

Nucleic Acid Vaccines for COVID-19: A Paradigm Shift in the Vaccine Development Arena


Abstract
:1. Introduction
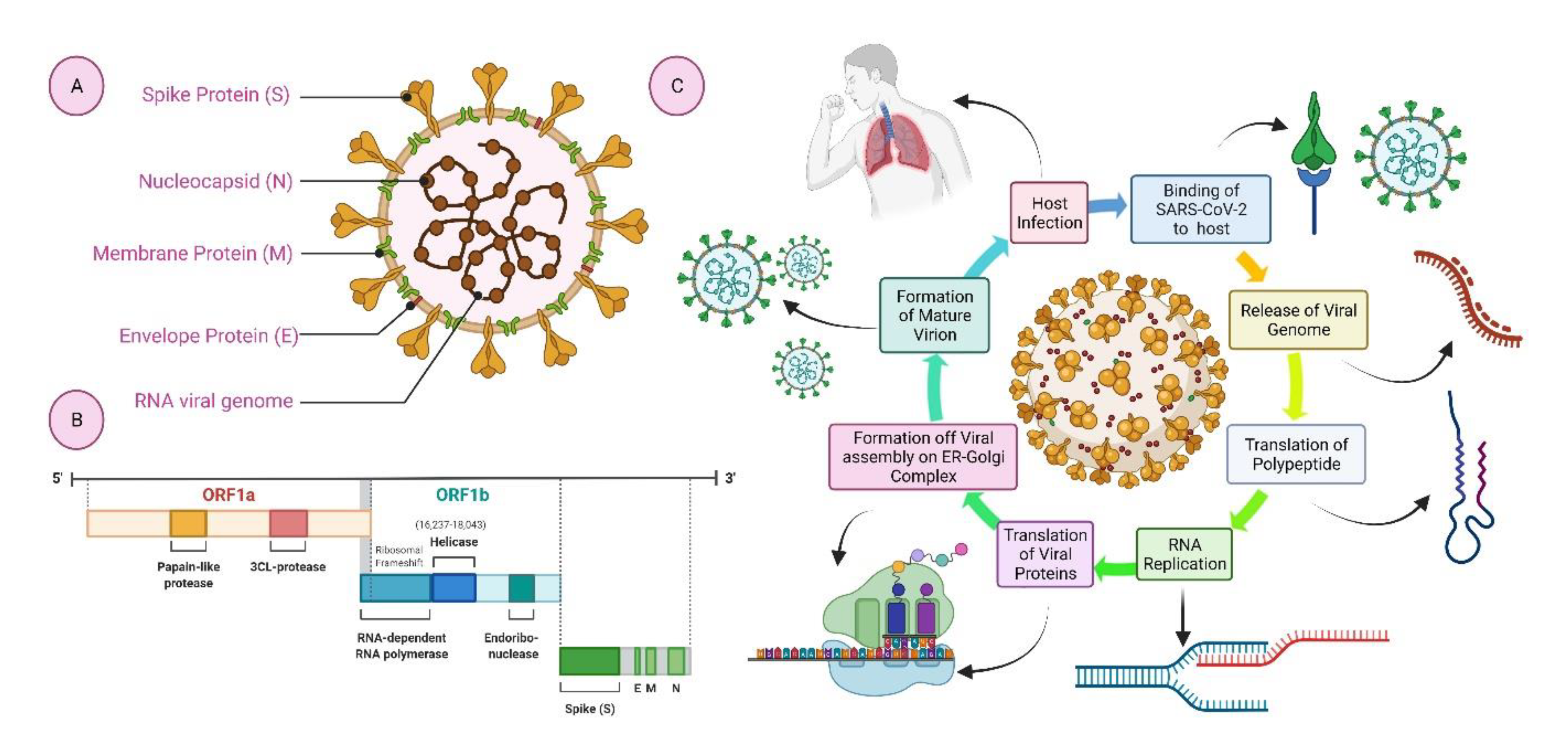
2. Structural Elements of COVID-19
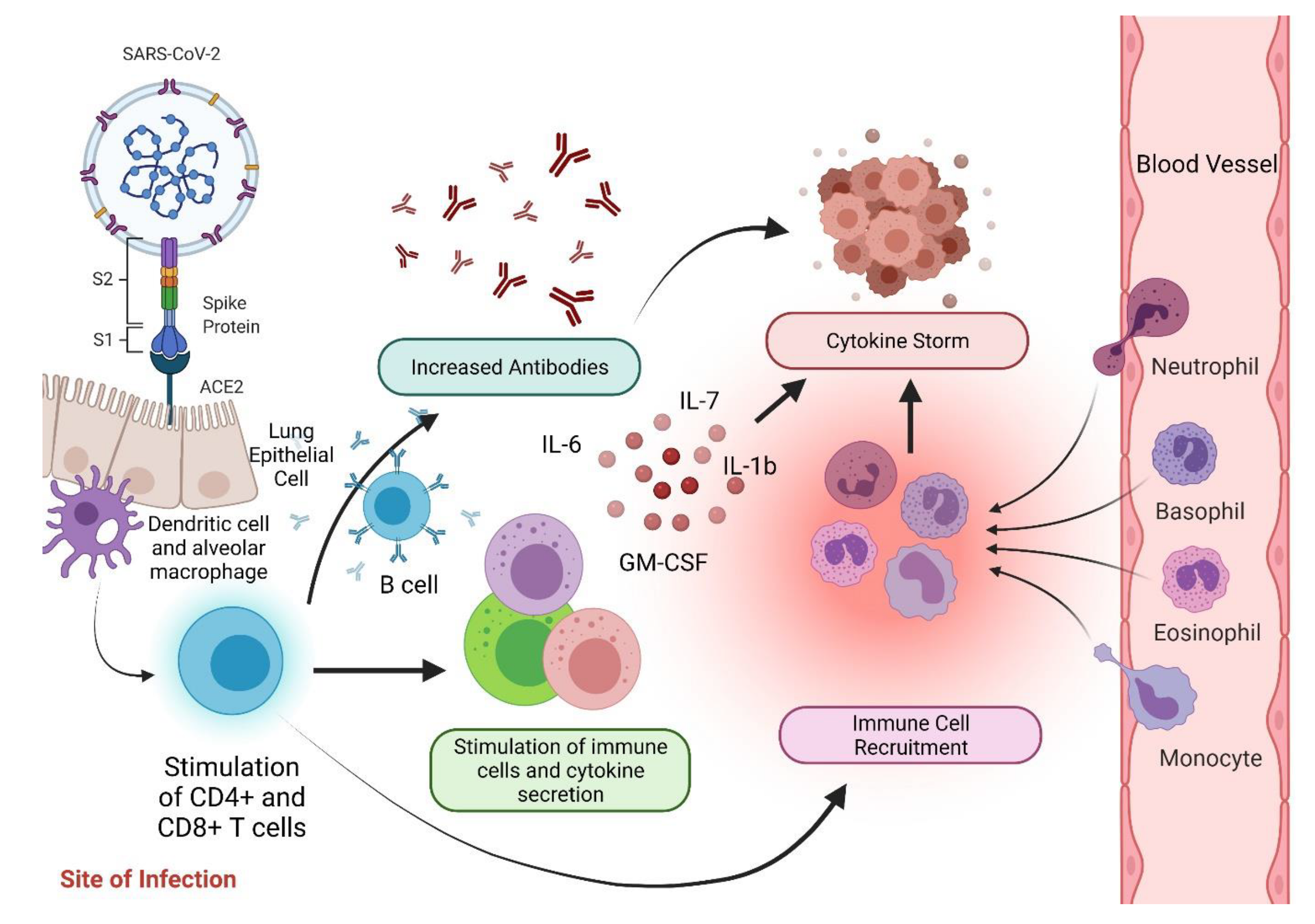
3. Immunopathophysiology of COVID-19
4. Anti-SARS-CoV-2 Vaccines
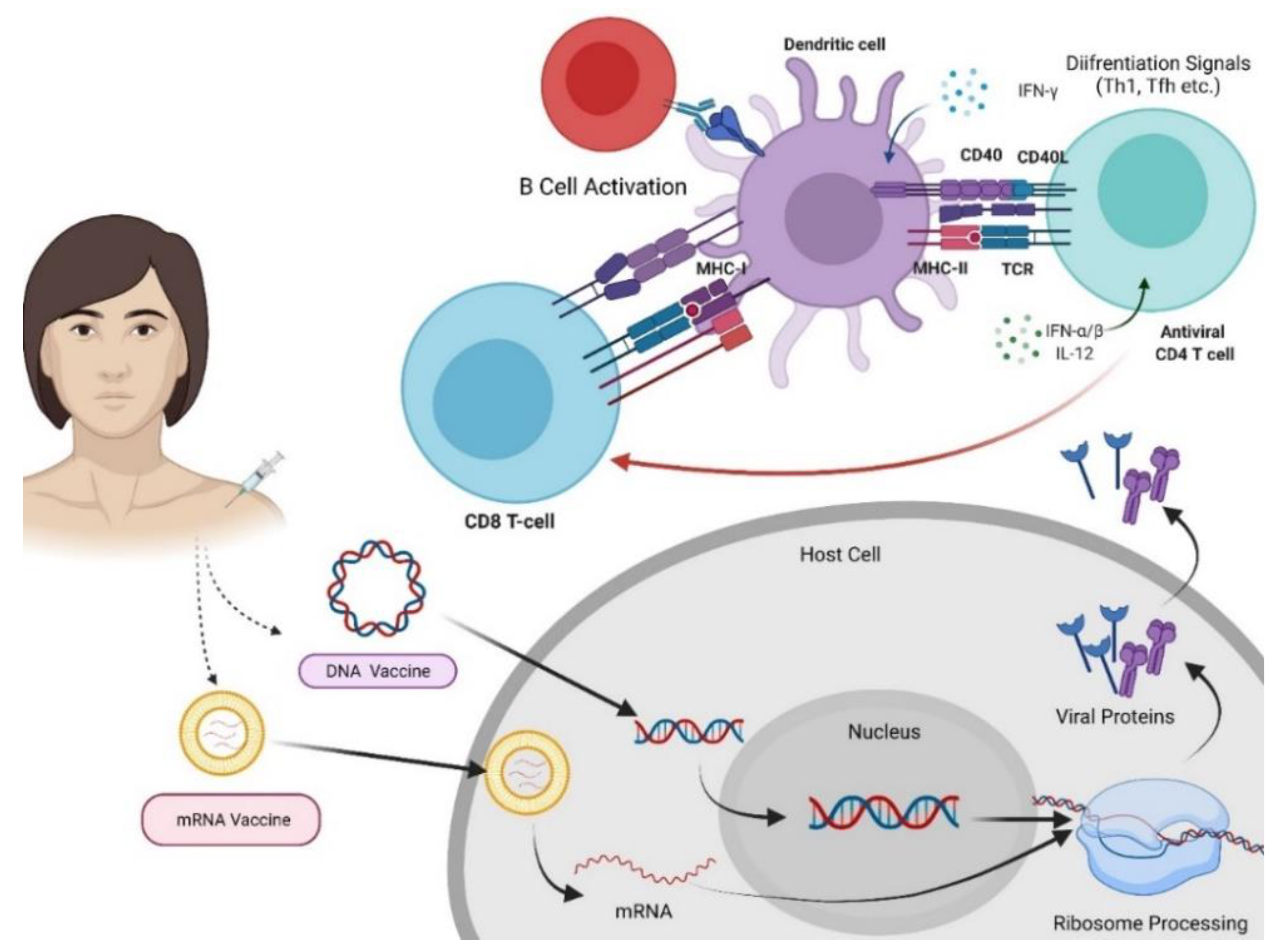
4.1. Nucleic Acid-Based Vaccines
4.1.1. RNA Vaccines
4.1.2. DNA Vaccines
5. Advantages of Nucleic Acid-Based Vaccines
6. Challenges for Nucleic Acid Vaccine Development
- ○
- DNA vaccines can change the genetic composition of the host. DNA vaccines are delivered into the nucleus of the cell and transcribed into mRNA, which enters the cytoplasm, and the cells make the antigen. As such, DNA vaccines are associated with the risk of altering the genetic makeup of the host cell permanently (insertional mutagenesis). mRNA-based vaccines do not pose this risk as they do not enter the nucleus [123,124].
- ○
- Naked DNA has low immunogenicity, and it is essential to include vectors, adjuvants and appropriate delivery methods to increase its immunogenicity.
- ○
- DNA vaccines are relatively cheap to produce compared to protein-based vaccines and are stable, making them viable for storage and worldwide distribution. The challenge for DNA-based vaccines is their poor immunogenicity, often requiring multiple booster injections.
- ○
- mRNA needs to cross the cell membrane to enter the cytoplasm. This is challenging due to its extremely large size, the negative charge of the molecule and degradability. Manufacturing clinical-grade mRNA is also a challenging task.
- ○
- With the emergence of the recent COVID-19 pandemic, a significant number of DNA (26 candidates) and mRNA (35 candidates) vaccines (Table 1) are in preclinical and clinical trials despite these limitations, and eventually, two vaccines have been approved from the mRNA platform for mass application [125,126], and one from DNA platform, i.e., ZycovD (Zydus Cadila, India), has been approved [104].
- ○
- The Pfizer-BioNTech and Moderna vaccines have demonstrated very good efficacy and safety in human trials, despite the evidence of increased risk of blood clots in a small number of subjects. However, the long-term safety, vaccine stability and efficacy still need to be established for this platform and is a subject matter of future studies.
- ○
- The efficacy of nucleic acid-based vaccines is hindered by viral mutations, and the approved mRNA vaccines have demonstrated variable reduced efficacy against these mutant strains as compared with the efficacy against original non-mutated strains [125,126]. As viruses are known to mutate, mutations will continue with the SARS-CoV-2 virus, and hence, constant modifications of the vaccine are required to be effective against the new variants.
- ○
- Some studies have shown that mutations in the target proteins of the SARS-CoV-2 virus may lead to the development of drug and vaccine resistance and eventually lead to vaccine in-efficacy.
- ○
- The mRNA-based vaccines have the advantage of being stable, cost-effective, easy to make, and there are no requirements of purification steps that are commonly used for protein-based vaccines. However, it requires ultra-cold storage limiting its worldwide distribution, and a few booster shots may be required to generate appropriate immunity [62,127,128].
- ○
- The elevated immune response induced by mRNA in the cytoplasm might cause cells to secrete greater portions of type-I IFN and other interferons, which can inhibit mRNA translation and inevitably lead to translational stagnation, RNA degradation, reduced activation of CD8 (cluster of differentiation 8) + T-cells and ultimately immune response cessation [65,78,85].
7. The Future of Nucleic Acid Vaccines
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dai, L.; Gao, G.F. Viral targets for vaccines against COVID-19. Nat. Rev. Immunol. 2020, 21, 73–82. [Google Scholar] [CrossRef]
- Elwood, J.M. Smallpox and its eradication. J. Epidemiol. Community Health 1989, 43, 92. [Google Scholar] [CrossRef] [Green Version]
- Nimmanapalli, R.; Gupta, V. Vaccines the tugboat for prevention-based animal production. Adv. Vet. Poult. Fish. 2019, 469–504. [Google Scholar] [CrossRef]
- Plotkin, S. History of vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 12283–12287. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.P.; Gupta, V. COVID-19 Vaccine: A comprehensive status report. Virus Res. 2020, 288, 198114. [Google Scholar] [CrossRef]
- World Health Organization. WHO SAGE Roadmap for Prioritizing Uses of COVID-19 Vaccines in the Context Of Limited Supply. Available online: https://www.who.int/publications/m/item/who-sage-roadmap-for-prioritizing-uses-of-covid-19-vaccines-in-the-context-of-limited-supply (accessed on 18 October 2021).
- Al-Qahtani, A.A. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Emergence, history, basic and clinical aspects. Saudi J. Biol. Sci. 2020, 27, 2531–2538. [Google Scholar] [CrossRef]
- Chavda, V.P.; Apostolopoulos, V. Mucormycosis—An opportunistic infection in the aged immunocompromised individual: A reason for concern in COVID-19. Maturitas 2021. [Google Scholar] [CrossRef] [PubMed]
- Sohrabi, C.; Alsafi, Z.; O’Neill, N.; Khan, M.; Kerwan, A.; Al-Jabir, A.; Iosifidis, C.; Agha, R. World Health Organization declares global emergency: A review of the 2019 novel coronavirus (COVID-19). Int. J. Surg. 2020, 76, 71–76. [Google Scholar] [CrossRef]
- National Geographic. Moderna’s mRNA Vaccine Reaches Its Final Phase. Here’s How It Works. Available online: https://www.nationalgeographic.com/science/2020/05/moderna-coronavirus-vaccine-how-it-works-cvd/ (accessed on 18 October 2021).
- World Heath Organization. The COVID-19 Vaccine Tracker and Landscape Compiles Detailed Information of Each COVID-19 Vaccine Candidate in Development by Closely Monitoring Their Progress through the Pipeline. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 18 October 2021).
- Zimmer, C.; Corum, J.; Wee, S. Coronavirus Vaccine Tracker. 2021. Available online: https://www.nytimes.com/interactive/2020/science/coronavirus-vaccine-tracker.html (accessed on 18 October 2021).
- McGill COVID19 Vaccine Tracker Team. COVID-19 Vaccine Tracker. 2021. Available online: https://covid19.trackvaccines.org/vaccines/ (accessed on 11 May 2021).
- World Health Organization. The Latest on the COVID-19 Global Situation & Vaccine Development. Available online: https://www.who.int/docs/default-source/coronaviruse/risk-comms-updates/update45-vaccines-developement.pdf?sfvrsn=13098bfc_5 (accessed on 18 October 2021).
- Singh, T.U.; Parida, S.; Lingaraju, M.C.; Kesavan, M.; Kumar, D.; Singh, R.K. Drug repurposing approach to fight COVID-19. Pharmacol. Rep. 2020, 72, 1479–1508. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Restifo, N.P.; Ying, H.; Hwang, L.; Leitner, W. The promise of nucleic acid vaccines. Gene Ther. 2000, 7, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Mittal, K.A.; Manjunath, R.K.; Ranjan, S.; Kaushik, S.; Kumar, V. COVID-19 pandemic: Insights into structure, function, and hACE2 receptor recognition by SARS-CoV-2. PLoS Pathog. 2020, 16, e1008762. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.-F.; Xu, W.; Liu, S.-W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Z.; Chen, Z.; Huang, X.; Xu, M.; He, T.; Zhang, Z. The establishment of reference sequence for SARS-CoV-2 and variation analysis. J. Med. Virol. 2020, 92, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Peng, Y.; Huang, B.; Ding, X.; Wang, X.; Niu, P.; Meng, J.; Zhu, Z.; Zhang, Z.; Wang, J.; et al. Genome Composition and Divergence of the Novel Coronavirus (2019-nCoV) Originating in China. Cell Host Microbe 2020, 27, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.F.-W.; Kok, K.-H.; Zhu, Z.; Chu, H.; To, K.K.-W.; Yuan, S.; Yuen, K.-Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satarker, S.; Nampoothiri, M. Structural Proteins in Severe Acute Respiratory Syndrome Coronavirus-2. Arch. Med. Res. 2020, 51, 482–491. [Google Scholar] [CrossRef]
- Schoeman, D.; Fielding, B.C. Coronavirus envelope protein: Current knowledge. Virol. J. 2019, 16, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Siu, Y.L.; Teoh, K.T.; Lo, J.; Chan, C.M.; Kien, F.; Escriou, N.; Tsao, S.W.; Nicholls, J.M.; Altmeyer, R.; Peiris, J.S.M.; et al. The M, E, and N Structural Proteins of the Severe Acute Respiratory Syndrome Coronavirus Are Required for Efficient Assembly, Trafficking, and Release of Virus-Like Particles. J. Virol. 2008, 82, 11318–11330. [Google Scholar] [CrossRef] [Green Version]
- Nieto-Torres, J.L.; DeDiego, M.L.; Álvarez, E.; Jiménez-Guardeño, J.M.; Regla-Nava, J.A.; Llorente, M.; Kremer, L.; Shuo, S.; Enjuanes, L. Subcellular location and topology of severe acute respiratory syndrome coronavirus envelope protein. Virology 2011, 415, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Voss, D.; Kern, A.; Traggiai, E.; Eickmann, M.; Stadler, K.; Lanzavecchia, A.; Becker, S. Characterization of severe acute respiratory syndrome coronavirus membrane protein. FEBS Lett. 2006, 580, 968–973. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Fan, Y.; Lai, Y.; Han, T.; Li, Z.; Zhou, P.; Pan, P.; Wang, W.; Hu, D.; Liu, X.; et al. Coronavirus infections and immune responses. J. Med. Virol. 2020, 92, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Baruah, V.; Bose, S. Immunoinformatics-aided identification of T cell and B cell epitopes in the surface glycoprotein of 2019-nCoV. J. Med. Virol. 2020, 92, 495–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Liu, S.; Liu, J.; Zhang, Z.; Wan, X.; Huang, B.; Chen, Y.; Zhang, Y. COVID-19: Immunopathogenesis and Immunotherapeutics. Signal Transduct. Target. Ther. 2020, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavda, V.; Feehan, J.; Apostolopoulos, V. A Veterinary Vaccine for SARS-CoV-2: The First COVID-19 Vaccine for Animals. Vaccines 2021, 9, 631. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Vora, L.K.; Vihol, D.R. COVAX-19® Vaccine: Completely blocks virus transmission to non-immune individuals. Clin. Complement. Med. Pharmacol. 2021, 1, 100004. [Google Scholar] [CrossRef]
- Moon, C. Fighting COVID-19 exhausts T cells. Nat. Rev. Immunol. 2020, 20, 277. [Google Scholar] [CrossRef]
- Tan, L.; Wang, Q.; Zhang, D.; Ding, J.; Huang, Q.; Tang, Y.-Q.; Wang, Q.; Miao, H. Lymphopenia predicts disease severity of COVID-19: A descriptive and predictive study. Signal Transduct. Target. Ther. 2020, 5, 1–3. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, W.; Li, J.; Chen, L.; Wang, Y.; Zhang, L.; Yu, L. Clinical features and progression of acute respiratory distress syndrome in coronavirus disease 2019. MedRxiv 2020. [Google Scholar] [CrossRef]
- Smith, C.K.; Trinchieri, G. The interplay between neutrophils and microbiota in cancer. J. Leukoc. Biol. 2018, 104, 701–715. [Google Scholar] [CrossRef]
- Deshmukh, H.S.; Liu, Y.; Menkiti, O.R.; Mei, J.; Dai, N.; O’Leary, C.E.; Oliver, P.M.; Kolls, J.K.; Weiser, J.N.; Worthen, G.S. The microbiota regulates neutrophil homeostasis and host resistance to Escherichia coli K1 sepsis in neonatal mice. Nat. Med. 2014, 20, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Fu, B.; Zheng, X.; Wang, D.; Zhao, C.; Qi, Y.; Sun, R.; Tian, Z.; Xu, X.; Wei, H. Aberrant pathogenic GM-CSF+ T cells and inflammatory CD14+ CD16+ monocytes in severe pulmonary syndrome patients of a new coronavirus. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Wen, W.; Su, W.; Tang, H.; Le, W.; Zhang, X.; Zheng, Y.; Liu, X.; Xie, L.; Li, J.; Ye, J.; et al. Immune cell profiling of COVID-19 patients in the recovery stage by single-cell sequencing. Cell Discov. 2020, 6, 1–18. [Google Scholar] [CrossRef]
- Wu, D.; Yang, X.O. TH17 responses in cytokine storm of COVID-19: An emerging target of JAK2 inhibitor Fedratinib. J. Microbiol. Immunol. Infect. 2020, 53, 368–370. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients with Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Yip, M.S.; Leung, H.L.; Li, P.H.; Cheung, C.Y.; Dutry, I.; Li, D.; Daëron, M.; Bruzzone, R.; Peiris, J.S.M.; Jaume, M. Antibody-dependent enhancement of SARS coronavirus infection and its role in the pathogenesis of SARS. Hong Kong Med. J. 2016, 22, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [Green Version]
- Roncati, L.; Nasillo, V.; Lusenti, B.; Riva, G. Signals of Th2 immune response from COVID-19 patients requiring intensive care. Ann. Hematol. 2020, 99, 1419–1420. [Google Scholar] [CrossRef]
- Malik, J.A.; Mulla, A.H.; Farooqi, T.; Pottoo, F.H.; Anwar, S.; Rengasamy, K.R. Targets and strategies for vaccine development against SARS-CoV-2. Biomed. Pharmacother. 2021, 137, 111254. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2020, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Vora, L.K.; Pandya, A.K.; Patravale, V.B. Intranasal vaccines for SARS-CoV-2: From challenges to potential in COVID-19 management. Drug Discov. Today 2021. [Google Scholar] [CrossRef] [PubMed]
- Van Riel, D.; De Wit, E. Next-generation vaccine platforms for COVID-19. Nat. Mater. 2020, 19, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Wu, S. Progress and Concept for COVID-19 Vaccine Development. Biotechnol. J. 2020, 15, e2000147. [Google Scholar] [CrossRef] [PubMed]
- Mehta, M.; Shyh, G.I. A Review of Remdesivir for COVID-19: Data to Date. Cardiol. Rev. 2020, 28, 332–334. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Singh, R.; Misra, A. Remdesivir in COVID-19: A critical review of pharmacology, pre-clinical and clinical studies. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Frediansyah, A.; Nainu, F.; Dhama, K.; Mudatsir, M.; Harapan, H. Remdesivir and its antiviral activity against COVID-19: A systematic review. Clin. Epidemiol. Glob. Health 2020, 9, 123–127. [Google Scholar] [CrossRef]
- Vasou, A.; Sultanoglu, N.; Goodbourn, S.; Randall, R.; Kostrikis, L.G. Targeting Pattern Recognition Receptors (PRR) for Vaccine Adjuvantation: From Synthetic PRR Agonists to the Potential of Defective Interfering Particles of Viruses. Viruses 2017, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- de Queiroz, N.M.G.; Marinho, F.V.; Chagas, M.A.; Leite, L.C.; Homan, E.J.; de Magalhães, M.T.; Oliveira, S.C. Vaccines for COVID-19: Perspectives from nucleic acid vaccines to BCG as delivery vector system. Microbes Infect. 2020, 22, 515–524. [Google Scholar] [CrossRef]
- Speiser, D.E.; Bachmann, M.F. COVID-19: Mechanisms of Vaccination and Immunity. Vaccines 2020, 8, 404. [Google Scholar] [CrossRef]
- Redding, L.; Weiner, D.B. DNA vaccines in veterinary use. Expert Rev. Vaccines 2009, 8, 1251–1276. [Google Scholar] [CrossRef] [Green Version]
- Piyush, R.; Rajarshi, K.; Chatterjee, A.; Khan, R.; Ray, S. Nucleic acid-based therapy for coronavirus disease 2019. Heliyon 2020, 6, e05007. [Google Scholar] [CrossRef]
- Vogel, F.R.; Sarver, N. Nucleic acid vaccines. Clin. Microbiol. Rev. 1995, 8, 406–410. [Google Scholar] [CrossRef]
- Liu, M.A. A Comparison of Plasmid DNA and mRNA as Vaccine Technologies. Vaccines 2019, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, S.; Jasny, E.; Schmidt, K.E.; Petsch, B. New Vaccine Technologies to Combat Outbreak Situations. Front. Immunol. 2018, 9, 1963. [Google Scholar] [CrossRef] [Green Version]
- Risitano, A.M.; Mastellos, D.C.; Huber-Lang, M.; Yancopoulou, D.; Garlanda, C.; Ciceri, F.; Lambris, J.D. Complement as a target in COVID-19? Nat. Rev. Immunol. 2020, 20, 343–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Yang, K.; Li, R.; Zhang, L. mRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection. Int. J. Mol. Sci. 2020, 21, 6582. [Google Scholar] [CrossRef] [PubMed]
- Teijaro, J.R.; Farber, D.L. COVID-19 vaccines: Modes of immune activation and future challenges. Nat. Rev. Immunol. 2021, 21, 195–197. [Google Scholar] [CrossRef]
- Heine, A.; Juranek, S.; Brossart, P. Clinical and immunological effects of mRNA vaccines in malignant diseases. Mol. Cancer 2021, 20, 1–20. [Google Scholar] [CrossRef]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. npj Vaccines 2021, 6, 1–17. [Google Scholar] [CrossRef]
- McNamara, M.A.; Nair, S.K.; Holl, E.K. RNA-Based Vaccines in Cancer Immunotherapy. J. Immunol. Res. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettini, E.; Locci, M. SARS-CoV-2 mRNA Vaccines: Immunological Mechanism and Beyond. Vaccines 2021, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Widge, A.T.; Rouphael, N.G.; Jackson, L.A.; Anderson, E.J.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Durability of Responses after SARS-CoV-2 mRNA-1273 Vaccination. N. Engl. J. Med. 2021, 384, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Paik, D.H.; Farber, D.L. Anti-viral protective capacity of tissue resident memory T cells. Curr. Opin. Virol. 2020, 46, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Lange, A.; Borowik, A.; Bocheńska, J.; Rossowska, J.; Jaskuła, E. Immune Response to COVID-19 mRNA Vaccine—A Pilot Study. Vaccines 2021, 9, 488. [Google Scholar] [CrossRef]
- Dey, A.; Rajanathan, T.C.; Chandra, H.; Pericherla, H.P.; Kumar, S.; Choonia, H.S.; Bajpai, M.; Singh, A.K.; Sinha, A.; Saini, G.; et al. Immunogenic potential of DNA vaccine candidate, ZyCoV-D against SARS-CoV-2 in animal models. Vaccine 2021, 39, 4108–4116. [Google Scholar] [CrossRef]
- Jackson, N.A.C.; Kester, K.E.; Casimiro, D.; Gurunathan, S.; DeRosa, F. The promise of mRNA vaccines: A biotech and industrial perspective. npj Vaccines 2020, 5, 1–6. [Google Scholar] [CrossRef]
- Maruggi, G.; Zhang, C.; Li, J.; Ulmer, J.B.; Yu, D. mRNA as a Transformative Technology for Vaccine Development to Control Infectious Diseases. Mol. Ther. 2019, 27, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Hogan, M.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Zeng, J.; Yan, J. COVID-19 mRNA vaccines. J. Genet. Genom. 2021, 48, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.; Rosenecker, J. Nanotechnologies in delivery of mRNA therapeutics using nonviral vector-based delivery systems. Gene Ther. 2017, 24, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Sun, X.; Aikins, M.E.; Moon, J.J. Non-viral COVID-19 vaccine delivery systems. Adv. Drug Deliv. Rev. 2020, 169, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. COVID-19 vaccines poised for launch, but impact on pandemic unclear. Nat. Biotechnol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rahman, G. RNA Covid-19 Vaccines Will Not Change Your DNA. 2021. Available online: https://fullfact.org/online/rna-vaccine-covid/ (accessed on 2 June 2021).
- Kauffman, K.J.; Webber, M.; Anderson, D.G. Materials for non-viral intracellular delivery of messenger RNA therapeutics. J. Control. Release 2016, 240, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of Pseudouridine Into mRNA Yields Superior Nonimmunogenic Vector with Increased Translational Capacity and Biological Stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Muramatsu, H.; Ludwig, J.; Weissman, D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 2011, 39, e142. [Google Scholar] [CrossRef] [Green Version]
- Thess, A.; Grund, S.; Mui, B.L.; Hope, M.J.; Baumhof, P.; Fotin-Mleczek, M.; Schlake, T. Sequence-engineered mRNA without Chemical Nucleoside Modifications Enables an Effective Protein Therapy in Large Animals. Mol. Ther. 2015, 23, 1456–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Tsui, N.B.; Ng, E.K.; Lo, Y.D. Stability of Endogenous and Added RNA in Blood Specimens, Serum, and Plasma. Clin. Chem. 2002, 48, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Kallen, K.-J.; Theß, A. A development that may evolve into a revolution in medicine: mRNA as the basis for novel, nucleotide-based vaccines and drugs. Ther. Adv. Vaccines 2013, 2, 10–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging Technologies for Low-Cost, Rapid Vaccine Manufacture. Biotechnol. J. 2018, 14, e1800376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383. [Google Scholar] [CrossRef]
- Liu, L.; Wei, Q.; Lin, Q.; Fang, J.; Wang, H.; Kwok, H.; Tang, H.; Nishiura, K.; Peng, J.; Tan, Z.; et al. Anti–spike IgG causes severe acute lung injury by skewing macrophage responses during acute SARS-CoV infection. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. N. Engl. J. Med. 2020, 383, 2427–2438. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Moderna COVID-19 Vaccine. Available online: https://www.fda.gov/emergency-preparedness-and-response/coronavirus-disease-2019-covid-19/moderna-covid-19-vaccine (accessed on 27 September 2021).
- Vogel, A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.; Kranz, L.M.; Walzer, K.C.; Hein, S.; Güler, A.; et al. A prefusion SARS-CoV-2 spike RNA vaccine is highly immunogenic and prevents lung infection in non-human primates. BioRxiv 2020. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Vogler, I.; Derhovanessian, E.; Kranz, L.M.; Vormehr, M.; Quandt, J.; Bidmon, N.; Ulges, A.; Baum, A.; et al. BNT162b2 induces SARS-CoV-2-neutralising antibodies and T cells in humans. medRxiv 2020. [Google Scholar] [CrossRef]
- USFDA. FDA Approves First COVID-19 Vaccine. 2021. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-covid-19-vaccine (accessed on 27 September 2021).
- Reautschnig, P.; Vogel, P.; Stafforst, T. The notorious R.N.A. in the spotlight—Drug or target for the treatment of disease. RNA Biol. 2016, 14, 651–668. [Google Scholar] [CrossRef] [Green Version]
- Stitz, L.; Vogel, A.; Schnee, M.; Voss, D.; Rauch, S.; Mutzke, T.; Ketterer, T.; Kramps, T.; Petsch, B. A thermostable messenger RNA based vaccine against rabies. PLOS Negl. Trop. Dis. 2017, 11, e0006108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostolopoulos, V.; Plebanski, M. The evolution of DNA vaccines. Curr. Opin. Mol. Ther. 2000, 2, 441–447. [Google Scholar] [PubMed]
- Chavda, V.P.; Pandya, R.; Apostolopoulos, V. DNA vaccines for SARS-CoV-2: Towards third generation vaccination era. Expert Rev. Vaccines 2021. [Google Scholar] [CrossRef]
- Singh, S. India Gives Emergency Approval for World’s First COVID-19 DNA Vaccine. Available online: https://www.reuters.com/business/healthcare-pharmaceuticals/india-approves-zydus-cadilas-covid-19-vaccine-emergency-use-2021-08-20/ (accessed on 21 August 2021).
- Cadila Healthcare Ltd. Zydus Cadila Receives Approvals from the DCGI to Start Phase III Clinical Trial of ZyCoV-D—Fully Indigenously Developed Vaccine. 2021. Available online: https://zyduscadila.com/public/pdf/pressrelease/Zydus_Cadila_receives_approvals_from_the_DCGI_to_start_Phase_III_Clinical_Trial_of_ZyCoV_D_fully_indigenously_developed_vaccine_-3_1_2021.pdf (accessed on 1 June 2021).
- Momin, T.; Kansagra, K.; Patel, H.; Sharma, S.; Sharma, B.; Patel, J.; Mittal, R.; Sanmukhani, J.; Maithal, K.; Dey, A.; et al. Safety and Immunogenicity of a DNA SARS-CoV-2 vaccine (ZyCoV-D): Results of an open-label, non-randomized phase I part of phase I/II clinical study by intradermal route in healthy subjects in India. EClinicalMedicine 2021, 38, 101020. [Google Scholar] [CrossRef]
- Modjarrad, K.; Roberts, C.C.; Mills, K.T.; Castellano, A.R.; Paolino, K.; Muthumani, K.; Reuschel, E.L.; Robb, M.L.; Racine, T.; Oh, M.-D.; et al. Safety and immunogenicity of an anti-Middle East respiratory syndrome coronavirus DNA vaccine: A phase 1, open-label, single-arm, dose-escalation trial. Lancet Infect. Dis. 2019, 19, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.R.F.; Patel, A.; Ramos, S.; Elwood, D.; Zhu, X.; Yan, J.; Gary, E.N.; Walker, S.N.; Schultheis, K.; Purwar, M.; et al. Immunogenicity of a DNA vaccine candidate for COVID-19. Nat. Commun. 2020, 11, 2601. [Google Scholar] [CrossRef] [PubMed]
- Tebas, P.; Yang, S.; Boyer, J.D.; Reuschel, E.L.; Patel, A.; Christensen-Quick, A.; Andrade, V.M.; Morrow, M.P.; Kraynyak, K.; Agnes, J.; et al. Safety and immunogenicity of INO-4800 DNA vaccine against SARS-CoV-2: A preliminary report of an open-label, Phase 1 clinical trial. EClinicalMedicine 2020, 31, 100689. [Google Scholar] [CrossRef]
- Schmeer, M.; Buchholz, T.; Schleef, M. Plasmid DNA Manufacturing for Indirect and Direct Clinical Applications. Hum. Gene Ther. 2017, 28, 856–861. [Google Scholar] [CrossRef]
- Brocato, R.L.; Kwilas, S.A.; Josleyn, M.D.; Long, S.; Zeng, X.; Perley, C.C.; Principe, L.M.; Somerville, B.; Cohen, M.V.; Hooper, J.W. Small animal jet injection technique results in enhanced immunogenicity of hantavirus DNA vaccines. Vaccine 2021, 39, 1101–1110. [Google Scholar] [CrossRef]
- Sokołowska, E.; Błachnio-Zabielska, A.U. A Critical Review of Electroporation as A Plasmid Delivery System in Mouse Skeletal Muscle. Int. J. Mol. Sci. 2019, 20, 2776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.A. Immunologic Basis of Vaccine Vectors. Immunity 2010, 33, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlake, T.; Thess, A.; Thran, M.; Jordan, I. mRNA as novel technology for passive immunotherapy. Experientia 2018, 76, 301–328. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Weissman, D. Nucleoside Modified mRNA Vaccines for Infectious Diseases. Methods Mol. Biol. 2016, 1499, 109–121. [Google Scholar] [CrossRef]
- Lim, B.; Lee, K. Stability of the Osmoregulated Promoter-DerivedproPmRNA Is Posttranscriptionally Regulated by RNase III in Escherichia coli. J. Bacteriol. 2015, 197, 1297–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarghampoor, F.; Azarpira, N.; Khatami, S.R.; Behzad-Behbahani, A.; Foroughmand, A.M. Improved translation efficiency of therapeutic mRNA. Gene 2019, 707, 231–238. [Google Scholar] [CrossRef]
- Li, S.S.; Kochar, N.K.; Elizaga, M.; Hay, C.M.; Wilson, G.J.; Cohen, K.W.; De Rosa, S.C.; Xu, R.; Ota-Setlik, A.; Morris, D.; et al. DNA Priming Increases Frequency of T-Cell Responses to a Vesicular Stomatitis Virus HIV Vaccine with Specific Enhancement of CD8 + T-Cell Responses by Interleukin-12 Plasmid DNA. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef] [Green Version]
- Apostolopoulos, V.; Weiner, D.B. Development of more efficient and effective DNA vaccines. Expert Rev. Vaccines 2009, 8, 1133–1134. [Google Scholar] [CrossRef]
- Silveira, M.M.; Moreira, G.M.S.G.; Mendonça, M. DNA vaccines against COVID-19: Perspectives and challenges. Life Sci. 2020, 267, 118919. [Google Scholar] [CrossRef]
- Tang, C.-K.; Sheng, K.-C.; Apostolopoulos, V.; Pietersz, G.A. Protein/peptide and DNA vaccine delivery by targeting C-type lectin receptors. Expert Rev. Vaccines 2008, 7, 1005–1018. [Google Scholar] [CrossRef]
- Chandler, M.; Johnson, M.B.; Panigaj, M.; Afonin, K.A. Innate immune responses triggered by nucleic acids inspire the design of immunomodulatory nucleic acid nanoparticles (NANPs). Curr. Opin. Biotechnol. 2019, 63, 8–15. [Google Scholar] [CrossRef]
- Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and Challenges in the Delivery of mRNA-Based Vaccines. Pharmaceutics 2020, 12, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, S.S.; Prazeres, D.M.; Azevedo, A.M.; Marques, M.P. mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine 2021, 39, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Anthony, D.P.S.; Sivakumar, K.; Venugopal, P.; Sriram, D.K.; George, M. Can mRNA Vaccines Turn the Tables During the COVID-19 Pandemic? Current Status and Challenges. Clin. Drug Investig. 2021, 41, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Hossain, K.; Hassanzadeganroudsari, M.; Feehan, J.; Apostolopoulos, V. COVID-19 Vaccines in the Pipeline, Are Antibodies Adequate? Vaccines 2021, 9, 241. [Google Scholar] [CrossRef] [PubMed]
- Hossain, K.; Hassanzadeganroudsari, M.; Apostolopoulos, V. The emergence of new strains of SARS-CoV-2. What does it mean for COVID-19 vaccines? Expert Rev. Vaccines 2021, 20, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.S.; O’Halloran, J.A.; Kalaidina, E.; Kim, W.; Schmitz, A.J.; Zhou, J.Q.; Lei, T.; Thapa, M.; Chen, R.E.; Case, J.B.; et al. SARS-CoV-2 mRNA vaccines induce persistent human germinal centre responses. Nature 2021, 596, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Hall, V.G.; Ferreira, V.H.; Ku, T.; Ierullo, M.; Majchrzak-Kita, B.; Chaparro, C.; Selzner, N.; Schiff, J.; McDonald, M.; Tomlinson, G.; et al. Randomized Trial of a Third Dose of mRNA-1273 Vaccine in Transplant Recipients. N. Engl. J. Med. 2021, 385, 1244–1246. [Google Scholar] [CrossRef] [PubMed]
- Forni, G.; Mantovani, A. COVID-19 vaccines: Where we stand and challenges ahead. Cell Death Differ. 2021, 28, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Calina, D.; Docea, A.O.; Petrakis, D.; Egorov, A.M.; Ishmukhametov, A.A.; Gabibov, A.G.; Shtilman, M.I.; Kostoff, R.; Carvalho, F.; Vinceti, M.; et al. Towards effective COVID-19 vaccines: Updates, perspectives and challenges (Review). Int. J. Mol. Med. 2020, 46, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Freyn, A.W.; da Silva, J.R.; Rosado, V.C.; Bliss, C.M.; Pine, M.; Mui, B.L.; Tam, Y.K.; Madden, T.D.; Ferreira, L.C.D.S.; Weissman, D.; et al. A Multi-Targeting, Nucleoside-Modified mRNA Influenza Virus Vaccine Provides Broad Protection in Mice. Mol. Ther. 2020, 28, 1569–1584. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.A.; Fuhr, R.; Smolenov, I.; Ribeiro, A.; Panther, L.; Watson, M.; Senn, J.J.; Smith, M.; Almarsson, Ö.; Pujar, H.S.; et al. mRNA vaccines against H10N8 and H7N9 influenza viruses of pandemic potential are immunogenic and well tolerated in healthy adults in phase 1 randomized clinical trials. Vaccine 2019, 37, 3326–3334. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, J.B.; Mansoura, M.K.; Geall, A.J. Vaccines ‘on demand’: Science fiction or a future reality. Expert Opin. Drug Discov. 2015, 10, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, E. Mix-and-match COVID vaccines trigger potent immune response. Nature 2021, 593, 491. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Vaccine Name | Innovator/Country | Vaccine Platform | Vaccine-Triggered Immune Response | Stage of Clinical Development | Clinical Trial ID Number (https://covid19.trackvaccines.org/vaccines/79/)— Accessed on 18 August 2021) |
|---|---|---|---|---|---|
| mRNA-1273 | Moderna/USA | RNA-based | Post-administration of mRNA vaccine, there is a trigger of type-1 interferon production, which subsequently promotes the Th1 response that is the characteristic of actual viral infection [69,70,71]. The innate immune responses from helper T-cells prime both CD8+ and CD4+ T cells to differentiate into effector and memory subsets [72]. | Emergency use approved (EUA) in 72 Countries. This vaccine is also manufactured by Takeda (TAK-919) | PHASE 1: NCT04813796, NCT04785144, NCT04839315, NCT04889209, NCT04283461 |
| PHASE 2: ISRCTN73765130 NCT04847050, NCT04889209, NCT04649151, NCT04748471, NCT04761822, and NCT04405076 NCT04894435 NCT04796896 | |||||
| PHASE 3: NCT04860297, NCT04649151, NCT04811664, and NCT04470427 NCT04796896 NCT04805125 NCT04806113 | |||||
| BNT162b2 (Tozinameran, Comirnaty) | Pfizer & BioNTech/USA | RNA-based (Encodes a prefusion stabilized, membrane-anchored SARS-CoV-2 full-length Spike protein) | The mRNA serves as an antigen as well as an adjuvant that will stimulate both adaptive and innate immune responses, respectively. Toll-like receptor 7 (TLR7) and melanoma differentiation-associated 5 (MDA5) are triggered by mRNA, which stimulates S-protein-specific naive T-cells, which become activated and differentiated into effector cells to form cytotoxic T-lymphocytes or helper T-cells. Strong Th1 cell response helps in antibody-secreting plasma cells. Stimulation of the type-1 interferon also aids in T-cell memory [66,72,73,74] | EUA in 99 countries. | PHASE 1: EUCTR2020-001038-36, and NCT04380701 NCT04839315, NCT04889209, NCT04816643 NCT04588480 |
| PHASE 2: ISRCTN73765130 and ISRCTN69254139 EUCTR2020-001038-36, and NCT04380701 NCT04368728 NCT04894435 NCT04889209, NCT04761822 and NCT04754594 NCT04824638 NCT04860739 and EUCTR2021-001978-37 NCT04649021 NCT04588480 | |||||
| PHASE 3: NCT04368728 NCT04805125 NCT04800133 NCT04816669, NCT04713553, and NCT04754594 | |||||
| TAK—919 (Moderna formulation) | Takeda/Japan | RNA-based | Post-administration of mRNA vaccine, there is a trigger of type-1 interferon production, which subsequently promotes the Th1 response that is the characteristic of actual viral infection [69,70,71]. The innate immune responses from helper T-cells prime both CD8+ and CD4+ T cells to differentiate into effector and memory subsets [72]. | EUA in 1 country. | PHASE 1 and PHASE 2: NCT04677660 |
| mRNA | Walvax/China | RNA-based | Post-vaccination, the mRNA binds with TLR7 and MDA5, which triggers IFN1 production along with a strong Th1 cell response that helps in antibody-secreting plasma cells [66,72]. | Under trials in 4 countries. Phase 1: 2 trials | PHASE 1: ChiCTR2000034112, and ChiCTR2000039212 |
| Phase 2: 1 trial | PHASE 2: ChiCTR2100041855 | ||||
| Phase 3: 1 trial | PHASE 3: NCT04847102 | ||||
| CVnCov | Curevac/Germany | RNA-based | Under trials in 12 countries. Phase 1: 1 trials | PHASE 1: NCT04449276 | |
| Phase 2: 4 trials | PHASE 2: ISRCTN73765130 2020-003998-22 NCT04652102 NCT04515147, PER-054-20 | ||||
| Phase 3: 6 trials | PHASE 3: NCT04838847 and NCT04848467 NCT04860258 EUCTR2020-004066-19, and NCT04674189 and NCT04652102 2020-003998-22 | ||||
| BNT162b1 | Pfizer & BioNTech/USA | RNA-based (Nucleoside-modified mRNA vaccine that encodes the trimerized receptor-binding domain) | Under trials in 5 countries. Phase 1: 2 trials | PHASE 1: EUCTR2020-001038-36, and NCT04380701 ChiCTR2000034825, and NCT04523571 | |
| Phase 2: 2 trials | PHASE 2: EUCTR2020-001038-36, and NCT04380701 NCT04368728 | ||||
| Phase 3: 1 trials | PHASE 3: NCT04368728 | ||||
| MRT5500 | Sanofi Pasteur/USA | RNA-based | Under trials in 1 country. Phase 1 and 2: 1 trial | PHASE 1 and PHASE 2: EUCTR2020-001038-36, NCT04380701 | |
| EXG 5003 | Elixirgen Therapeutic Inc./USA | RNA-based | Under trials in 1 country. Phase 1: 1 trial Phase 2: 1 trial | PHASE 1 and PHASE 2: NCT04863131 | |
| BNT162a1 | Pfizer & BioNTech/Germany | RNA-based (Encodes an optimized SARS-CoV-2 receptor-binding domain) | Under phase 1 and phase 2 trials in Germany | PHASE 1 and PHASE 2: EUCTR2020-001038-36, NCT04380701 | |
| BNT162c2 | Pfizer & BioNTech/Germany | RNA-based (A candidate using self-amplifying mRNA | Under phase 1 and phase 2 trials in Germany | PHASE 1 and PHASE 2: EUCTR2020-001038-36, NCT04380701 | |
| BNT162b3 | Pfizer & BioNTech/Germany | RNA-based (A candidate using self-amplifying mRNA | Under phase 1 and phase 2 trials in Germany | PHASE 1 and PHASE 2: NCT04537949, and EUCTR2020-003267-26-DE | |
| DS-5670a | Daiichi Sankyo Co., Ltd./Japan | RNA-based | Under phase 1 and 2 trial in Japan | PHASE 1 and PHASE 2: NCT04821674 | |
| Chulacov19 | Chulalongkorn University/Thailand | RNA-based | Under phase 1 and phase 2 trial in Thailand | PHASE 1 and PHASE 2: NCT04566276 | |
| LUNAR-cov19/ARCT-021 | Arcturus Therapeutics inc/USA | RNA-based | Phase 1: 1 trials | PHASE 1: NCT04480957 | |
| Phase 2: 3 trials | PHASE 2: NCT04668339 NCT04728347 and NCT04480957 | ||||
| PTX-COVID19-B | Providence therapeutics holding Inc/Canada | RNA-based | Under phase 1 trial in Canada | PHASE 1: PRO-CL-001, NCT04765436 | |
| HDT-301 | Senai cimatec/Brazil | RNA-based | Under phase 1 trial | PHASE 1: NCT04844268 | |
| mRNA-1283 | Moderna/USA | RNA-based | Under phase 1 trial in America | PHASE 1: NCT04813796 | |
| mRNACOVID19 vaccine | Stemirna Therapeutics Co., Ltd./China | RNA-based | Phase 1 trial in 0 country. | PHASE 1: ChiCTR2100045984 | |
| LNP-nCoVsaRNA | Imperial/UK | RNA-based | This vaccine is on hold | PHASE 1: ISRCTN17072692 | |
| AG0302-COVID19 | AnGes/Japan | DNA-based | Post-vaccination, dsDNA triggers TLR9 to induce type-I interferon, which stimulates S-protein-specific naive T-cells, which become activated and differentiated into effector cells to form cytotoxic T-lymphocytes or helper T-cells [66,72]. | Under phase 1, 2 and 3 trials in Japan | PHASE 1: NCT04527081 |
| PHASE 2: NCT04655625 and NCT04527081 | |||||
| PHASE 3: NCT04655625 | |||||
| ZyCoV-D | Zydus cadila/India | DNA-based | Post vaccination dsDNA triggers TLR9 to induce type I interferon which stimulates S-protein-specific naive T cells, which become activated and differentiated into effector cells to form cytotoxic T-lymphocytes or helper T cells. Strong Th1 cell response helps in antibody-secreting plasma cells. Stimulation of Type 1 interferon also aids in T cell memory [66,75]. | EUA in India | PHASE 1: CTRI/2020/07/026352 and CTRI/2021/03/032051 |
| PHASE 2: CTRI/2020/07/026352 and CTRI/2021/03/032051 | |||||
| PHASE 3: CTRI/2021/01/030416 | |||||
| INO-4800 | Inovio/USA | DNA-based | Post-vaccination, dsDNA triggers TLR9 to induce type-I interferon, which stimulates S-protein-specific naive T-cells, which become activated and differentiated into effector cells to form cytotoxic T-lymphocytes or helper T-cells [66,72]. | Under phase 1, 2 and 3 trials in 3 countries | PHASE 1: NCT04336410 NCT04447781 |
| PHASE 2: NCT04642638 ChiCTR2000040146 NCT04447781 | |||||
| PHASE 3: NCT04642638 | |||||
| GX-19 | Genexine/Korea | DNA-based | Under phase 1 and phase 2 trials in Korea | PHASE 1 and PHASE 2: NCT04715997 and NCT04445389 | |
| AG0301-COVID19 | AnGes/Japan | DNA-based | Under phase 1 and 2 trials in Japan | PHASE 1 and PHASE 2: NCT04463472 | |
| GLS-5310 | GeneOne Life Science Inc./Korea | DNA-based | Under phase 1 and 2 trials in Korea | PHASE 1 and PHASE 2: NCT04673149 | |
| COVID-eVax | Takis/Italy | DNA-based | Under phase 1 and 2 trial in Italy | PHASE 1 and PHASE 2: EUCTR2020-003734-20, and NCT04788459 | |
| COVIGEN | University of Sydney/Australia | DNA-based | Under phase 1 trial in Australia | PHASE 1: NCT04742842 | |
| bacTRL-Spike | Symvivo/Australia | DNA-based | Under phase 1 trial in Australia | PHASE 1: NCT04334980 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chavda, V.P.; Hossain, M.K.; Beladiya, J.; Apostolopoulos, V. Nucleic Acid Vaccines for COVID-19: A Paradigm Shift in the Vaccine Development Arena. Biologics 2021, 1, 337-356. https://doi.org/10.3390/biologics1030020
Chavda VP, Hossain MK, Beladiya J, Apostolopoulos V. Nucleic Acid Vaccines for COVID-19: A Paradigm Shift in the Vaccine Development Arena. Biologics. 2021; 1(3):337-356. https://doi.org/10.3390/biologics1030020
Chicago/Turabian StyleChavda, Vivek P., Md Kamal Hossain, Jayesh Beladiya, and Vasso Apostolopoulos. 2021. "Nucleic Acid Vaccines for COVID-19: A Paradigm Shift in the Vaccine Development Arena" Biologics 1, no. 3: 337-356. https://doi.org/10.3390/biologics1030020