
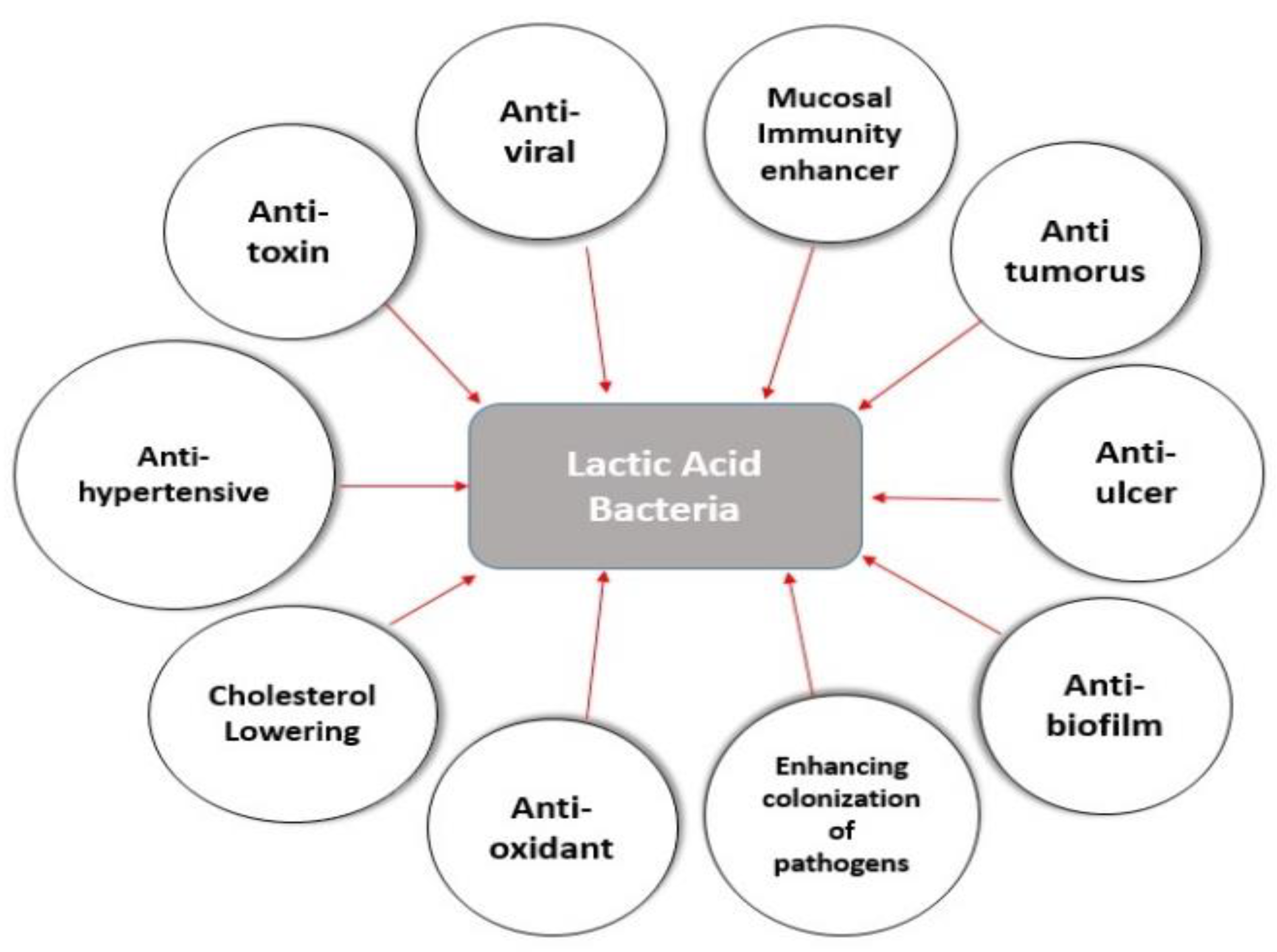
Lactic Acid Bacteria as Mucosal Immunity Enhancers and Antivirals through Oral Delivery

Abstract
:1. Introduction
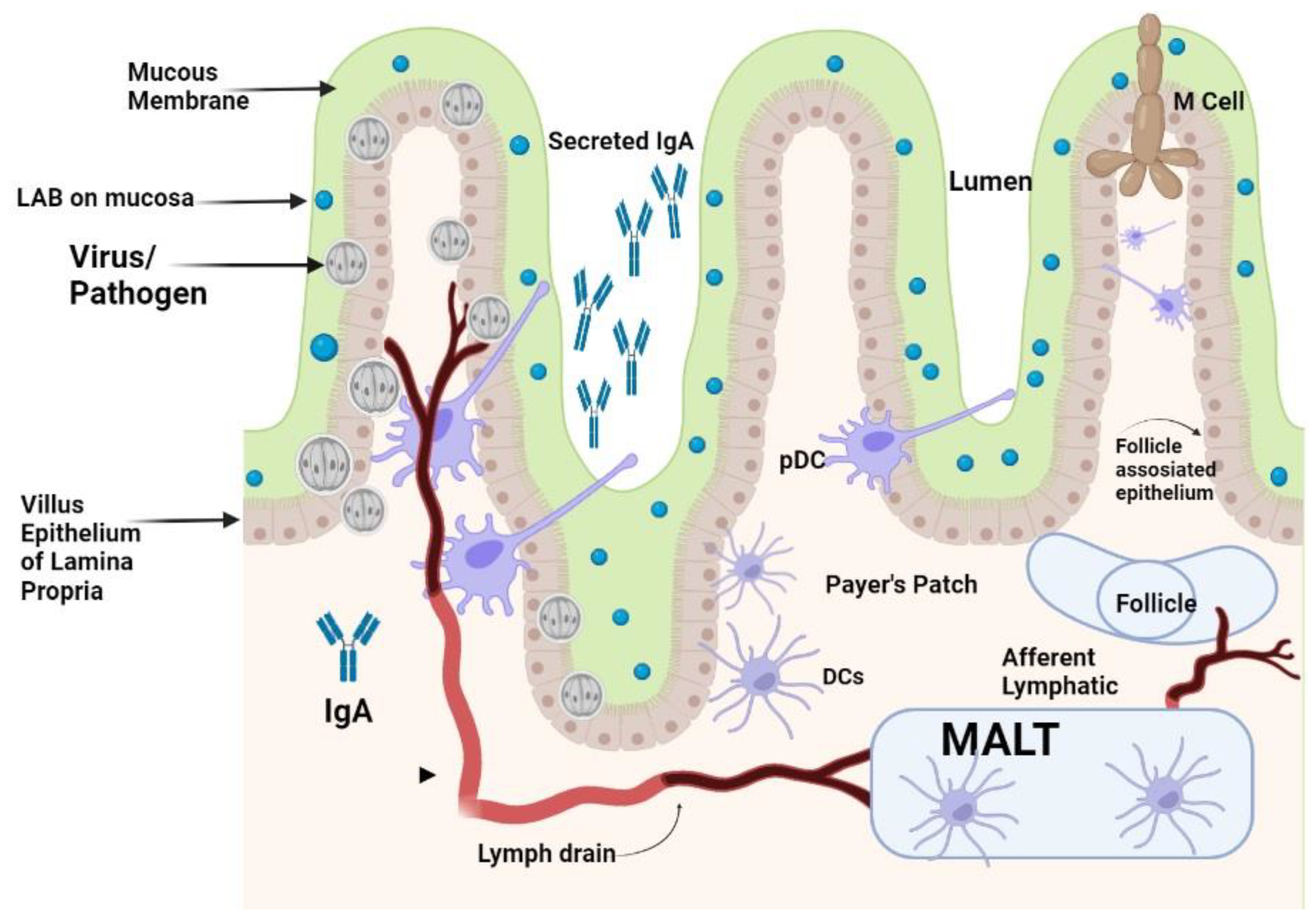
2. LAB as Immunomodulators and Mucosal Immunity Enhancers
2.1. Immunomodulatory Responses of LAB on Cytokines and Immune Cells
2.2. PRRs-Associated Immunomodulation
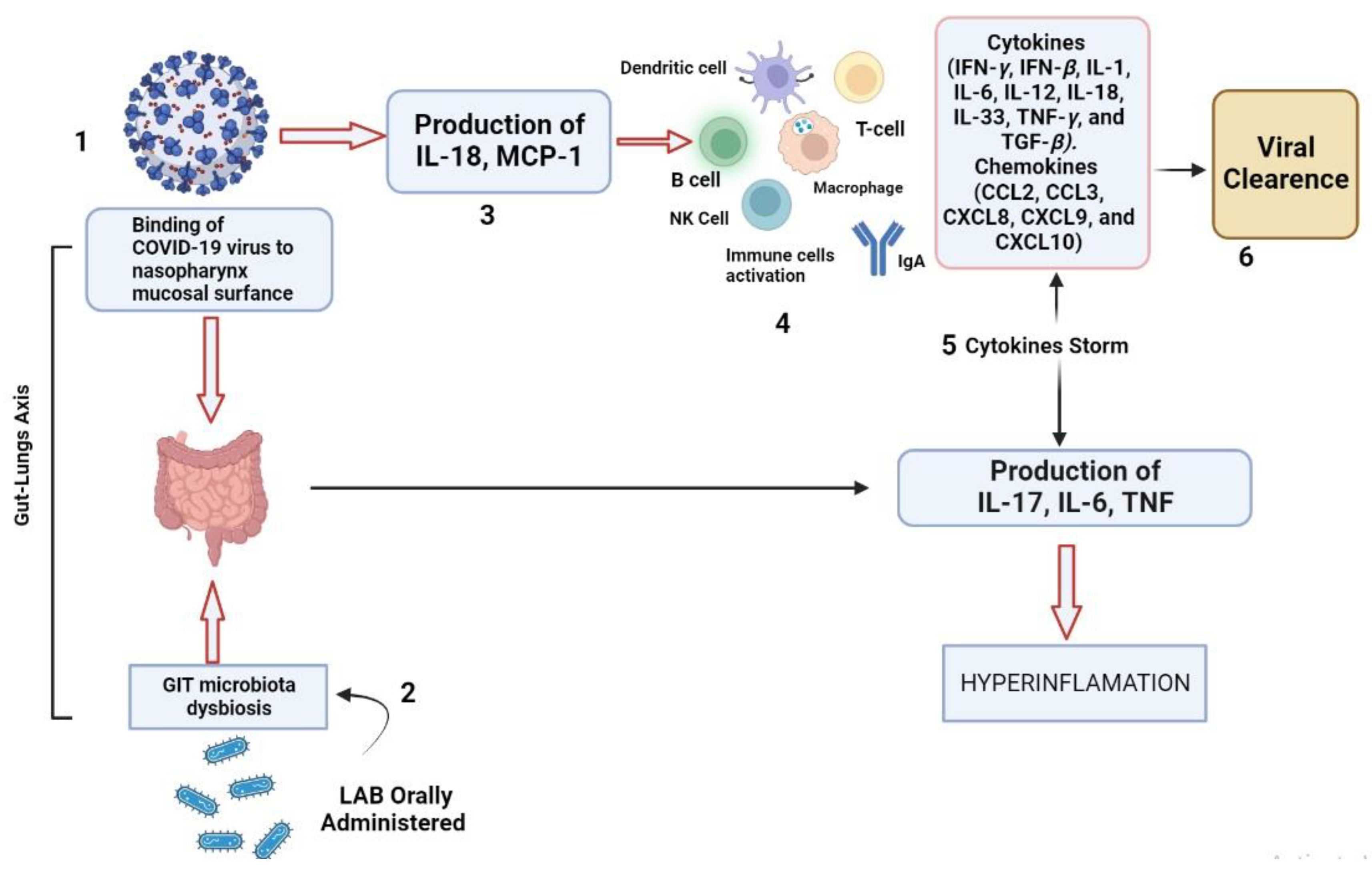
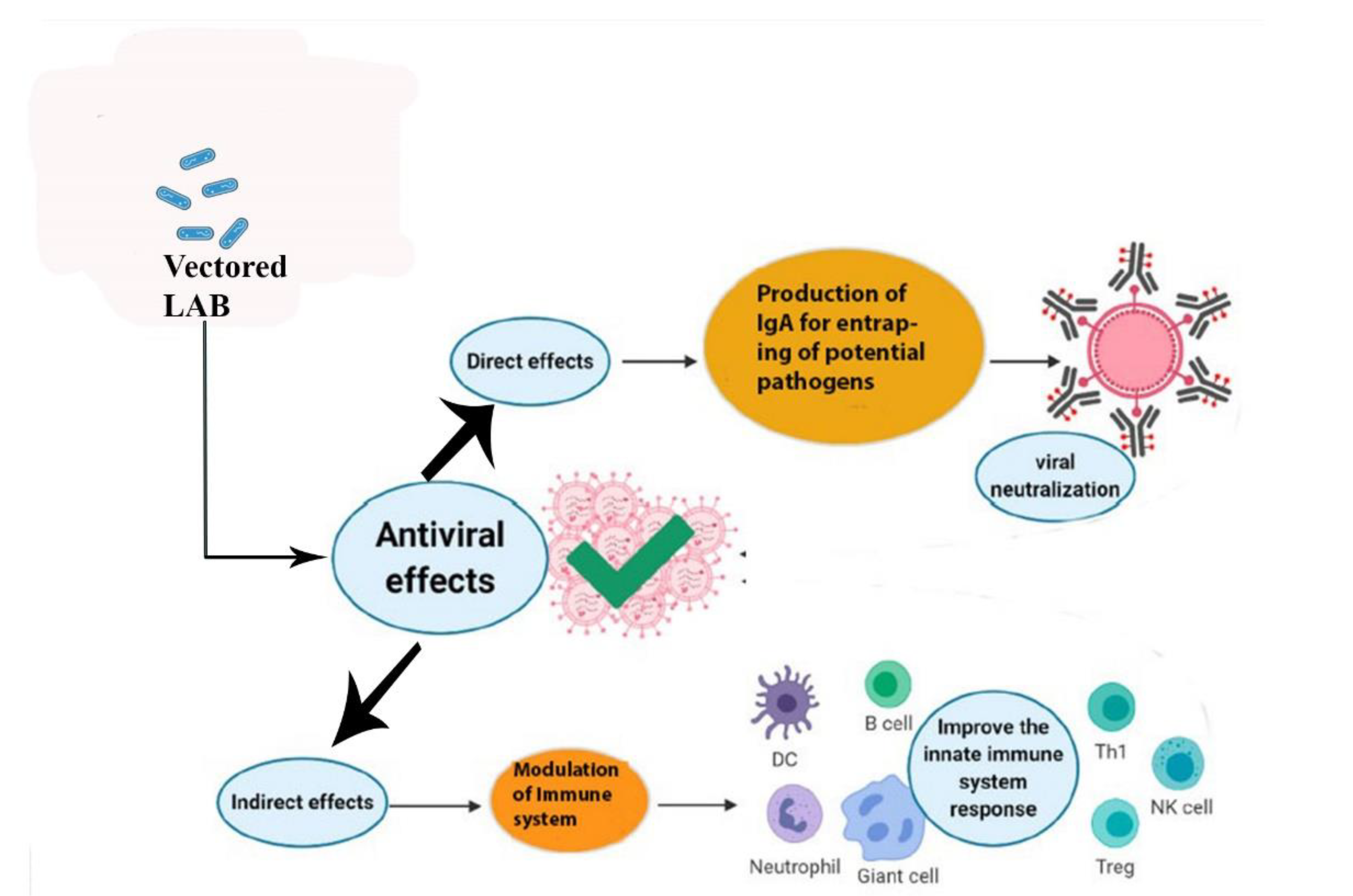
3. LAB as Antivirals
3.1. LAB-Produced EPSs as Antivirals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/Vehicle | Targeted Disease | Route | Immune Response | References |
|---|---|---|---|---|
| Enterococcus faecium L3 | Influenza virus (H1N1, H3N2) | In vitro MDCK (Madin-Darby canine kidney cell line), and in vivo (female mice). | Enterocin B stimulates IFN production and boosts the innate immune response. | [79,80] |
| L. reuteri ATCC 55730 | Enteroviruses and Coxsackie viruses CA6 and CA16 | Intestinal Caco-2 cells and in vitro (skeletal muscle RD-cell line culture). | Immune modulation of chemokines and inflammatory cytokines production. | [81] |
| L. casei DK128 | Influenza virus (H3N2 and H1N1) | In vivo administration in female mice. | Rapid induction of IgG2a and IgG1 antibodies and induction of innate immune response and production of cytokines. | [80,82] |
| L. acidophilus, L. rhamnosus(LGG) | Enteroviruses | In vitro. | Induction of IL-12 and IgA, modulation of Th1 immune response. | [80,83] |
| S thermophiles, L. plantarum | Influenza virus | In vitro. | Boost up of Th1 immune response and induction of IL-12 and IgA. | [21] |
| L. lactis | HIV-1 | In vivo by the administration in mouse through intra-gastric route with Cholera toxin as an adjuvant. | Enhanced fecal Antibodies and serum antibodies. | [84,85] |
| L. lactis | Rotavirus | Administered in mice by the intragastric route. | Production of antibodies against rotavirus infection. | [85,86] |
| L. casei | SARS-CoV | Administered in mice by intragastric and intranasal routes. | Modulation of mucosal IgA antibodies and serum antibodies. | [85,87] |
| L. lactis JCM5805 | IFV | Human | Increased expression of IFN-α and ISGs. | [88] |
| L. gasseri SBT2055 | IFV | Mouse | Increased expression of ISGs. | [89] |
| L. acidophilus | HIV-1 | Murine BALB/C | Increased Intestinal and Vaginal Epitope-Specific IgA B cells. | [90,91] |
| L. acidophilus ATCC 4356 | H9N2 | DCs | Stimulation of type-I IFNs signaling pathway. | [40] |
| L. casei | Porcine epidemic diarrhea virus (Core neutralizing epitope) | Murine BALB/C | Increased Intestinal, Vaginal, Nasal, Ocular, and Serum IgA levels. | [92] |
| L. casei | Porcine rotavirus (VP4 capsid protein) | Murine BALB/c | Increased Intestinal, Vaginal, Nasal, Ocular, and Serum IgA. | [91,93] |
| L. plantarum | Avian influenza (hemagglutinin antigen) | Murine BALB/C | Increased CD4+ T Cell IFN- (MLN), IL-4 (MLN, Splenic), IL-17 (MLN, Splenic) and CD8+ T Cell IFN-γ (MLN, Splenic). | [91,94] |
| L. lactis L. plantarum | Human papilloma virus (E7) | Murine C57BL/6 | Increased serum IgG; Increased GAL, IgA Increased IFN-γ | [91] |
| L. plantarum | NDV (Hemagglutinin-neuraminidase) | Chicken | Increased splenic and peripheral blood CD4+ T cells. | [91,95] |
3.2. LAB-Produced Peptides as Antivirals
3.3. Reactive Species Produced by LAB as Antivirals
4. LAB as Mucosal Vaccine Vectors
4.1. Suitability of LAB as Vectors
4.2. Limitations and Risks of LAB as Oral Vaccines
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Lamberte, L.E.; Van Schaik, W. Antibiotic Resistance in the Commensal Human Gut Microbiota. Cur. Opin. Microbiol. 2022, 68, 102150. [Google Scholar] [CrossRef] [PubMed]
- De Vrese, M.; Schrezenmeir. Probiotics, Prebiotics, and Synbiotics. In Food Biotechnology; Springer: Berlin/Heidelberg, Germany, 2008; Volume 111, pp. 1–66. [Google Scholar] [CrossRef]
- Singh, K.; Rao, A. Probiotics: A Potential Immunomodulator in COVID-19 Infection Management. Nutr. Res. 2021, 87, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.M.; Malcata, F.X. Bifidobacterium spp. and Lactobacillus acidophilus: Biological, Biochemical, Technological and Therapeutical Properties Relevant for Use as Probiotics. Trends Food Sci. Technol. 1999, 10, 139–157. [Google Scholar] [CrossRef]
- Arena, M.P.; Capozzi, V.; Russo, P.; Drider, D.; Spano, G.; Fiocco, D. Immunobiosis and Probiosis: Antimicrobial Activity of Lactic Acid Bacteria with a Focus on Their Antiviral and Antifungal Properties. Appl. Microbiol. Biotechnol. 2018, 102, 9949–9958. [Google Scholar] [CrossRef] [PubMed]
- Garrity, G.M.; Holt, J.G. The Road Map to the Manual. In Bergey’s Manual of Systematic Bacteriology; Springer: New York, NY, USA, 2001; pp. 119–166. [Google Scholar] [CrossRef]
- Tsuda, H.; Miyamoto, T. Production of Exo-polysaccharide by Lactobacillus plantarum and the Prebiotic Activity of Exo-polysaccharide. Food Sci. Technol. Res. 2010, 16, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Chong, E.S.L. A Potential Role of Probiotics in Colorectal Cancer Prevention: Review of Possible Mechanisms of Action. World J. Microbiol. Biotechnol. 2014, 30, 351–374. [Google Scholar] [CrossRef] [PubMed]
- Gasbarrini, G.; Bonvicini, F.; Gramenzi, A. Probiotics History. J. Clin. Gastroenterol. 2016, 50, S116–S119. [Google Scholar] [CrossRef]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. A Comprehensive Review of Anticancer, Immunomodulatory and Health Beneficial Effects of the Lactic Acid Bacteria Exo-polysaccharides. Carbhydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef]
- Toushik, S.H.; Mizan, M.F.; Hossain, M.I.; Ha, S.D. Fighting with Old Foes: The Pledge of Microbe-Derived Biological Agents to Defeat Mono and Mixed Bacterial Biofilms Concerning Food Industries. Trends Food Sci. Technol. 2020, 99, 413–425. [Google Scholar] [CrossRef]
- Nomura, R.; Matayoshi, S.; Otsugu, M.; Kitamura, T.; Teramoto, N.; Nakano, K. Contribution of Severe Dental Caries Induced by Streptococcus mutans to the Pathogenicity of Infective Endocarditis. Infect. Immun. 2020, 88, e00897-19. [Google Scholar] [CrossRef]
- Patel, S.; Majumder, A.; Goyal, A. Potentials of Exopolysaccharides from Lactic Acid Bacteria. Indian J. Microbiol. 2012, 52, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Kassaa, I. New Insights on Antiviral Probiotics; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Larsson, M.; Beignon, A.S.; Bhardwaj, N. DC-Virus Interplay: A Double-Edged Sword. Semin. Immunol. 2004, 16, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Perdigon, G. The Probiotic Bacterium Lactobacillus casei Induces Activation of the Gut Mucosal Immune System through Innate Immunity. Clin. Vaccine Immunol. 2006, 13, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toushik, S.H.; Kim, K.; Ashrafudoulla, M.; Mizan, M.F.; Roy, P.K.; Nahar, S.; Kim, Y.; Ha, S.D. Korean Kimchi Derived Lactic Acid Bacteria Inhibit Foodborne Pathogenic Biofilm Growth on Seafood and Food Processing Surface Materials. Food Contr. 2021, 129, 108276. [Google Scholar] [CrossRef]
- Sutherland, D.B.; Fagarasan, S. IgA Synthesis: A Form of Functional Immune Adaptation Extending beyond Gut. Curr. Opin. Immunol. 2012, 24, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.L.; Sahay, B.; Mohamadzadeh, M. New Generation of Oral Mucosal Vaccines Targeting Dendritic Cells. Curr. Opin. Chem. Biol. 2013, 17, 918–924. [Google Scholar] [CrossRef] [Green Version]
- Mestecky, J.; McGhee, J. Prospects for Human Mucosal Vaccines. Adv. Exp. Med. Biol. 1992, 327, 13–23. [Google Scholar] [CrossRef]
- Kawashima, T.; Hayashi, K.; Kosaka, A.; Kawashima, M.; Igarashi, T.; Tsutsui, H.; Tsuji, N.M.; Nishimura, I.; Hayashi, T.; Obata, A. Lactobacillus plantarum Strain YU from Fermented Foods Activates Th1 and Protective Immune Responses. Int. Immunopharmacol. 2011, 11, 2017–2024. [Google Scholar] [CrossRef]
- Christensen, H.R.; Frokier, H.; Pestka, J.J. Lactobacilli Differentially Modulate Expression of Cytokines and Maturation Surface Markers in Murine Dendritic Cells. J. Immunol. 2002, 168, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.Y.; Giraud, A.; Seignez, C.; Ahl, D.; Guo, F.; Sedin, J.; Walden, T.; Oh, J.H.; Van Pijkeren, J.P.; Holm, L.; et al. Distinct B Cell Subsets in Peyer’s Patches Convey Probiotic Effects by Limosilactobacillus reuteri. Microbiome 2021, 9, 198. [Google Scholar] [CrossRef]
- Peng, W.; Li, Y.H.; Yang, G.; Duan, J.L.; Yang, L.Y.; Chen, L.X.; Hou, S.L.; Huang, X.G. Oral Administration of Lactobacillus delbrueckii Enhances Intestinal Immunity through Inducing Dendritic Cell Activation in Suckling Piglets. Food Funct. 2022, 13, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Pellon, A.; Barriales, D.; Pena-Cearra, A.; Castelo-Careaga, J.; Palacios, A.; Lopez, N.; Atondo, E.; Pascual-Itoiz, M.A.; Martin-Ruiz, I.; Sampedro, L. The Commensal Bacterium Lactiplantibacillus plantarum Imprints Innate Memory-Like Responses in Mononuclear Phagocytes. Gut Microbes 2021, 13, 1939598. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, K.A.; Bennett, K.M.; Lo, D.D. Mucosal Vaccine Design and Delivery. Annu. Rev. Biomed. Eng. 2012, 14, 17–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, T.K.; Singh, M.; Dhawan, M.; Mitra, S.; Emran, T.B.; Rabaan, A.A.; Mutair, A.A.; Alawi, Z.A.; Alhumaid, S.; Dhama, K. Regulatory T Cells (Tregs) and Their Therapeutic Potential against Autoimmune Disorders Advances and Challenges. Hum. Vaccin. Immunother. 2022, 18, 2035117. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The Dendritic Cell Lineage: Ontogeny and Function of Dendritic Cells and Their Subsets in the Steady State and the Inflamed Setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varol, C.; Vallon Eberhard, A.; Elinav, E.; Aychek, T.; Shapira, Y.; Luche, H.; Fehling, H.J.; Hardt, W.D.; Shakhar, G.; Jung, S. Intestinal Lamina Propria Dendritic Cell Subsets Have Different Origins and Functions. Immunity 2009, 31, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Chinen, T.; Rudensky, A.Y. The Effects of Commensal Microbiota on Immune Cell Subsets and Inflammatory Responses. Immunol. Rev. 2012, 245, 45–55. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Chong, L.C.; Hor, Y.Y.; Lew, L.C.; Rather, I.A.; Choi, S.B. Role of Probiotics in the Management of COVID-19: A Computational Perspective. Nutrients 2022, 14, 274. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Kandasamy, S.; Chattha, K.S.; Rajashekara, G.; Saif, L.J. Comparison of Probiotic Lactobacilli and Bifidobacteria Effects, Immune Responses, and Rotavirus Vaccines and Infection in Different Host Species. Vet. Immunol. Immunopathol. 2016, 172, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Gu, C.; Hu, H.; Tang, J.; Chen, D.; Yu, B.; He, J.; Yu, J.; Luo, J.; Tian, G. Dietary Lactobacillus rhamnosus GG Supplementation Improves the Mucosal Barrier Function in the Intestine of Weaned Piglets Challenged by Porcine Rotavirus. PLoS ONE 2016, 11, e0146312. [Google Scholar] [CrossRef]
- Wu, S.; Yuan, L.; Zhang, Y.; Liu, F.; Li, G.; Wen, K.; Kocher, J.; Yang, X.; Sun, J. Probiotic Lactobacillus rhamnosus GG Mono Association Suppresses Human Rotavirus-Induced Autophagy in the Gnotobiotic Piglet Intestine. Gut Pathog. 2013, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, S.; Takeshita, M.; Kikuchi, Y.; Dashnyam, B.; Kawahara, S.; Yoshida, H.; Watanabe, W.; Muguruma, M.; Kurokawa, M. Efficacy of Oral Administration of Heat-Killed Probiotics from Mongolian Dairy Products against Influenza Infection in Mice: Alleviation of Influenza Infection by Its Immunomodulatory Activity through Intestinal Immunity. Int. Immunopharmacol. 2011, 11, 1976–1983. [Google Scholar] [CrossRef] [PubMed]
- Van Baarlen, P.; Troost, F.; Van der Meer, C.; Hooiveld, G.; Boekschoten, M.; Brummer, R.J.; Kleerebezem, M. Human Mucosal in vivo Transcriptome Responses to Three Lactobacilli Indicate How Probiotics May Modulate Human Cellular Pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 4562–4569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki Yoshida, A.; Saito, S.; Fukiya, S.; Aoki, R.; Takayama, Y.; Suzuki, C.; Sonoyama, K. Lactobacillus rhamnosus GG Increases Toll-Like Receptor 3 Gene Expression in Murine Small Intestine ex vivo and in vivo. Benef. Microbes 2016, 7, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Konstantinov, S.R.; Smidt, H.; de Vos, W.M.; Bruijns, S.C.; Singh, S.K.; Valence, F.; Molle, D.; Lortal, S.; Altermann, E.; Klaenhammer, T.R.; et al. S Layer Protein A of Lactobacillus acidophilus NCFM Regulates Immature Dendritic Cell and T Cell Functions. Proc. Natl. Acad. Sci. USA 2008, 105, 19474–19479. [Google Scholar] [CrossRef] [Green Version]
- Rice, T.A.; Brenner, T.A.; Percopo, C.M.; Ma, M.; Keicher, J.D.; Domachowske, J.B.; Rosenberg, H.F. Signaling via Pattern Recognition Receptors NOD2 And TLR2 Contributes to Immunomodulatory Control of Lethal Pneumovirus Infection. Antiviral Res. 2016, 132, 131–140. [Google Scholar] [CrossRef]
- Jeong, D.; Kim, D.H.; Kang, I.B.; Kim, H.; Song, K.Y.; Kim, H.S.; Seo, K.H. Characterization and Antibacterial Activity of a Novel Exo-polysaccharide Produced by Lactobacillus kefiranofaciens DN1 Isolated from Kefir. Food Control 2017, 78, 436–442. [Google Scholar] [CrossRef]
- Kotzamanidis, C.; Kourelis, A.; Litopoulou-Tzanetaki, E.; Tzanetakis, N.; Yiangou, M. Evaluation of Adhesion Capacity, Cell Surface Traits and Immunomodulatory Activity of Presumptive Probiotic Lactobacillus Strains. Int. J. Food Microbiol. 2010, 140, 154–163. [Google Scholar] [CrossRef]
- Weiss, G.; Rasmussen, S.; Zeuthen, L.H.; Nielsen, B.N.; Jarmer, H.; Jespersen, L.; Frokiær, H. Lactobacillus acidophilus Induces Virus Immune Defence Genes in Murine Dendritic Cells by a Toll-Like Receptor-2-Dependent Mechanism. Immunology 2010, 131, 268–281. [Google Scholar] [CrossRef]
- Kitazawa, H.; Villena, J. Modulation of Respiratory TLR3 Antiviral Response by Probiotic Microorganisms: Lessons Learned from Lactobacillus rhamnosus CRL1505. Front. Immunol. 2014, 5, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awate, S.; Babiuk, L.A.B.; Mutwiri, G. Mechanisms of Action of Adjuvants. Front. Immunol. 2013, 4, 114. [Google Scholar] [CrossRef] [Green Version]
- Albarracin, L.; Kobayashi, H.; Iida, H.; Sato, N.; Nochi, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Transcriptomic Analysis of the Innate Antiviral Immune Response in Porcine Intestinal Epithelial Cells: Influence of Immunobiotic Lactobacilli. Front. Immunol. 2017, 8, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinderola, G.; Perdigon, G.; Duarte, J.; Farnworth, E.; Matar, C. Effects of Oral Administration of the Exo-polysaccharide Produced by Lactobacillus kefiranofaciens on the Gut Mucosal Immunity. Cytokine 2006, 36, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P.; Bjerke, K.; Kett, K.; Kvale, D.; Rognum, T.; Scott, H.; Sollid, L.; Valnes, K. Production and Secretion of Immunoglobulins in the Gastrointestinal Tract. Ann. Allergy 1987, 59, 21–39. [Google Scholar]
- Villena, J.; Chiba, E.; Vizoso-Pinto, M.G.; Tomosada, Y.; Takahashi, T.; Ishizuka, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H. Immunobiotic Lactobacillus rhamnosus Strains Differentially Modulate Antiviral Immune Response in Porcine Intestinal Epithelial and Antigen-Presenting Cells. BMC Microbiol. 2014, 14, 126. [Google Scholar] [CrossRef] [Green Version]
- Bernal, P.; Llamas, M.A. Promising Biotechnological Applications of Antibiofilm Exopolysaccharides. Microb. Biotechnol. 2012, 5, 670–673. [Google Scholar] [CrossRef] [Green Version]
- Abedfar, A.; Hossininezhad, M. Overview of The Most Important Characterization of Exopolysaccharides Produced by Probiotics Bacteria and Their Biological Function. J. Environ. Sci. Toxicol. Food Technol. 2016, 10, 47–55. [Google Scholar] [CrossRef]
- Deepak, V.; Ram Kumar Pandian, S.; Sivasubramaniam, S.D.; Nellaiah, H.; Sundar, K. Optimization of Anticancer Exopolysaccharide Production from Probiotic Lactobacillus acidophilus By Response Surface Methodology. Prep. Biochem. Biotechnol. 2016, 46, 288–297. [Google Scholar] [CrossRef]
- Liu, C.T.; Chu, F.J.; Chou, C.C.; Yu, R.C. Antiproliferative and Anticytotoxic Effects of Cell Fractions and Exo-polysaccharides from Lactobacillus casei 01. Mutat. Res. 2011, 721, 157–162. [Google Scholar] [CrossRef]
- Castellone, V.; Bancalari, E.; Rubert, J.; Gatti, M.; Neviani, E.; Bottari, B. Eating Fermented: Health Benefits of LAB-Fermented Foods. Foods 2021, 10, 2639. [Google Scholar] [CrossRef] [PubMed]
- Vahed, S.Z.; Barzegari, A.; Saadat, Y.R.; Goreyshi, A.; Omidi, Y. Leuconostoc Mesenteroides Derived Anticancer Pharmaceuticals Hinder Inflammation and Cell Survival in Colon Cancer Cells by Modulating NF-κB/AKT/PTEN/MAPK Pathways. Biomed. Pharmacother. 2017, 94, 1094–1100. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. P Production, Properties, and Industrial Food Application of Lactic Acid Bacteria-Derived Exo-polysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- West, T. Effect of Temperature on Bacterial Gellan Production. World J. Microbiol. Biotechnol. 2003, 19, 649–652. [Google Scholar] [CrossRef]
- Daba, G.M.; Elnahas, M.O.; Elkhateeb, W.A. Contributions of Exo-polysaccharides from Lactic Acid Bacteria as Biotechnological Tools in Food, Pharmaceutical, and Medical Applications. Int. J. Biol. Macromol. 2021, 173, 79–89. [Google Scholar] [CrossRef]
- Ogel, Z.B.; Ozturk, H.I. Antiviral Mechanisms Related to Lactic Acid Bacteria and Fermented Food Products. Biotechnol. Studies 2020, 29, 18–28. [Google Scholar] [CrossRef]
- Alsaadi, L.G.; Baker, B.A.; Kadhem, B.M.; Mahdi, L.H.; Mater, H.N. Exopolysaccharide as Antiviral, Antimicrobial and as Immunostimulants: A Review. Plant Arch. 2020, 20, 5859–5875. [Google Scholar]
- Mousavi, E.; Makvandi, M.; Teimoori, A.; Ataei, A.; Ghafari, S.; Samarbaf-Zadeh, A. Antiviral Effects of Lactobacillus crispatus against HSV-2 in Mammalian Cell Lines. J. Chin. Med. Assoc. 2018, 81, 262–267. [Google Scholar] [CrossRef]
- Abdalla, A.K.; Ayyash, M.M.; Olaimat, A.N.; Osaili, T.M.; Al Nabulsi, A.A.; Shah, N.P.; Holley, R. Exopolysaccharides as Antimicrobial Agents: Mechanism and Spectrum of Activity. Front. Microbiol. 2021, 12, 664395. [Google Scholar] [CrossRef]
- Xiao, H.; Killip, M.J.; Staeheli, P.; Randall, R.E.; Jackson, D. The Human Interferon-Induced MxA Protein Inhibits Early Stages of Influenza A Virus Infection by Retaining the Incoming Viral Genome in the Cytoplasm. J. Virol. 2013, 87, 13053–13058. [Google Scholar] [CrossRef] [Green Version]
- Botic, T.; Dano, T.; Weingartl, H.; Cencic, A. A Novel Eukaryotic Cell Culture Model to Study Antiviral Activity of Potential Probiotic Bacteria. Int. J. Food Microbiol. 2007, 115, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Al Kassaa, I.; Hober, D.; Hamze, M.; Chihib, N.E.; Drider, D. Antiviral Potential of Lactic Acid Bacteria and their Bacteriocins. Probiotics Antimicrob. Proteins 2014, 6, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.A.; Sheikh, F.N.; Jamal, S.; Ezeh, J.K.; Akhtar, A. Coronavirus (COVID-19): A Review of Clinical Features, Diagnosis, and Treatment. Cureus 2020, 12, e7355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rautava, S.; Salminen, S.; Isolauri, E. Specific Probiotics in Reducing the Risk of Acute Infections in Infancy: A Randomised, Double-blind, Placebo-Controlled Study. Br. J. Nutr. 2008, 101, 1722–1726. [Google Scholar] [CrossRef]
- King, S.; Tancredi, D.; Lenoir Wijnkoop, I.; Gould, K.; Vann, H.; Connors, G.; Sanders, M.E.; Linder, J.A.; Shane, A.L.; Merenstein, D. Does Probiotic Consumption Reduce Antibiotic Utilization for Common Acute Infections: A Systematic Review and Meta-Analysis. Eur. J. Public Health 2019, 29, 494–499. [Google Scholar] [CrossRef]
- Antunes, A.E.; Vinderola, G.; Xavier-Santos, D.; Sivieri, K. Potential Contribution of Beneficial Microbes to Face the COVID-19 Pandemic. Food Res. Intl. 2020, 136, 109577. [Google Scholar] [CrossRef]
- Khaneghah, A.M.; Abhari, K.; Eş, I.; Soares, M.B.; Oliveira, R.B.; Hosseini, H.; Rezaei, M.; Balthazar, C.F.; Silva, R.; Cruz, A.G.; et al. Interactions between Probiotics and Pathogenic Microorganisms in Hosts and Foods: A Review. Trends Food Sci. Technol. 2020, 95, 205–218. [Google Scholar] [CrossRef]
- Montazeri-Najafabady, N.; Kazemi, K.; Gholami, A. Recent Advances in Antiviral Effects of Probiotics: Potential Mechanism Study in Prevention and Treatment of SARS-CoV-2. Biologia 2022, 1–18. [Google Scholar] [CrossRef]
- Bottari, B.; Castellone, V.; Neviani, E. Probiotics and COVID-19. Int. J. Food Sci. Nutr. 2021, 72, 293–299. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological Findings of COVID-19 Associated with Acute Respiratory Distress Syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Baud, D.; Agri, V.D.; Gibson, G.R.; Reid, G.; Giannoni, E. Using Probiotics to Flatten the Curve of Coronavirus Disease COVID-2019 Pandemic. Front. Public Health 2020, 8, 186. [Google Scholar] [CrossRef] [PubMed]
- Starosila, D.; Rybalko, S.; Varbanetz, L.; Ivanskaya, N.; Sorokulova, I. Anti-Influenza Activity of Bacillus subtilis Probiotic Strain. Antimicrob. Agents Chemother. 2017, 61, e00539-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottari, B.; Quartieri, A.; Prandi, B.; Raimondi, S.; Leonardi, A.; Rossi, M.; Ulrici, A.; Gatti, M.; Sforza, S.; Nocetti, M.; et al. Characterization of Peptide Fraction from Digested Parmigiano Reggiano Cheese and Its Effect on Growth of Lactobacilli and Bifidobacteria. Int. J. Food Microbiol. 2017, 255, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.; Chattopadhyay, K.; Marschall, M.; Karmakar, P.; Mandal, P.; Ray, B. Focus on Antivirally Active Sulfated Polysaccharides: From Structure Activity Analysis to Clinical Evaluation. Glycobiology 2009, 19, 2–15. [Google Scholar] [CrossRef]
- Angelin, J.; Kavitha, M. Exo-polysaccharides from Probiotic Bacteria and Their Health Potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Ermolenko, E.; Desheva, Y.; Kolobov, A.; Kotyleva, M.; Sychev, I.; Suvorov, A. Anti- Influenza Activity of Enterocin B in vitro and Protective Effect of Bacteriocinogenic Enterococcal Probiotic Strain on Influenza Infection in Mouse Model. Probiotics Antimicrob. Proteins 2019, 11, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Muhialdin, B.J.; Zawawi, N.; Razis, A.F.; Bakar, J.; Zarei, M. Antiviral Activity of Fermented Foods and Their Probiotic Bacteria towards Respiratory and Alimentary Tracts Viruses. Food Control 2021, 127, 108140. [Google Scholar] [CrossRef]
- Ang, L.; Too, H.; Tan, E.; Chow, T.; Shek, P.; Tham, E.; Alonso, S. Antiviral Activity of Lactobacillus Reuteri Protects against Coxsackievirus A and Enterovirus 71 Infection in Human Skeletal Muscle and Colon Cell Lines. Virol. J. 2016, 13, 111. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.-J.; Lee, Y.-T.; Le Ngo, V.; Cho, Y.-H.; Ko, E.-J.; Hong, S.-M.; Kim, K.-H.; Jang, J.-H.; Oh, J.-S.; Park, M.-K.; et al. Heat-Killed Lactobacillus casei Confers Broad Protection against Influenza A Virus Primary Infection and Develops Heterosubtypic Immunity against Future Secondary Infection. Sci. Rep. 2017, 7, 17360. [Google Scholar] [CrossRef]
- Choi, H.J.; Song, J.H.; Ahn, Y.J.; Baek, S.H.; Kwon, D.H. Antiviral Activities of Cell-Free Supernatants of Yogurts Metabolites against Some RNA Viruses. Eur. Food Res. Technol. 2009, 228, 945–950. [Google Scholar] [CrossRef]
- Xin, K.Q.; Hoshino, Y.; Toda, Y.; Igimi, S.; Kojima, Y.; Jounai, N.; Ohba, K.; Kushiro, A.; Kiwaki, M.; Hamajima, K.; et al. Immunogenicity and Protective Efficacy of Orally Administered Recombinant Lactococcus lactis Expressing Surface-Bound HIV Env. Blood 2003, 102, 223–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, J.M.; Mercenier, A. Mucosal Delivery of Therapeutic and Prophylactic Molecules Using Lactic Acid Bacteria. Nat. Rev. Microbiol. 2008, 6, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Perez, C.; Eichwald, C.; Burrone, O.; De Mendoza, D. Rotavirus VP7 Antigen Produced by Lactococcus lactis Induces Neutralizing Antibodies in Mice. J. Appl. Microbiol. 2005, 99, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Poo, H.; Han, D.P.; Hong, S.P.; Kim, K.; Cho, M.W.; Kim, E.; Sung, M.H.; Kim, C.J. Mucosal Immunization with Surface-Displayed Severe Acute Respiratory Syndrome Coronavirus Spike Protein on Lactobacillus casei Induces Neutralizing Antibodies in Mice. J. Virol. 2006, 80, 4079–4087. [Google Scholar] [CrossRef] [Green Version]
- Sugimura, T.; Takahashi, H.; Jounai, K.; Ohshio, K.; Kanayama, M.; Tazumi, K. Effects of Oral Intake of pDC Stimulative Lactic Acid Bacterial Strain on Pathogenesis of Influenza-Like Illness and Immunological Response to Influenza Virus. Br. J. Nutr. 2015, 114, 727–733. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, K.; Fujitani, N.; Nakagawa, H.; Miyazaki, T. Prevention of Respiratory Syncytial Virus Infection with Probiotic Lactic Acid Bacterium Lactobacillus gasseri SBT2055. Sci. Rep. 2019, 9, 4812. [Google Scholar] [CrossRef] [Green Version]
- Kajikawa, A.; Zhang, L.; LaVoy, A.; Bumgardner, S.; Klaenhammer, T.R.; Dean, G.A. Mucosal immunogenicity of genetically modified Lactobacillus acidophilus expressing an HIV-1 epitope within the surface layer protein. PLoS ONE 2015, 10, e0141713. [Google Scholar] [CrossRef]
- Vilander, A.C.; Dean, G.A. Adjuvant strategies for lactic acid bacterial mucosal vaccines. Vaccines 2019, 7, 150. [Google Scholar] [CrossRef] [Green Version]
- Ge, J.W.; Liu, D.Q.; Li, Y.J. Construction of Recombinant Lactobacilli Expressing the Core Neutralizing Epitope (COE) of Porcine Epidemic Diarrhea Virus and a Fusion Protein Consisting of COE and Escherichia coli Heat-Labile Enterotoxin B, and Comparison of the Immune Responses by Orogastric Immunization. Can. J. Microbiol. 2012, 58, 1258–1267. [Google Scholar]
- Indo, Y.; Kitahara, S.; Tomokiyo, M.; Araki, S.; Islam, M.A.; Zhou, B.; Albarracin, L.; Miyazaki, A.; Ikeda Ohtsubo, W.; Nochi, T.; et al. Ligilactobacillus salivarius Strains Isolated from the Porcine Gut Modulate Innate Immune Responses in Epithelial Cells and Improve Protection against Intestinal Viral Bacterial Superinfection. Front. Immunol. 2021, 12, 652923. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, G.; Wang, Q.; Wang, Z.; Yang, W.; Gu, W.; Shi, C.; Wang, J.; Huang, H.; Wang, C. Molecular Mechanisms Underlying Protection Against H9N2 Influenza Virus Challenge in Mice by Recombinant Lactobacillus plantarum with Surface Displayed HA2-LTB. J. Biotechnol. 2017, 259, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Hu, J.; Guo, Y.; Yang, W.; Ye, L.; Shi, C.; Liu, Y.; Yang, G.; Wang, C. Construction and Immunological Evaluation of Recombinant Lactobacillus plantarum Expressing HN of Newcastle Disease Virus and DC-Targeting Peptide Fusion Protein. J. Biotechnol. 2015, 216, 82–89. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini Review on Antimicrobial Peptides, Sources, Mechanism and Recent Applications. Protein Pept. Lett. 2020, 27, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Patch, J.A.; Barron, A.E. Mimicry of Bioactive Peptides via Non-natural, Sequence-specific Peptidomimetic Oligomers. Cur. Opin. Chem. Biol. 2002, 6, 872–877. [Google Scholar] [CrossRef]
- Bosch, T.C.; Zasloff, M. Antimicrobial Peptides or How Our Ancestors Learned to Control the Microbiome. mBio 2021, 12, e01847-21. [Google Scholar] [CrossRef] [PubMed]
- Wachsman, M.B.; Castilla, V.; de Ruiz Holgado, A.P.; De Torres, R.A.; Sesma, F.; Coto, C.E. Enterocin CRL35 Inhibits Late Stages of HSV-1 and HSV-2 Replication in vitro. Antivir. Res. 2003, 58, 17–24. [Google Scholar] [CrossRef]
- Serkedjieva, J.; Danova, S.; Ivanova, I. Antiinfluenza Virus Activity of a Bacteriocin Produced by Lactobacillus delbrueckii. App. Biochem. Biotechnol. 2000, 88, 285–298. [Google Scholar] [CrossRef]
- Hill, J.A.; Anderson, D.J. Human Vaginal Leukocytes and the Effects of Vaginal Fluid on Lymphocyte and Macrophage Defense Functions. Am. J. Obstet. Gynecol. 1992, 166, 720–726. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface Components and Metabolites of Probiotics for Regulation of Intestinal Epithelial Barrier. Microb. Cell Fact. 2020, 19, 23. [Google Scholar] [CrossRef] [Green Version]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. Vaginal Fluid, Bacteria Associated with Bacterial Vaginosis Can Be Suppressed with Lactic Acid but Not Hydrogen Peroxide. BMC Infect. Dis. 2011, 11, 200. [Google Scholar] [CrossRef]
- Nahui Palomino, R.A.; Zicari, S.; Vanpouille, C.; Vitali, B.; Margolis, L. Vaginal Lactobacillus Inhibits HIV-1 Replication in Human Tissues ex vivo. Front. Microbiol. 2017, 8, 906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyssen, D.; Wang, Y.; Hayward, J.; Agius, P.; DeLong, K.; Aldunate, M. Anti-HIV-1 Activity of Lactic Acid in Human Cervicovaginal Fluid. mSphere 2018, 3, e00055-18. [Google Scholar] [CrossRef] [Green Version]
- Gosmann, C.; Anahtar, M.N.; Handley, S.A.; Farcasanu, M.; Abu-Ali, G.; Bowman, B.A.; Padavattan, N.; Desai, C.; Droit, L.; Moodley, A.; et al. Lactobacillus Deficient Cervicovaginal Bacterial Communities Are Associated with Increased HIV Acquisition in Young South African Women. Immunity 2017, 46, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanderWaal, K.; Deen, J. Global Trends in Infectious Diseases of Swine. Proc. Natl. Acad. Sci. USA 2018, 115, 11495–11500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliavia, M.; Nicosia, A. Advanced Strategies for Food Grade Protein Production: A New E. coli Lactic Acid Bacteria Shuttle Vector for Improved Cloning and Food Grade Expression. Microorganisms 2019, 7, 116. [Google Scholar] [CrossRef] [Green Version]
- Mercenier, A.; Müller-Alouf, H.; Grangette, C. Lactic Acid Bacteria as Live Vaccines. Curr. Issues Mol. Biol. 2000, 2, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Szatraj, K.; Szczepankowska, A.K.; Chmielewska-Jenza, M. Lactic Acid Bacteria Promising Vaccine Vectors: Possibilities, Limitations, and Doubts. J. Appl. Microbiol. 2017, 123, 325–339. [Google Scholar] [CrossRef] [Green Version]
- Wells, J. Mucosal Vaccination and Therapy with Genetically Modified Lactic Acid Bacteria. Annu. Rev. Food Science Technol. 2011, 2, 423–445. [Google Scholar] [CrossRef]
- Pontes, D.S.; De Azevedo, M.S.; Chatel, J.M.; Langella, P.; Azevedo, V.; Miyoshi, A. Lactococcus Lactis a Live Vector: Heterologous Protein Production and DNA Delivery Systems. Protein Expr. Purif. 2011, 79, 165–175. [Google Scholar] [CrossRef]
- Zhang, J.; Rodríguez, F.; Navas, M.J.; Costa Hurtado, M.; Almagro, V.; Bosch Camos, L.; Lopoz, E.; Cuadrado, R.; Accensi, F.; Pina Pedrero, S.; et al. Fecal Microbiota Transplantation from Warthog to Pig Confirms the Influence of the Gut Microbiota on African Swine Fever Susceptibility. Sci. Rep. 2020, 10, 17605. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, W.; Sun, Z.; Zhu, C.; Werid, G.M.; Ibrahim, Y.M.; Zhang, W.; Pan, Y.; Shi, D.; Chen, H.; et al. Abundance of Lactobacillus in Porcine Gut Microbiota Is Closely Related to Immune Response Following PRRSV Immunization. Vet. Microbiol. 2021, 259, 109134. [Google Scholar] [CrossRef] [PubMed]
- Morita, N.; Umemoto, E.; Fujita, S.; Hayashi, A.; Kikuta, J.; Kimura, I.; Haneda, T.; Imai, T.; Inoue, A.; Mimuro, H.; et al. GPR31-Dependent Dendrite Protrusion of Intestinal CX3CR1+ Cells by Bacterial Metabolites. Nature 2019, 566, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Van Baarlen, P.; Wells, J.M.; Kleerebezem, M. Regulation of Intestinal Homeostasis and Immunity with Probiotic Lactobacilli. Trends Immunol. 2013, 34, 208–215. [Google Scholar] [CrossRef]
- Oh, S.H.; Kim, S.H.; Jeon, J.H.; Kim, E.B.; Lee, N.K.; Beck, S.; Choi, Y.J.; Kang, S.K. Cytoplasmic Expression of Model Antigen with M Cell Targeting Moiety in Lactic Acid Bacteria and Implication of the Mechanism as a Mucosal Vaccine via Oral Route. Vaccine 2021, 39, 4072–4081. [Google Scholar] [CrossRef] [PubMed]




| EPSs | Chemical Composition | Producer Strain | Application | References |
|---|---|---|---|---|
| HoPSs | ||||
| Mutan | α-1,3 glycosidic linkage of the monomers of glucose. | Streptococcus mutans, Leuconostoc, and Lactobacillus species | Water-soluble HoPSs used as a starter culture. | [55] |
| Dextran | α-1,6 glycosidic linkages of the monomer of glucose. | Leuconostoc, Lactobacillus, and Streptococcus species | As an adjuvant, emulsifier, anti-coagulant, blood flow enhancer, and cholesterol-lowering agent. | [13] |
| Reuteran | α-1,4 and α-1-6 glycosidic linkage of the monomer of glucose. | Lactobaciluus reuteri | Water-soluble HoPSs used in bakery. | [13] |
| Alternan | α-1,6 and α-1,3 glycosidic linkage of the monomer of glucose. | Lactobacillus mesenteriodes | Highly water-soluble, low viscosity, and very less resistant to being hydrolyzed, used as probiotics. | [56] |
| Levan | β-2,6 and β-2,1 linkage of the monomer of fructose. | Streptococcus salivarius, L. mesenteroides, L. sanfranciscensis, and S. mutans | Cholesterol-lowering properties, natural adhesives, anti-tumorous effects, and are non-toxic to animals and humans. | [10,56] |
| Inulin | β-1,2 glycosidic linkage of the monomer of fructose. | S. mutans JC2, Leuconostoc citreum, L. reuteri, and Lactobacillus johnsoni | Indigestible, probiotics, prevent the attachment of viruses by reducing gastrointestinal wall pH, use as a vehicle for targeted drugs in colon carcinoma, and substitute for fat in food products. | [10,56] |
| HePSs | ||||
| Keiferan | An equal proportion of glucose and galactose in linkage. | Lactobaccilus kefiranum, Lactobacillus parakefir, and L. beijerinck, L. kefiri | Wound healing, cholesterol-lowering medicines, anti-hypertensive drugs, tumor growth retardation medications, and enhancing mucosal immunity by IgA production. | [10,56] |
| Gellan | A tetra-saccharide structure is composed of 60% glucose, 20% rhamnose, and 20% glucuronic acid. | Sphingomonas paucimobilis ATCC 31461 | Use as a thickening agent or an adhesive in foods and as an agar substitute. | [57] |
| Xanthan | The backbone is made up of glucose and side chains consist of mannose-glucuronic acid-mannose. | Xanthomonas campestris | Moisture retention in wound dressings, suspension agents, and emulsion stabilizers in the food industry. | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, A.; Sun, Y.; Wang, Y.; Huang, J.; Zafar Khan, M.U.; Qiu, H.-J. Lactic Acid Bacteria as Mucosal Immunity Enhancers and Antivirals through Oral Delivery. Appl. Microbiol. 2022, 2, 837-854. https://doi.org/10.3390/applmicrobiol2040064
Moon A, Sun Y, Wang Y, Huang J, Zafar Khan MU, Qiu H-J. Lactic Acid Bacteria as Mucosal Immunity Enhancers and Antivirals through Oral Delivery. Applied Microbiology. 2022; 2(4):837-854. https://doi.org/10.3390/applmicrobiol2040064
Chicago/Turabian StyleMoon, Assad, Yuan Sun, Yanjin Wang, Jingshan Huang, Muhammad Umar Zafar Khan, and Hua-Ji Qiu. 2022. "Lactic Acid Bacteria as Mucosal Immunity Enhancers and Antivirals through Oral Delivery" Applied Microbiology 2, no. 4: 837-854. https://doi.org/10.3390/applmicrobiol2040064