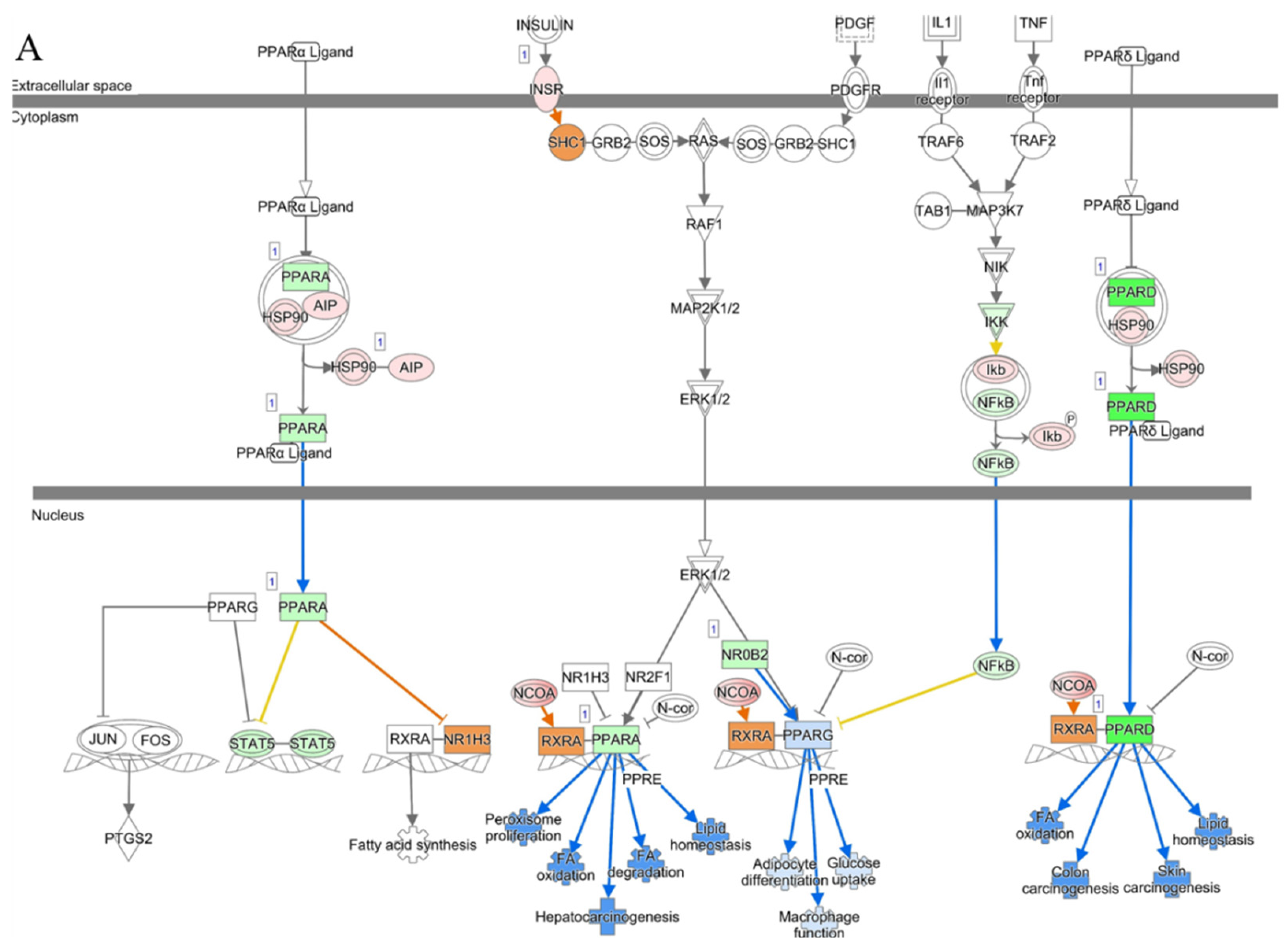
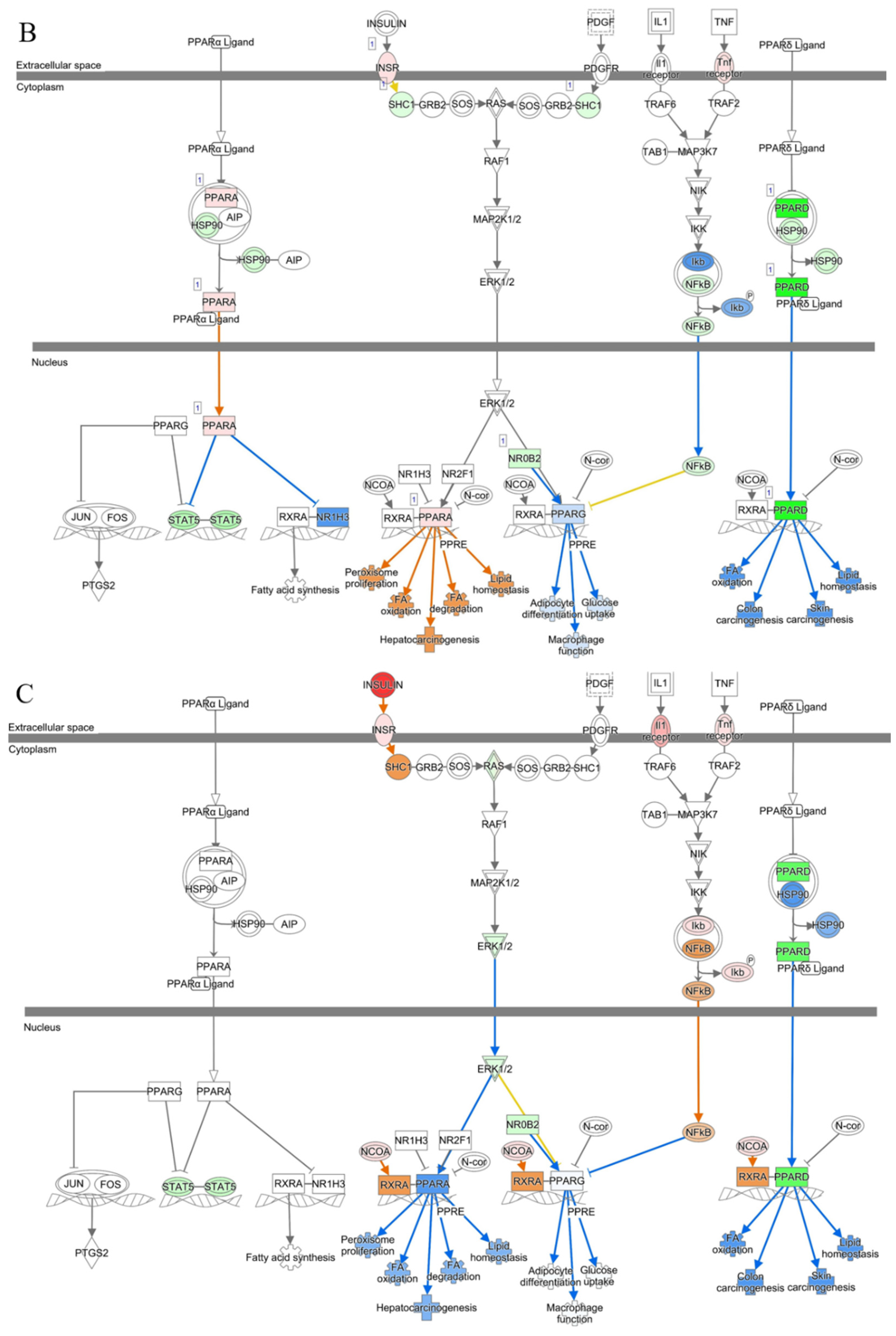
3.3. Canonical Pathway Analysis Identifies the Pathway of Cholesterol Biosynthesis in Diabetic and Piggybac Transduced Liver
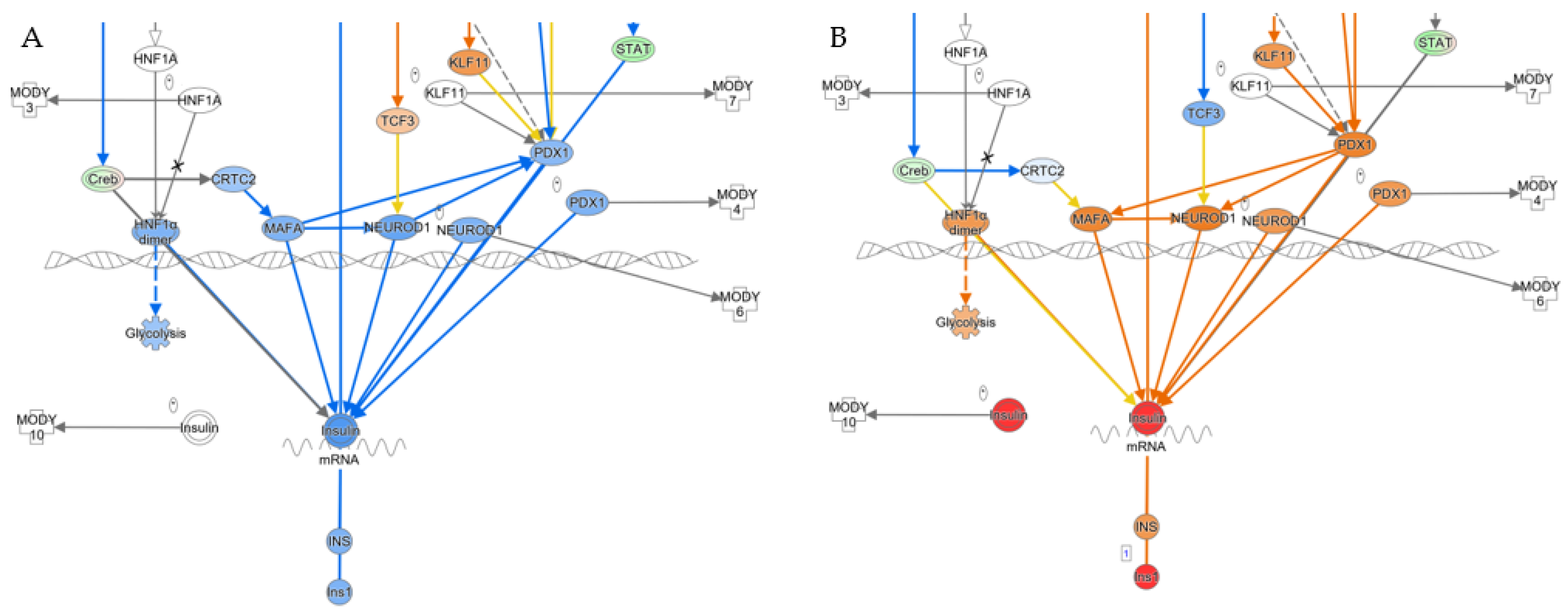
3.4. Pancreatic Markers Are Upregulated in AAV8/piggyBac-INS-FUR Transduced Liver and Downregulated in Diabetic Liver
3.5. Liver Function Is Maintained in AAV8/piggyBac-INS-FUR Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| AAV | Adeno-Associated Viral Vector-INS-FUR transduced liver |
| AAV8 | Adeno-Associated Viral Vector Serotype 8 treatment |
| AAVPB | Adeno-Associated Viral Vector/piggyBac-INS-FUR transduced liver |
| CYP51A1 | Cytochrome P450 Family 51 Subfamily A Member 1 |
| DEG | Differentially Expressed Gene |
| DHCR7 | 7-Dehydrocholesterol Reductase |
| DL | Diabetic liver |
| FFO | Full flow occlusion |
| Gck | glucokinase |
| GE | Gene Expression |
| HMGCR | 3-Hydroxy-3-Methylglutaryl-CoA Reductase |
| HNF4-α | Hepatocyte nuclear factor 4 alpha |
| Ins1 | Endogenous mouse insulin |
| INS-FUR | Furin cleavable insulin gene |
| IPA | Ingenuity Pathway Analysis |
| NOD | Non-obese diabetic |
| PCSK9 | Proprotein convertase subtilisin/kexin type 9 |
| Pdx1 | Pancreatic and duodenal homeobox 1 |
| piggyBac | Adeno-Associated Viral Vector/piggyBac-INS-FUR treatment |
| PPAR-α | Peroxisome proliferator-activated receptor-alpha |
| PPAR-δ | Peroxisome proliferator-activated receptor-delta |
| RNA-Seq | RNA Sequencing Data |
| T1D | Type 1 Diabetes |
References
- Federation, I.D. Type 1 Diabetes. Available online: https://idf.org/about-diabetes/type-1-diabetes/ (accessed on 23 August 2023).
- Atkinson, M.A.; Maclaren, N.K. The pathogenesis of insulin-dependent diabetes mellitus. N. Engl. J. Med. 1994, 331, 1428–1436. [Google Scholar] [PubMed]
- Hovorka, R. Closed-loop insulin delivery: From bench to clinical practice. Nat. Rev. Endocrinol. 2011, 7, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.A.; Lakey, J.R.; Paty, B.W.; Imes, S.; Korbutt, G.S.; Kneteman, N.M.; Bigam, D.; Rajotte, R.V.; Shapiro, A.J. Successful islet transplantation: Continued insulin reserve provides long-term glycemic control. Diabetes 2002, 51, 2148–2157. [Google Scholar] [CrossRef] [PubMed]
- Nathwani, A.C.; Reiss, U.M.; Tuddenham, E.G.; Rosales, C.; Chowdary, P.; McIntosh, J.; Della Peruta, M.; Lheriteau, E.; Patel, N.; Raj, D. Long-term safety and efficacy of factor IX gene therapy in hemophilia B. N. Engl. J. Med. 2014, 371, 1994–2004. [Google Scholar] [CrossRef]
- Cavazzana-Calvo, M.; Hacein-Bey, S.; Basile, G.v.d.S.; Gross, F.; Yvon, E.; Nusbaum, P.; Selz, F.; Hue, C.; Certain, S.; Casanova, J.-L. Gene therapy of human severe combined immunodeficiency (SCID)-X1 disease. Science 2000, 288, 669–672. [Google Scholar] [CrossRef]
- Acland, G.M.; Aguirre, G.D.; Ray, J.; Zhang, Q.; Aleman, T.S.; Cideciyan, A.V.; Pearce-Kelling, S.E.; Anand, V.; Zeng, Y.; Maguire, A.M. Gene therapy restores vision in a canine model of childhood blindness. Nat. Genet. 2001, 28, 92–95. [Google Scholar] [CrossRef]
- Ramamoorth, M.; Narvekar, A. Non viral vectors in gene therapy-an overview. J. Clin. Diagn. Res. JCDR 2015, 9, GE01. [Google Scholar] [CrossRef]
- Zhang, Y.; Chirmule, N.; Gao, G.-p.; Qian, R.; Croyle, M.; Joshi, B.; Tazelaar, J.; Wilson, J.M. Acute cytokine response to systemic adenoviral vectors in mice is mediated by dendritic cells and macrophages. Mol. Ther. 2001, 3, 697–707. [Google Scholar] [CrossRef]
- Mahoney, A.L.; Nassif, N.T.; O’Brien, B.A.; Simpson, A.M. Viral vectors in gene therapy and clinical applications. In Molecular Cloning; IntechOpen: Rijeka, Croatia, 2022. [Google Scholar] [CrossRef]
- Thorens, B. GLUT2, glucose sensing and glucose homeostasis. Diabetologia 2015, 58, 221–232. [Google Scholar] [CrossRef]
- Ren, B.; O’Brien, B.A.; Byrne, M.R.; Ch’ng, E.; Gatt, P.N.; Swan, M.A.; Nassif, N.T.; Wei, M.Q.; Gijsbers, R.; Debyser, Z. Long-term reversal of diabetes in non-obese diabetic mice by liver-directed gene therapy. J. Gene Med. 2013, 15, 28–41. [Google Scholar] [CrossRef]
- La, Q.T.; Ren, B.; Logan, G.J.; Cunningham, S.C.; Khandekar, N.; Nassif, N.T.; O’Brien, B.A.; Alexander, I.E.; Simpson, A.M. Use of a hybrid adeno-associated viral vector transposon system to deliver the insulin gene to diabetic NOD mice. Cells 2020, 9, 2227. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-T.; Jiang, F.-X.; Shi, P.; Zhang, T.; Liu, X.-Y.; Lin, X.-W.; San, Z.-Y.; Pang, X.-N. In vitro reprogramming of rat bmMSCs into pancreatic endocrine-like cells. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wan, J.; Guo, Y.; Zhu, S.; Wang, Y.; Wang, L.; Guo, Q.; Lu, Y.; Wang, Z. Transcriptome Analysis of Induced Pluripotent Stem Cell (iPSC)-derived Pancreatic β-like Cell Differentiation. Cell Transplant. 2017, 26, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Fenton, N.M.; Nguyen, T.B.; Sharpe, L.J.; Brown, A.J. Refining sugar’s involvement in cholesterol synthesis. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2022, 1868, 159266. [Google Scholar] [CrossRef]
- Dubois, V.; Eeckhoute, J.; Lefebvre, P.; Staels, B. Distinct but complementary contributions of PPAR isotypes to energy homeostasis. J. Clin. Investig. 2017, 127, 1202–1214. [Google Scholar] [CrossRef]
- Brown, A.J.; Coates, H.W.; Sharpe, L.J. Cholesterol synthesis. In Biochemistry of Lipids, Lipoproteins and Membranes; Elsevier: Amsterdam, The Netherlands, 2021; pp. 317–355. [Google Scholar]
- Motaln, H.; Wagner, K.D.; Debeljak, N.; Rassoulzadegan, M.; Ačimovič, J.; Rozman, D.; Horvat, S. Mouse knockout of the cholesterogenic cytochrome P450 lanosterol 14α-demethylase (Cyp51) resembles Antley-Bixler syndrome. J. Biol. Chem. 2011, 286, 29086–29097. [Google Scholar]
- Wang, Y.; Rogers, P.M.; Su, C.; Varga, G.; Stayrook, K.R.; Burris, T.P. Regulation of cholesterologenesis by the oxysterol receptor, LXRα. J. Biol. Chem. 2008, 283, 26332–26339. [Google Scholar] [CrossRef]
- Da Dalt, L.; Ruscica, M.; Bonacina, F.; Balzarotti, G.; Dhyani, A.; Di Cairano, E.; Baragetti, A.; Arnaboldi, L.; De Metrio, S.; Pellegatta, F. PCSK9 deficiency reduces insulin secretion and promotes glucose intolerance: The role of the low-density lipoprotein receptor. Eur. Heart J. 2019, 40, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; McCabe, T.; Condra, J.H.; Ni, Y.G.; Peterson, L.B.; Wang, W.; Strack, A.M.; Wang, F.; Pandit, S.; Hammond, H. An anti-PCSK9 antibody reduces LDL-cholesterol on top of a statin and suppresses hepatocyte SREBP-regulated genes. Int. J. Biol. Sci. 2012, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Edwards, P.A.; Muroya, H.; Gould, R.G. In vivo demonstration of the circadian rhythm of cholesterol biosynthesis inthe liver and intestine of the rat. J. Lipid Res. 1972, 13, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, A.V.; Luu, W.; Li, D.; Sharpe, L.J.; Brown, A.J. DHCR7: A vital enzyme switch between cholesterol and vitamin D production. Prog. Lipid Res. 2016, 64, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.F.; Price, T.B.; Bergeron, R. Regulation of net hepatic glycogenolysis and gluconeogenesis during exercise: Impact of type 1 diabetes. J. Clin. Endocrinol. Metab. 2004, 89, 4656–4664. [Google Scholar] [CrossRef]
- Leonard, J.; Peers, B.; Johnson, T.; Ferreri, K.; Lee, S.; Montminy, M. Characterization of somatostatin transactivating factor-1, a novel homeobox factor that stimulates somatostatin expression in pancreatic islet cells. Mol. Endocrinol. 1993, 7, 1275–1283. [Google Scholar]
- Hansen, S.K.; Párrizas, M.; Jensen, M.L.; Pruhova, S.; Ek, J.; Boj, S.F.; Johansen, A.; Maestro, M.A.; Rivera, F.; Eiberg, H. Genetic evidence that HNF-1α–dependent transcriptional control of HNF-4α is essential for human pancreatic β cell function. J. Clin. Investig. 2002, 110, 827–833. [Google Scholar] [CrossRef]
- Evans, R.M. The steroid and thyroid hormone receptor superfamily. Science 1988, 240, 889–895. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835. [Google Scholar] [CrossRef]
- Patsouris, D.; Mandard, S.; Voshol, P.J.; Escher, P.; Tan, N.S.; Havekes, L.M.; Koenig, W.; März, W.; Tafuri, S.; Wahli, W. PPARα governs glycerol metabolism. J. Clin. Investig. 2004, 114, 94–103. [Google Scholar] [CrossRef]
- Matsuoka, T.-A.; Kawashima, S.; Miyatsuka, T.; Sasaki, S.; Shimo, N.; Katakami, N.; Kawamori, D.; Takebe, S.; Herrera, P.L.; Kaneto, H. Mafa enables Pdx1 to effectively convert pancreatic islet progenitors and committed islet α-cells into β-cells in vivo. Diabetes 2017, 66, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-P.; Thorel, F.; Boyer, D.F.; Herrera, P.L.; Wright, C.V. Context-specific α-to-β-cell reprogramming by forced Pdx1 expression. Genes Dev. 2011, 25, 1680–1685. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Guo, P.; Shiota, C.; Zhang, T.; Coudriet, G.M.; Fischbach, S.; Prasadan, K.; Fusco, J.; Ramachandran, S.; Witkowski, P. Endogenous reprogramming of alpha cells into beta cells, induced by viral gene therapy, reverses autoimmune diabetes. Cell Stem Cell 2018, 22, 78–90.e74. [Google Scholar] [CrossRef]
- Akinci, E.; Banga, A.; Tungatt, K.; Segal, J.; Eberhard, D.; Dutton, J.R.; Slack, J.M. Reprogramming of various cell types to a beta-like state by Pdx1, Ngn3 and MafA. PLoS ONE 2013, 8, e82424. [Google Scholar] [CrossRef]
- Nagasaki, H.; Katsumata, T.; Oishi, H.; Tai, P.-H.; Sekiguchi, Y.; Koshida, R.; Jung, Y.; Kudo, T.; Takahashi, S. Generation of insulin-producing cells from the mouse liver using β cell-related gene transfer including Mafa and Mafb. PLoS ONE 2014, 9, e113022. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway or Molecule | Activation Z-Score * | ||
|---|---|---|---|
| DL v Normal | AAV v Normal | AAVPB v Normal | |
| Cholesterol Biosynthesis I | −3.32 | 0.38 | 2.71 |
| Cholesterol Biosynthesis II | −3.32 | 0.38 | 2.71 |
| Cholesterol Biosynthesis III | −3.32 | 0.38 | 2.71 |
| Gluconeogenesis | 2.71 | 2.33 | −2.45 |
| DHCR7 | −5.34 | N/A | 1.54 |
| HMGCR | −4.66 | 2.46 | 2.76 |
| Akt | −2.28 | 0.64 | 1.62 |
| Pathway | Analysis | Activation Z-Score | p-Value of Overlap |
|---|---|---|---|
| Insulin Secretion Pathway | AAVPB v Normal | 0.90 | <0.05 |
| AAV v Normal | 1.57 | <0.05 | |
| DL v Normal | 1.00 | >0.05 | |
| Beta cell v others | 1.54 | <0.05 |
| Pathway | Analysis | Activation Z-Score | p-Value of Overlap |
|---|---|---|---|
| PPAR Signaling | AAVPB v Normal | 0 | <0.05 |
| AAV v Normal | 1.51 | >0.05 | |
| DL v Normal | 0.28 | <0.05 | |
| Beta cell v others | −0.63 | >0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahoney, A.L.G.; Joshua, S.; Nassif, N.T.; Simpson, A.M. Transcriptomic Analysis of Insulin-Secreting Murine Hepatocytes Transduced with an Integrating Adeno-Associated Viral Vector. Int. J. Transl. Med. 2023, 3, 374-388. https://doi.org/10.3390/ijtm3030026
Mahoney ALG, Joshua S, Nassif NT, Simpson AM. Transcriptomic Analysis of Insulin-Secreting Murine Hepatocytes Transduced with an Integrating Adeno-Associated Viral Vector. International Journal of Translational Medicine. 2023; 3(3):374-388. https://doi.org/10.3390/ijtm3030026
Chicago/Turabian StyleMahoney, Alexandra L. G., Sergio Joshua, Najah T. Nassif, and Ann M. Simpson. 2023. "Transcriptomic Analysis of Insulin-Secreting Murine Hepatocytes Transduced with an Integrating Adeno-Associated Viral Vector" International Journal of Translational Medicine 3, no. 3: 374-388. https://doi.org/10.3390/ijtm3030026