
Responses to Ice Formation and Reasons of Frost Injury in Potato Leaves

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (1)
- After E1, mesophyll cells of potato leaves show freeze-dehydration;
- (2)
- After E1, although it is non-injurious, gas exchange activity stops as ice formation in the intercellular spaces between mesophyll cells blocks gas diffusion;
- (3)
- During E2, the mesophyll cells freeze intracellularly, which causes membrane rupture and irreversibly destroys the cells, resulting in lethality.
2. Material and Methods
2.1. Plant Material
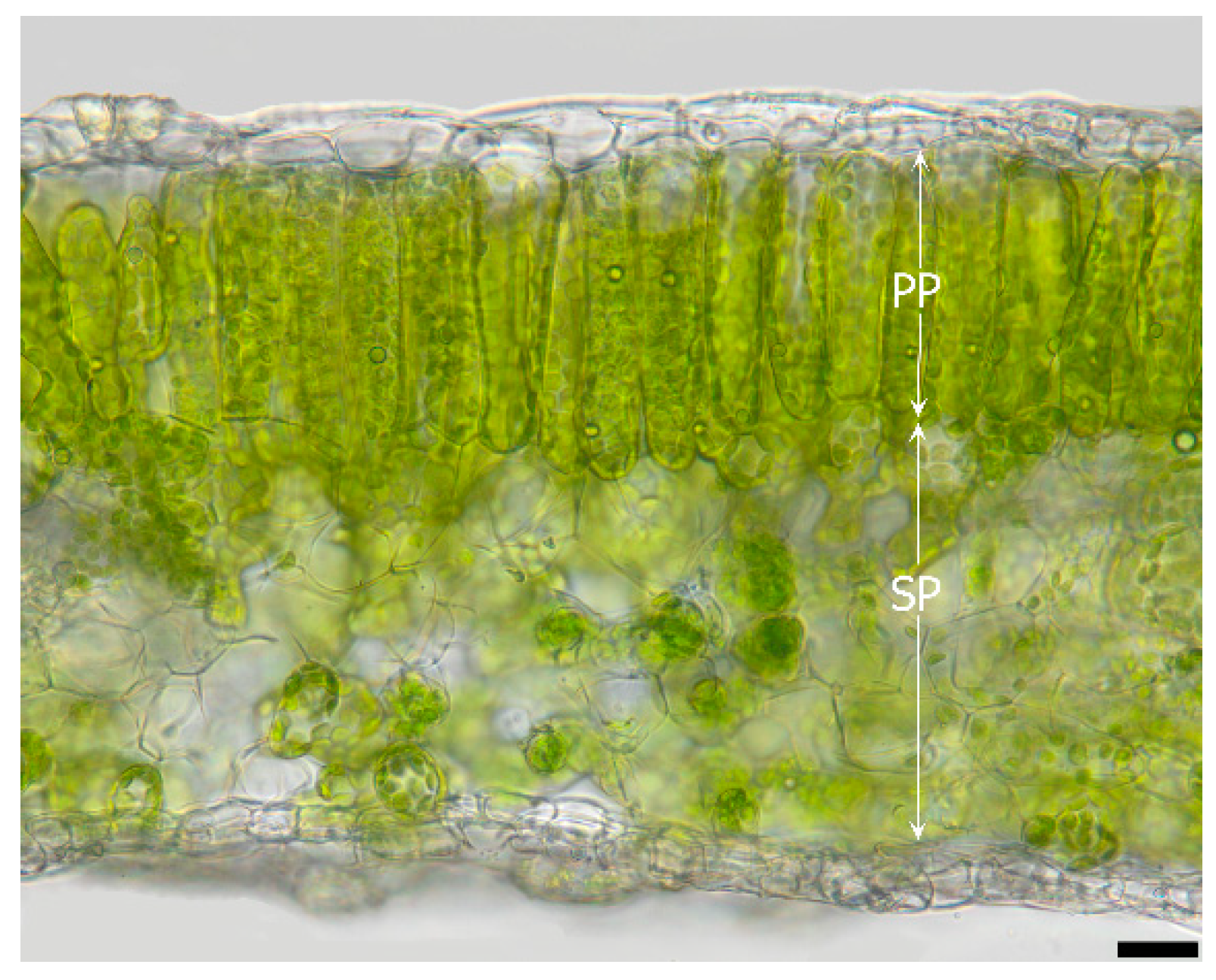
2.2. Leaf Anatomy
2.3. Freezing Treatments
2.4. Effects of Freezing Events on CO2 Gas Exchange
2.5. Cryo-Microscopy
2.5.1. Semi-Thin Sections of High-Pressure Frozen Leaves
2.5.2. Cross-Sections Viewed by Light Microscope
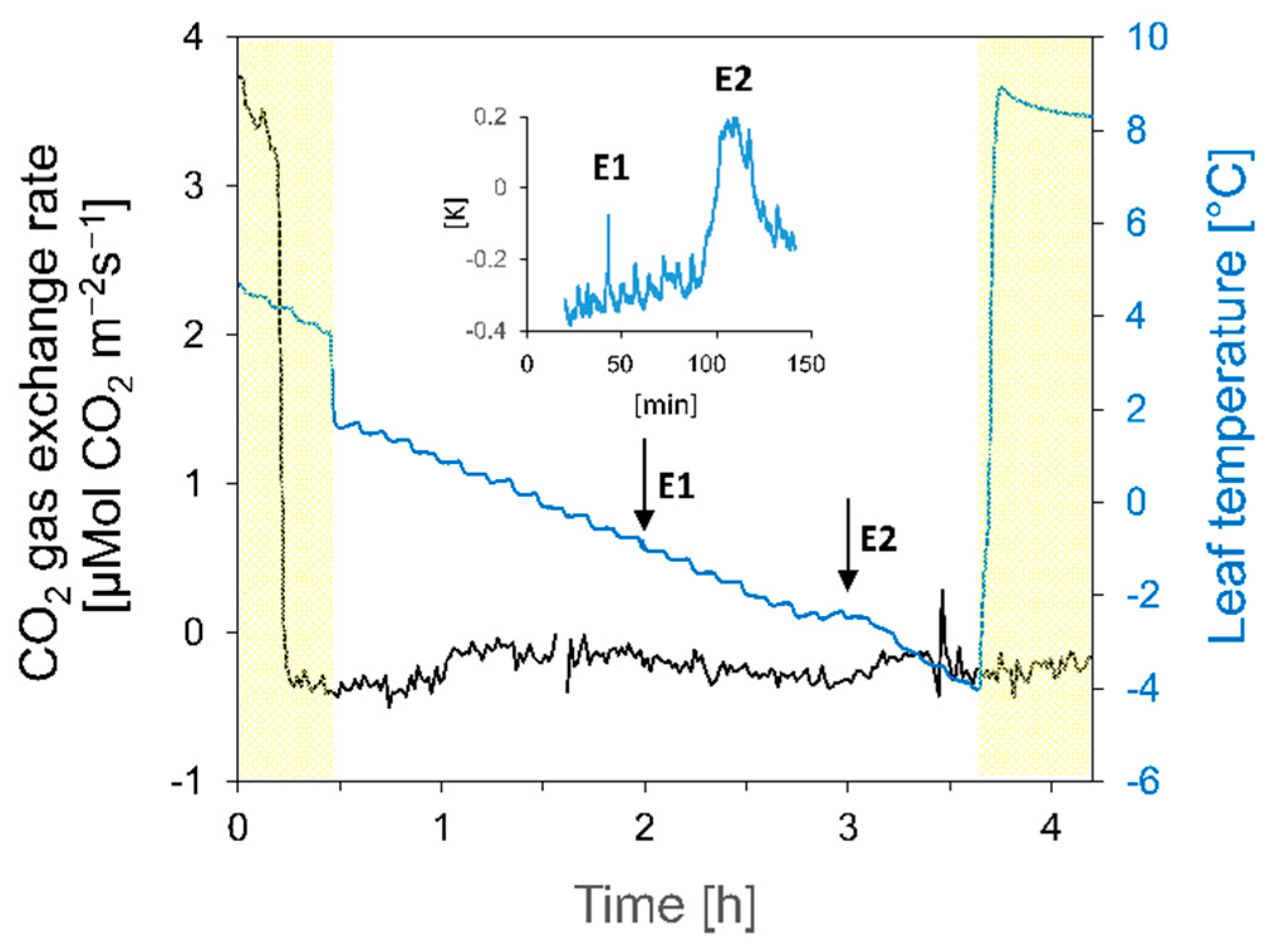
3. Results
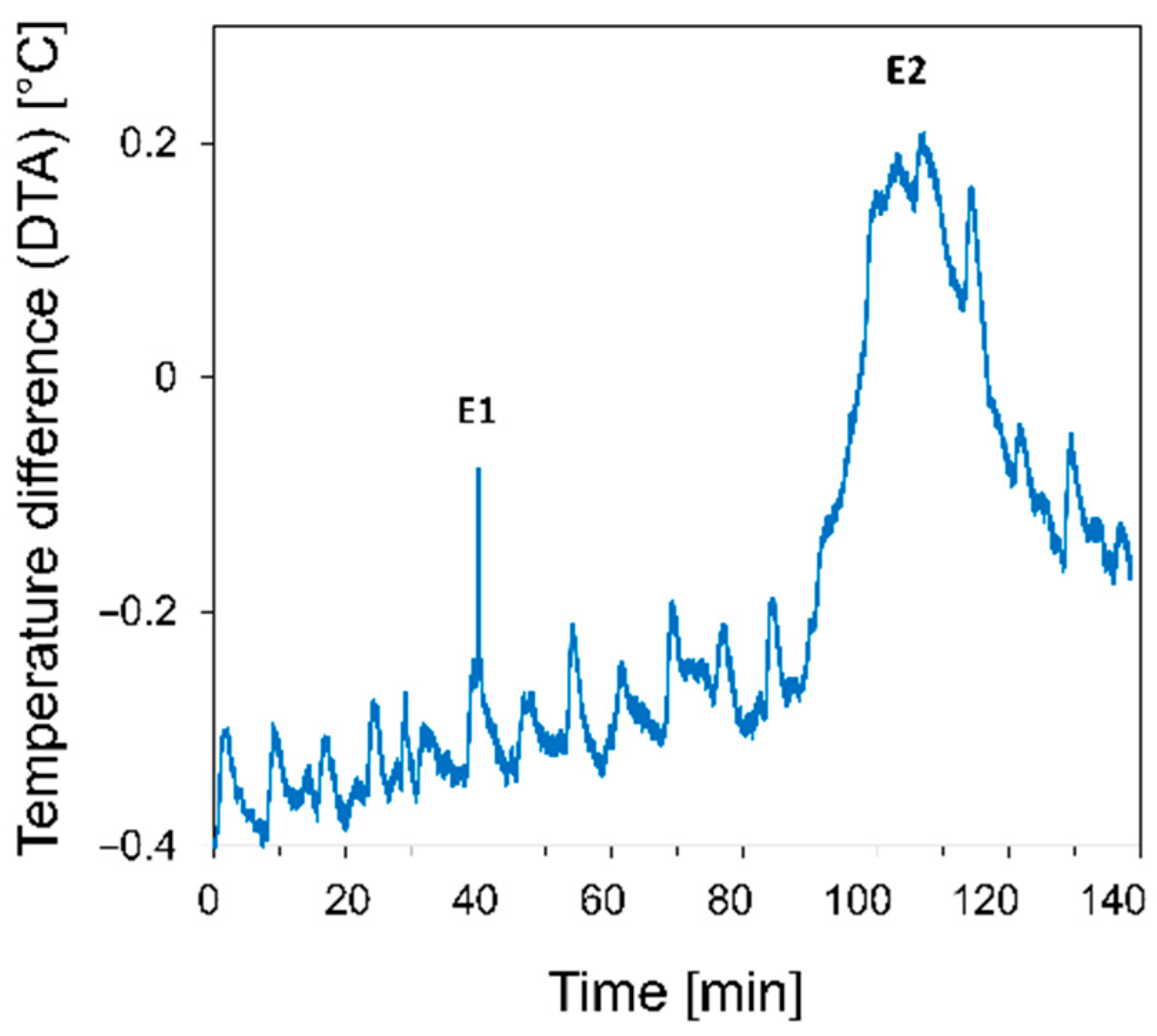
3.1. Responses to the First Freezing Event
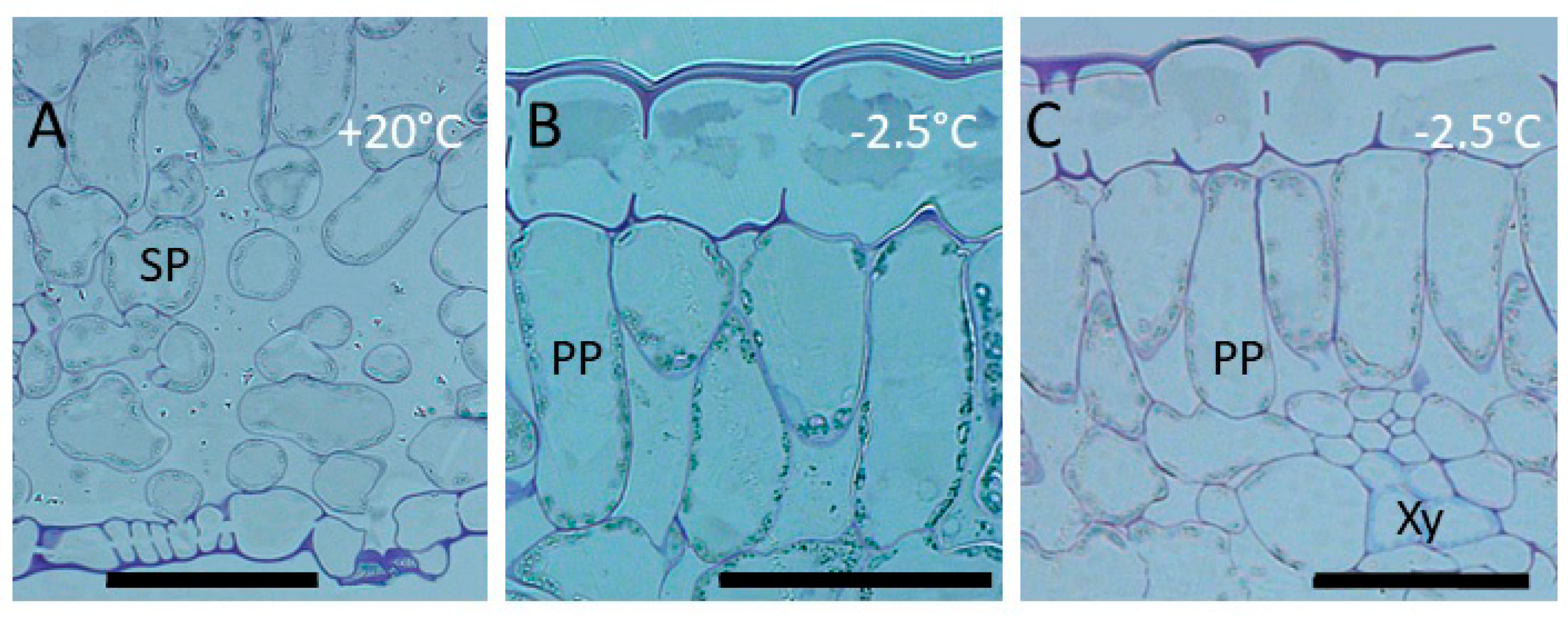
3.1.1. Cytological Aspects
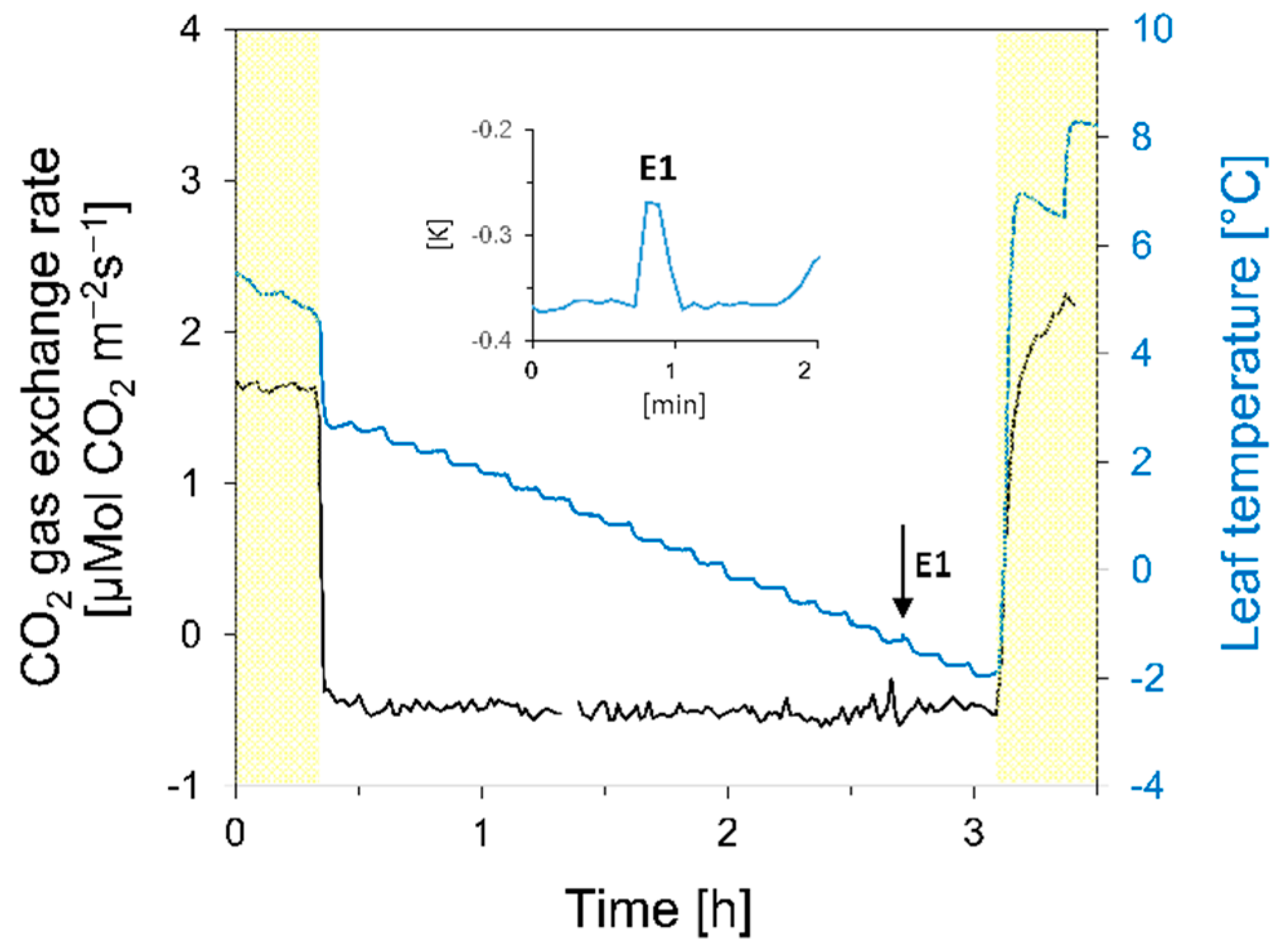
3.1.2. Gas Exchange and Photosystem II
3.2. Responses to the Second Freezing Event
3.2.1. Cytological Aspects
3.2.2. Gas Exchange and Photosystem II
4. Discussion
4.1. Localization of Ice and Cellular Responses to the First Freezing Event
4.2. The Second Freezing Event Is Intracellular
4.3. Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reddy, B.J.; Mandal, R.; Chakroborty, M.; Hijam, L.; Dutta, P. A review on potato (Solanum tuberosum L.) and its genetic diversity. Genetics 2018, 10, 360–364. [Google Scholar] [CrossRef]
- Glendinning, D.R. Potato introductions and breeding up to the early 20th century. New Phytol. 1983, 94, 479–505. [Google Scholar] [CrossRef]
- Gu, L.; Hanson, P.J.; Mac Post, W.; Kaiser, D.P.; Yang, B.; Nemani, R.; Pallardy, S.G.; Meyers, T. The 2007 eastern US spring freezes: Increased cold damage in a warming world? Bioscience 2008, 58, 253–262. [Google Scholar] [CrossRef]
- Richardson, D.G.; Weiser, C.J. Foliage frost resistance in tuber-bearing Solanums. HortScience 1972, 7, 19–22. [Google Scholar] [CrossRef]
- Li, H.; Luo, W.; Ji, R.; Xu, Y.; Xu, G.; Qiu, S.; Tang, H. A comparative proteomic study of cold responses in potato leaves. Heliyon 2021, 7, e06002. [Google Scholar] [CrossRef]
- Arora, R. Mechanism of freeze-thaw injury and recovery: A cool retrospective and warming up to new ideas. Plant Sci. 2018, 270, 301–313. [Google Scholar] [CrossRef]
- Palta, J.P.; Weiss, L.S.; Harbage, J.F.; Bamberg, J.B.; Stone, J.M. Molecular mechanisms of freeze-thaw injury and cold acclimation in herbaceous plants: Merging physiological and genetic approaches. In Interacting Stresses on Plants in a Changing Climate; NATO ASI Series; Jackson, M.B., Black, C.R., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; Volume 16, pp. 659–680. [Google Scholar]
- Chen, P.M.; Burke, M.J.; Li, P.H. Frost hardiness of several solanum species in relation to freezing of water, melting-point depression, and tissue water content. Bot. Gaz. 1976, 137, 313–317. [Google Scholar] [CrossRef]
- Stegner, M.; Schäfernolte, T.; Neuner, G. New insights in potato leaf freezing by infrared thermography. Appl. Sci. 2019, 9, 819. [Google Scholar] [CrossRef] [Green Version]
- Li, P.H.; Huner, N.P.A.; Toivio-Kinnucan, M.; Chen, H.H.; Palta, J.P. Potato freezing-injury and survival, and their relationships to other stress. AJPR 1981, 58, 15–29. [Google Scholar] [CrossRef]
- Chen, H.-H.; Li, P.H. Characteristics of cold acclimation and deacclimation in tuber-bearing Solanum species. Plant Physiol. 1980, 65, 1146–1148. [Google Scholar] [CrossRef]
- Steffen, K.L.; Arora, R.; Palta, J.P. Relative sensitivity of photosynthesis and respiration to freeze-thaw stress in herbaceous species: Importance of realistic freeze-thaw protocols. Plant Physiol. 1989, 89, 1372–1379. [Google Scholar] [CrossRef] [Green Version]
- Larcher, W. Physiological plant ecology. In Ecophysiology and Stress Physiology of Functional Groups; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Levitt, J. Responses of plants to environmental stresses. In Chilling, Freezing, and High Temperature Stresses; Academic Press: London, UK; New York, NY, USA, 1980. [Google Scholar]
- Asahina, E. The freezing process of plant cells. Contrib. Inst. Low Temp. Sci. 1956, 10, 83–126. [Google Scholar]
- Neuner, G.; Hacker, J. Ice formation and propagation in alpine plants. In Plants in Alpine Regions: Cell Physiology of Adaptation and Survival Strategies; Lütz, C., Ed.; Springer: Wien, Austria; New York, NY, USA, 2012; pp. 163–174. [Google Scholar]
- Buchner, O.; Karadar, M.; Bauer, I.; Neuner, G. A novel system for in situ determination of heat tolerance of plants: First results on alpine dwarf shrubs. Plant Methods 2013, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Buchner, O.; Steiner, P.; Andosch, A.; Holzinger, A.; Stegner, M.; Neuner, G.; Lütz-Meindl, U. A new technical approach for preparing frozen biological samples for electron microscopy. Plant Methods 2020, 16, 48. [Google Scholar] [CrossRef] [Green Version]
- Burke, M.J.; Gusta, L.V.; Quamme, H.A.; Weiser, C.J.; Li, P.H. Freezing and injury in plants. Annu. Rev. Plant Physiol. 1976, 27, 507–528. [Google Scholar] [CrossRef]
- Sakai, A.; Larcher, W. Frost survival of plants. In Responses and Adaptation to Freezing Stress; Springer: Berlin/Heidelberg, Germany; New York, NY, USA; London, UK; Paris, France; Tokyo, Japan, 1987; Volume 62, p. 321. [Google Scholar]
- Ashworth, E.N.; Anderson, J.A.; Davis, G.A.; Lightner, G.W. Ice formation in Prunus persica under field conditions. J. Amer. Soc. Hortic. Sci. 1985, 110, 322–324. [Google Scholar] [CrossRef]
- Neuner, G.; Bannister, P.; Larcher, W. Ice formation and foliar frost resistance in attached and excised shoots from seedlings and adult trees of Nothofagus menziesii. N. Z. J. Bot. 1997, 35, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Kuprian, E.; Munkler, C.; Resnyak, A.; Zimmermann, S.; Tuong, T.D.; Gierlinger, N.; Muller, T.; Livingston, D.P.; Neuner, G. Complex bud architecture and cell-specific chemical patterns enable supercooling of Picea abies bud primordia. Plant Cell Environ. 2017, 40, 3101–3112. [Google Scholar] [CrossRef] [Green Version]
- Meindl, U.; Lancelle, S.; Hepler, P.K. Vesicle production and fusion during lobe formation in Micrasterias visualized by high-pressure freeze fixation. Protoplasma 1992, 170, 104–114. [Google Scholar] [CrossRef]
- Holzinger, A.; Roleda, M.Y.; Lütz, C. The vegetative arctic freshwater green alga Zygnema is insensitive to experimental UV exposure. Micron 2009, 40, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Kuprian, E.; Tuong, T.D.; Pfaller, K.; Wagner, J.; Livingston, D.P.; Neuner, G. Persistent supercooling of reproductive shoots is enabled by structural ice barriers being active despite an intact xylem connection. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Stegner, M.; Buchner, O.; Geßlbauer, M.; Lindner, J.; Floerl, A.; Xiao, N.; Holzinger, A.; Gierlinger, N.; Neuner, G. Frozen mountain pine needles: Endodermal confinement of ice and prevented freeze-dehydration of lignified mesophyll cells allow gas exchange. 2022; submitted manuscript. [Google Scholar]
- Beck, E.; Schulze, E.-D.; Senser, M.; Scheibe, R. Equilibrium freezing of leaf water and extracellular ice formation in Afroalpine ‘giant rosette’ plants. Planta 1984, 162, 276–282. [Google Scholar] [CrossRef]
- Hansen, J.; Beck, E. Evidence for ideal and non-ideal equilibrium freezing of leaf water in frosthardy Ivy (Hedera helix) and Winter Barley (Hordeum vulgare). Bot. Acta 1988, 101, 76–82. [Google Scholar] [CrossRef]
- Palta, J.P.; Levitt, J.; Stadelmann, E.J. Freezing injury in onion bulb cells: I. Evaluation of the conductivity method and analysis of ion and sugar efflux from injured cells. Plant Physiol. 1977, 60, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Buchner, O.; Neuner, G. Freezing cytorrhysis and critical temperature thresholds for photosystem II in the peat moss Sphagnum capillifolium. Protoplasma 2010, 243, 63–71. [Google Scholar] [CrossRef]
- Stegner, M.; Lackner, B.; Schäfernolte, T.; Buchner, O.; Xiao, N.; Gierlinger, N.; Holzinger, A.; Neuner, G. Winter nights during summer time: Stress physiological response to ice and the facilitation of freezing cytorrhysis by elastic cell wall components in leaves of a nival species. Int. J. Mol. Sci. 2020, 21, 7042. [Google Scholar] [CrossRef]
- Anderson, J.; Gusta, L.; Buchanan, D.; Burke, M. Freezing of water in Citrus leaves. J. Amer. Soc. Hortic. Sci. 1983, 108, 397–400. [Google Scholar] [CrossRef]
- Zhu, J.-J.; Beck, E. Water relations of Pachysandra leaves during freezing and thawing: Evidence for a negative pressure potential alleviating freeze-dehydration stress. Plant Physiol. 1991, 97, 1146–1153. [Google Scholar] [CrossRef] [Green Version]
- Stegner, M.; Floerl, A.; Lindner, J.; Plangger, S.; Schäfernolte, T.; Strasser, A.; Thoma, V.; Walde, J.; Neuner, G. Freeze dehydration vs. supercooling of mesophyll cells: Impact of cell wall, cellular and tissue traits on the extent of water displacement. Physiol. Plant. 2022; accepted manuscript. [Google Scholar] [CrossRef]
- Ashworth, E.N.; Pearce, R.S. Extracellular freezing in leaves of freezing-sensitive species. Planta 2002, 214, 798–805. [Google Scholar] [CrossRef]
- Larcher, W.; Meindl, U.; Ralser, E.; Ishikawa, M. Persistent supercooling and silica deposition in cell walls of palm leaves. J. Plant Physiol. 1991, 139, 146–154. [Google Scholar] [CrossRef]
- Ishikawa, M.; Oda, A.; Fukami, R.; Kuriyama, A. Factors contributing to deep supercooling capability and cold survival in dwarf bamboo (Sasa senanensis) leaf blades. Front. Plant Sci. 2014, 5, 791. [Google Scholar] [CrossRef] [Green Version]
- Palta, J.P.; Li, P.H. Frost-hardiness in relation to leaf anatomy and natural distribution of several Solanum species. Crop Sci. 1979, 19, 665–671. [Google Scholar] [CrossRef]
- Hirano, E. Relative abundance of stomata in Citrus and some related genera. Bot. Gaz. 1931, 92, 296–310. [Google Scholar] [CrossRef]
- Huner, N.P.A.; Palta, J.P.; Li, P.H.; Carter, J.V. Anatomical changes in leaves of puma rye in response to growth at cold-hardening temperatures. Bot. Gaz. 1981, 142, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Cox, W.; Levitt, J. An improved, leaf-disk method for determining the freeze-killing temperature of leaves. Cryobiology 1972, 9, 251–256. [Google Scholar] [CrossRef]
- Tanaka, H.; Sato, M.; Ogasawara, Y.; Hamashima, N.; Buchner, O.; Holzinger, A.; Toyooka, K.; Kodama, Y. Chloroplast aggregation during the cold-positioning response in the liverwort Marchantia polymorpha. J. Plant Res. 2017, 130, 1061–1070. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stegner, M.; Buchner, O.; Schäfernolte, T.; Holzinger, A.; Neuner, G. Responses to Ice Formation and Reasons of Frost Injury in Potato Leaves. Crops 2022, 2, 378-389. https://doi.org/10.3390/crops2040026
Stegner M, Buchner O, Schäfernolte T, Holzinger A, Neuner G. Responses to Ice Formation and Reasons of Frost Injury in Potato Leaves. Crops. 2022; 2(4):378-389. https://doi.org/10.3390/crops2040026
Chicago/Turabian StyleStegner, Matthias, Othmar Buchner, Tanja Schäfernolte, Andreas Holzinger, and Gilbert Neuner. 2022. "Responses to Ice Formation and Reasons of Frost Injury in Potato Leaves" Crops 2, no. 4: 378-389. https://doi.org/10.3390/crops2040026