
Mitigation of Emergent Bacterial Pathogens Using Pseudomonas syringae pv. actinidiae as a Case Study—From Orchard to Gene and Everything in Between
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Integrated Approaches to Orchard Management
2.1. Pathogen Detection and Identification
2.2. Understanding Pathogen Epidemiology
2.3. Improvement of Orchard Management
2.4. Anticipating Pathogen Dispersal
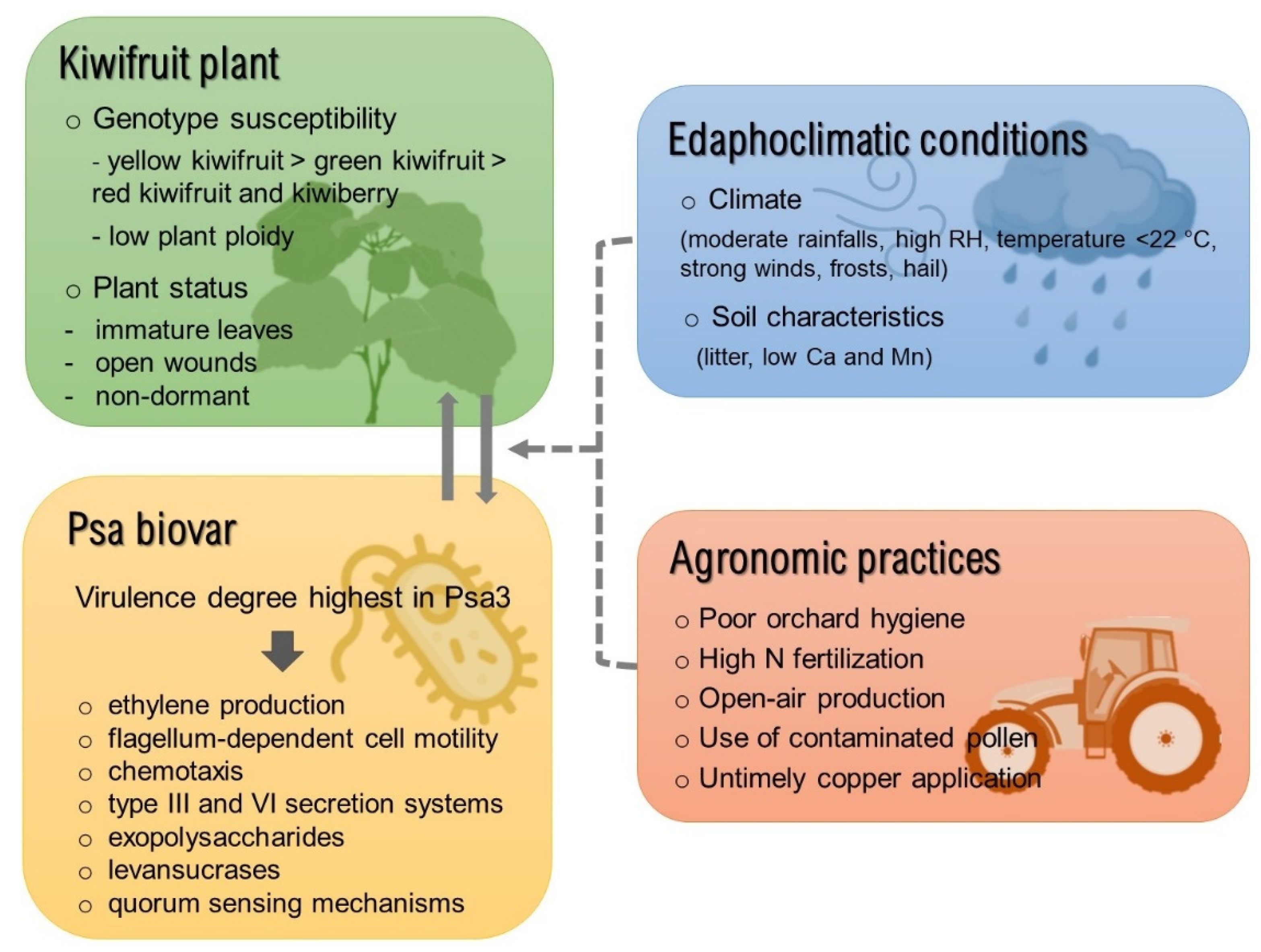
3. Biotic and Abiotic Factors Implicated in Disease Development
3.1. Plant Genotype
3.2. Bacterial Strain
3.3. Edaphoclimatic Conditions
3.4. Agronomical Practices
4. Exploring Plant Genetic Resources for Increased Tolerance
4.1. Pathogen Recognition and Triggered Immunity
4.2. Plant Antioxidant Response
4.3. Phytohormone Regulation
4.4. Primary and Secondary Metabolism
5. Future Prospects
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunjan, M.S.; Lore, J.S. Climate Change: Impact on Plant Pathogens, Diseases, and Their Management. In Crop Protection under Changing Climate; Jabran, K., Florentine, S., Chauhan, B.S., Eds.; Springer International Publishing: Cham, The Netherlands, 2020; pp. 85–100. [Google Scholar]
- European and Mediterranean Plant Protection Organization. EPPO A1 and A2 Lists of Pests Recommended for Regulation as Quarantine Pests, Version 2021-09; Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/A2_list (accessed on 1 August 2022).
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef]
- Bull, C.T.; De Boer, S.; Denny, T.; Firrao, G.; Saux, M.F.-L.; Saddler, G.; Scortichini, M.; Stead, D.; Takikawa, Y. Comprehensive list of names of plant pathogenic bacteria, 1980–2007. J. Plant Pathol. 2010, 551–592. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. Pseudomonas syringae pv. actinidiae—Distribution; Available online: https://gd.eppo.int/taxon/PSDMAK/distribution (accessed on 1 August 2022).
- Serizawa, S.; Ichikawa, T.; Takikawa, Y.; Tsuyumu, S.; Goto, M. Occurrence of bacterial canker of kiwifruit in Japan description of symptoms, isolation of the pathogen and screening of bactericides. Jpn. J. Phytopathol. 1989, 55, 427–436. [Google Scholar] [CrossRef]
- Ferrante, P.; Scortichini, M. Identification of Pseudomonas syringae pv. actinidiae as causal agent of bacterial canker of yellow kiwifruit (Actinidia chinensis Planchon) in Central Italy. J. Phytopathol. 2009, 157, 768–770. [Google Scholar] [CrossRef]
- Scortichini, M.; Marcelletti, S.; Ferrante, P.; Petriccione, M.; Firrao, G. Pseudomonas syringae pv. actinidiae: A re-emerging, multi-faceted, pandemic pathogen. Mol. Plant Pathol. 2012, 13, 631–640. [Google Scholar] [CrossRef]
- Vanneste, J.L. The scientific, economic, and social impacts of the New Zealand outbreak of bacterial canker of kiwifruit (Pseudomonas syringae pv. actinidiae). Annu. Rev. Phytopathol. 2017, 55, 377–399. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. Guidelines on Pest Risk Analysis; EPPO: Paris, France, 1993; pp. 191–198. [Google Scholar]
- Abelleira, A.; Ares, A.; Aguín, O.; Mansilla, P.; López, M.C. Distribution, detection protocols and characterization of Pseudomonas syringae pv. actinidiae from kiwifruit in Galicia (northwest Spain). Acta Hortic. 2015, 1095, 49–56. [Google Scholar] [CrossRef]
- Donati, I.; Cellini, A.; Sangiorgio, D.; Vanneste, J.L.; Scortichini, M.; Balestra, G.M.; Spinelli, F. Pseudomonas syringae pv. actinidiae: Ecology, infection dynamics and disease epidemiology. Microb. Ecol. 2020, 80, 81–102. [Google Scholar] [CrossRef]
- Al-Saffar, M.F.; Jarallah, E.M. Isolation and characterization of Pseudomonas aeruginosa from Babylon Province. Biochem. Cell. Arch. 2019, 19, 203–209. [Google Scholar]
- Cornelis, P.; Matthijs, S. Pseudomonas Siderophores and Their Biological Significance. In Microbial Siderophores; Varma, A., Chincholkar, S.B., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 193–203. [Google Scholar]
- Kałużna, M. Characterization and phylogeny of the novel taxon of Pseudomonas spp., closely related to Pseudomonas avellanae as causal agent of a bacterial leaf blight of cornelian cherry (Cornus mas L.) and Pseudomonas syringae pv. syringae as a new bacterial pathogen of red dogwood (Cornus sanguinea L.). J. Plant Pathol. 2019, 101, 251–261. [Google Scholar] [CrossRef]
- Nepali, B.; Bhattarai, S.; Shrestha, J. Identification of Pseudomonas fluorescens using different biochemical tests. Int. J. Appl. Biol. 2018, 2, 27–32. [Google Scholar] [CrossRef]
- González, A.J.; Rodicio, M.R.; Mendoza, M.C. Identification of an emergent and atypical Pseudomonas viridiflava lineage causing bacteriosis in plants of agronomic importance in a Spanish region. Appl. Environ. Microbiol. 2003, 69, 2936–2941. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. PM 7/120 (1) Pseudomonas syringae pv. actinidiae; pp. 360–375. Available online: https://onlinelibrary.wiley.com/doi/pdf/10.1111/epp.12171 (accessed on 1 August 2022).
- Prencipe, S.; Gullino, M.L.; Spadaro, D. Pseudomonas syringae pv. actinidiae isolated from Actinidia chinensis var. deliciosa in Northern Italy: Genetic diversity and virulence. Eur. J. Plant Pathol. 2018, 150, 191–204. [Google Scholar] [CrossRef]
- Brunetti, A.; Pucci, N.; Modesti, V.; Lumia, V.; Latini, A.; Loreti, S.; Pilotti, M. In vitro and in planta screening of compounds for the control of Pseudomonas syringae pv. actinidiae in Actinidia chinensis var. chinensis. Eur. J. Plant Pathol. 2020, 158, 829–848. [Google Scholar] [CrossRef]
- Sawada, H.; Takeuchi, T.; Matsuda, I. Comparative analysis of Pseudomonas syringae pv. actinidiae and pv. phaseolicola based on phaseolotoxin-resistant ornithine carbamoyltransferase gene (argK) and 16S-23S rRNA intergenic spacer sequences. Appl. Environ. Microbiol. 1997, 63, 282–288. [Google Scholar] [CrossRef]
- Koh, Y.J.; Nou, I.S. DNA markers for identification of Pseudomonas syringae pv. actinidiae. Mol. Cells 2002, 13, 309–314. [Google Scholar]
- Rees-George, J.; Vanneste, J.; Cornish, D.; Pushparajah, I.; Yu, J.; Templeton, M.; Everett, K. Detection of Pseudomonas syringae pv. actinidiae using polymerase chain reaction (PCR) primers based on the 16S–23S rDNA intertranscribed spacer region and comparison with PCR primers based on other gene regions. Plant Pathol. 2010, 59, 453–464. [Google Scholar] [CrossRef]
- Gallelli, A.; L’Aurora, A.; Loreti, S. Gene sequence analysis for the molecular detection of Pseudomonas syringae pv. actinidiae: Developing diagnostic protocols. J. Plant Pathol. 2011, 93, 425–435. [Google Scholar] [CrossRef]
- Gallelli, A.; Talocci, S.; Pilotti, M.; Loreti, S. Real-time and qualitative PCR for detecting Pseudomonas syringae pv. actinidiae isolates causing recent outbreaks of kiwifruit bacterial canker. Plant Pathol. 2014, 63, 264–276. [Google Scholar] [CrossRef]
- Balestra, G.M.; Taratufolo, M.C.; Vinatzer, B.A.; Mazzaglia, A. A multiplex PCR assay for detection of Pseudomonas syringae pv. actinidiae and differentiation of populations with different geographic origin. Plant Dis. 2013, 97, 472–478. [Google Scholar] [CrossRef]
- Ciarroni, S.; Gallipoli, L.; Taratufolo, M.C.; Butler, M.I.; Poulter, R.T.; Pourcel, C.; Vergnaud, G.; Balestra, G.M.; Mazzaglia, A. Development of a multiple loci variable number of tandem repeats analysis (MLVA) to unravel the intra-pathovar structure of Pseudomonas syringae pv. actinidiae populations worldwide. PLoS ONE 2015, 10, e0135310. [Google Scholar] [CrossRef] [PubMed]
- Cunty, A.; Cesbron, S.; Poliakoff, F.; Jacques, M.A.; Manceau, C. Origin of the outbreak in France of Pseudomonas syringae pv. actinidiae biovar 3, the causal agent of bacterial canker of kiwifruit, revealed by a multilocus variable-number tandem-repeat analysis. Appl. Environ. Microbiol. 2015, 81, 6773–6789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzaglia, A.; Turco, S.; Taratufolo, M.C.; Tatì, M.; Rahi, Y.J.; Gallipoli, L.; Balestra, G.M. Improved MLVA typing reveals a highly articulated structure in Pseudomonas syringae pv. actinidiae populations. Physiol. Mol. Plant Pathol. 2021, 114, 101636. [Google Scholar] [CrossRef]
- Sawada, H.; Fujikawa, T. Genetic diversity of Pseudomonas syringae pv. actinidiae, pathogen of kiwifruit bacterial canker. Plant Pathol. 2019, 68, 1235–1248. [Google Scholar] [CrossRef]
- Pan, X.; Zhao, S.; Wang, Y.; Li, M.; He, L.; Zhuang, Q. Complete genome sequencing of Pseudomonas syringae pv. actinidiae Biovar 3, P155, kiwifruit pathogen originating from China. Biosci. J. 2020, 36, 2220–2228. [Google Scholar] [CrossRef]
- Andersen, M.T.; Templeton, M.D.; Rees-George, J.; Vanneste, J.L.; Cornish, D.A.; Yu, J.; Cui, W.; Braggins, T.J.; Babu, K.; Mackay, J.F.; et al. Highly specific assays to detect isolates of Pseudomonas syringae pv. actinidiae biovar 3 and Pseudomonas syringae pv. actinidifoliorum directly from plant material. Plant Pathol. 2018, 67, 1220–1230. [Google Scholar] [CrossRef]
- Barrett-Manako, K.; Andersen, M.; Martínez-Sánchez, M.; Jenkins, H.; Hunter, S.; Reese-George, J.; Montefiori, M.; Wohlers, M.; Rikkerink, E.; Templeton, M.; et al. Real-time PCR and droplet digital PCR are accurate and reliable methods to quantify Pseudomonas syringae pv. actinidiae biovar 3 in kiwifruit infected plantlets. Plant Dis. 2021, 105, 1748–1757. [Google Scholar] [CrossRef]
- Jayaraman, J.; Chatterjee, A.; Hunter, S.; Chen, R.; Stroud, E.A.; Saei, H.; Hoyte, S.; Deroles, S.; Tahir, J.; Templeton, M.D.; et al. Rapid methodologies for assessing Pseudomonas syringae pv. actinidiae colonization and effector-mediated hypersensitive response in kiwifruit. Mol. Plant-Microbe Interact. 2021, 34, 880–890. [Google Scholar] [CrossRef]
- Ruinelli, M.; Schneeberger, P.H.H.; Ferrante, P.; Bühlmann, A.; Scortichini, M.; Vanneste, J.L.; Duffy, B.; Pothier, J.F. Comparative genomics-informed design of two LAMP assays for detection of the kiwifruit pathogen Pseudomonas syringae pv. actinidiae and discrimination of isolates belonging to the pandemic biovar 3. Plant Pathol. 2017, 66, 140–149. [Google Scholar] [CrossRef]
- Suzaki, K.; Sawada, H.; Kisaki, G. Loop-mediated isothermal amplification of bacterial effector genes to detect Pseudomonas syringae pv. actinidiae biovars 1 and 3. J. Gen. Plant Pathol. 2022, 88, 2–9. [Google Scholar] [CrossRef]
- Liu, P.; Xue, S.; He, R.; Hu, J.; Wang, X.; Jia, B.; Gallipoli, L.; Mazzaglia, A.; Balestra, G.M.; Zhu, L. Pseudomonas syringae pv. actinidiae isolated from non-kiwifruit plant species in China. Eur. J. Plant Pathol. 2016, 145, 743–754. [Google Scholar] [CrossRef]
- Li, L.; Pan, H.; Deng, L.; Feng, D.D.; Zhong, C.H. First report of bacterial leaf spot disease of Broussonetia papyrifera caused by Pseudomonas syringae pv. actinidiae in China. Plant Dis. 2021, 105, 696. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, F.; Donati, I.; Vanneste, J.; Costa, M.; Costa, G. Real time monitoring of the interactions between Pseudomonas syringae pv. actinidiae and Actinidia species. Acta Hortic. 2010, 913, 461–465. [Google Scholar] [CrossRef]
- Ferrante, P.; Fiorillo, E.; Marcelletti, S.; Marocchi, F.; Mastroleo, M.; Simeoni, S.; Scortichini, M. The importance of the main colonization and penetration sites of Pseudomonas syringae pv. actinidiae and prevailing weather conditions in the development of epidemics in yellow kiwifruit, recently observed in Central Italy. J. Plant Pathol. 2012, 94, 455–461. [Google Scholar] [CrossRef]
- Serizawa, S.; Ichikawa, T. Epidemiology of bacterial canker of kiwifruit 1. Infection and bacterial movement in tissue of new canes. Jpn. J. Phytopathol. 1993, 59, 452–459. [Google Scholar] [CrossRef]
- Petriccione, M.; Salzano, A.M.; Di Cecco, I.; Scaloni, A.; Scortichini, M. Proteomic analysis of the Actinidia deliciosa leaf apoplast during biotrophic colonization by Pseudomonas syringae pv. actinidiae. J. Proteom. 2014, 101, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Renzi, M.; Copini, P.; Taddei, A.R.; Rossetti, A.; Gallipoli, L.; Mazzaglia, A.; Balestra, G.M. Bacterial canker on kiwifruit in Italy: Anatomical changes in the wood and in the primary infection sites. Phytopathology 2012, 102, 827–840. [Google Scholar] [CrossRef]
- Vanneste, J.L.; Cornish, D.A.; Yu, J.; Stokes, C.A. First report of Pseudomonas syringae pv. actinidiae the causal agent of bacterial canker of kiwifruit on Actinidia arguta vines in New Zealand. Plant Dis. 2014, 98, 418. [Google Scholar] [CrossRef]
- Beatson, R.A.; Datson, P.M.; Ferguson, A.R.; Montefiori, M. Use of kiwifruit germplasm resources for genetic improvement. Acta Hortic. 2014, 1048, 25–34. [Google Scholar] [CrossRef]
- Serizawa, S.; Ichikawa, T. Epidemiology of bacterial canker of kiwifruit, 3: The seasonal changes of bacterial population in lesions and of its exudation from lesion. Jpn. J. Phytopathol. 1993, 59, 469–476. [Google Scholar] [CrossRef]
- Serizawa, S.; Ichikawa, T. Epidemiology of bacterial canker of kiwifruit. 2. The most suitable times and environments for infection on new canes. Jpn. J. Phytopathol. 1993, 59, 460–468. [Google Scholar] [CrossRef]
- Serizawa, S.; Ichikawa, T.; Suzuki, H. Epidemiology of bacterial canker of kiwifruit. 5. Effect of infection in fall to early winter on the disease development in branches and trunk after winter. Jpn. J. Phytopathol. 1994, 60, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Serizawa, S.; Ichikawa, T. Epidemiology of bacterial canker of kiwifruit. 4. Optimum temperature for disease development on new canes. Jpn. J. Phytopathol. 1993, 59, 694–701. [Google Scholar] [CrossRef]
- Mauri, S.; Cellini, A.; Buriani, G.; Donati, I.; Costa, G.; Spinelli, F. Optimization of cultural practices to reduce the development of Pseudomonas syringae pv. actinidiae, causal agent of the bacterial canker of kiwifruit. J. Berry Res. 2016, 6, 355–371. [Google Scholar] [CrossRef]
- Aguín, O.; Ares, A.; Abelleira, A.; Mansilla, P. Survival of Pseudomonas syringae pv. actinidiae in leaf-litter of Actinidia deliciosa in Galicia (northwest Spain). Acta Hortic. 2015, 1095, 111–115. [Google Scholar] [CrossRef]
- Tyson, J.; Curtis, C.; Manning, M.; Dobson, S.; McKenna, C. Preliminary investigations of the risk of plant debris as a Pseudomonas syringae pv actinidiae inoculum source. N. Z. Plant Prot. 2016, 69, 11–16. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. Pest Risk Analysis for Pseudomonas syringae pv. actinidiae; Available online: https://gd.eppo.int/download/doc/1255_pra_exp_PSDMAK.pdf (accessed on 1 August 2022).
- Koh, Y.J.; Park, S.Y.; Lee, D.H. Characteristics of bacterial canker of kiwifruit occurring in Korea and its control by trunk injection. Plant Pathol. J. 1996, 12, 324–330. [Google Scholar]
- Kennelly, M.M.; Cazorla, F.M.; de Vicente, A.; Ramos, C.; Sundin, G.W. Pseudomonas syringae diseases of fruit trees: Progress toward understanding and control. Plant Dis. 2007, 91, 4–17. [Google Scholar] [CrossRef]
- Koh, Y.J.; Kim, G.H.; Jung, J.S.; Lee, Y.S.; Hur, J.S. Outbreak of bacterial canker on Hort16A (Actinidia chinensis Planchon) caused by Pseudomonas syringae pv. actinidiae in Korea. N. Z. J. Crop Hortic. Sci. 2010, 38, 275–282. [Google Scholar] [CrossRef]
- Direção-Geral de Alimentação e Veterinária, D. Plano de ação Nacional para o Controlo da Pseudomonas syringae pv. actinidiae do Kiwi (Psa); Available online: https://www.dgav.pt/wp-content/uploads/2021/01/Plano-de-Acao-Nacional-de-Controlo-da-Pseudomonas-syringae-pv-actinidiae-do-kiwi-2014.pdf (accessed on 1 August 2022).
- Cornish, D.A.; Yu, J.; Oldham, J.M.; Benge, J.R.; Max, W.; Vanneste, J.L. In vitro inhibition of Pseudomonas syringae pv. actinidiae by wound protectants. N. Z. Plant Prot. 2015, 68, 332–339. [Google Scholar] [CrossRef]
- Everett, K.R.; Pushparajah, I.P.S.; Vergara, M.; Shahjahan, K.; Parry, B. S C. Monitoring Effectiveness of Wound Protectants against Psa. Milestone N° 66873. A Report Prepared for ZESPRI Group Limited; Plant & Food Research: Auckland, New Zealand, 2016. [Google Scholar]
- Everett, K.R.; Shahjahan, K.; Pushparajah, I.P.S.; Ramos, L.; Parry, B.; Hasna, L.; Middleditch, C.; Vergara, M.J. Monitoring Effectiveness of Wound Protectants against Psa. Milestone N° 68497: VI1712-30-G. A Report Prepared for ZESPRI Group Limited; Plant & Food Research: Auckland, New Zealand, 2017. [Google Scholar]
- Woodcock, S.D. A Review of Research and Development Undertaken on Psa. A Report Prepared for ZESPRI Group Limited; Kiwifruit Vine Health (KVH), Mt.: Maunganui, New Zealand, 2016. [Google Scholar]
- Cameron, A.; Sarojini, V. Pseudomonas syringae pv. actinidiae: Chemical control, resistance mechanisms and possible alternatives. Plant Pathol. 2014, 63, 1–11. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, G.H.; Song, Y.-R.; Oh, C.-S.; Koh, Y.J.; Jung, J.S. Streptomycin resistant isolates of Pseudomonas syringae pv. actinidiae in Korea. Res. Plant Dis. 2020, 26, 44–47. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.-N. Thirteen decades of antimicrobial copper compounds applied in agriculture. A review. Agron. Sustain. Dev. 2018, 38, 28. [Google Scholar] [CrossRef] [Green Version]
- Sciubba, F.; Di Cocco, M.E.; Angori, G.; Spagnoli, M.; De Salvador, F.R.; Engel, P.; Delfini, M. NMR-based metabolic study of leaves of three species of Actinidia with different degrees of susceptibility to Pseudomonas syringae pv. actinidiae. Nat. Prod. Res. 2019, 34, 2043–2050. [Google Scholar] [CrossRef] [PubMed]
- Ashrafzadeh, S.; Leung, D.W.M. In vitro breeding-shortcut to Pseudomonas syringae pv. actinidiae (Psa) tolerant kiwifruit. Arch. Phytopathol. Plant Prot. 2019, 52, 501–506. [Google Scholar] [CrossRef]
- Scortichini, M. Field efficacy of a zinc-copper-hydracid of citric acid biocomplex compound to reduce oozing from winter cankers caused by Pseudomonas syringae pv. actinidiae to Actinidia spp. J. Plant Pathol. 2016, 98, 651–655. [Google Scholar]
- Monchiero, M.; Gullino, M.L.; Pugliese, M.; Spadaro, D.; Garibaldi, A. Efficacy of different chemical and biological products in the control of Pseudomonas syringae pv. actinidiae on kiwifruit. Australas. Plant Pathol. 2015, 44, 13–23. [Google Scholar] [CrossRef]
- Kim, G.H.; Lee, Y.S.; Jung, J.S.; Koh, Y.J.; Poulter, R.T.M.; Butler, M. Genomic analyses of Pseudomonas syringae pv. actinidiae isolated in Korea suggest the transfer of the bacterial pathogen via kiwifruit pollen. J. Med. Microbiol. 2020, 69, 132–138. [Google Scholar] [CrossRef]
- Narouei-Khandan, H.A.; Worner, S.P.; Viljanen, S.L.; van Bruggen, A.H.; Balestra, G.M.; Jones, E. The potential global climate suitability of kiwifruit bacterial canker disease (Pseudomonas syringae pv. actinidiae (Psa)) using three modelling approaches: CLIMEX, Maxent and Multimodel Framework. Climate 2022, 10, 14. [Google Scholar] [CrossRef]
- Wang, R.; Li, Q.; He, S.; Liu, Y.; Wang, M.; Jiang, G. Modeling and mapping the current and future distribution of Pseudomonas syringae pv. actinidiae under climate change in China. PLoS ONE 2018, 13, e0192153. [Google Scholar] [CrossRef]
- Nardozza, S.; Boldingh, H.; Richardson, A.; Walter, M.; Kashuba, P.; Seelye, R.; Clearwater, M.; Gould, N. Kiwifruit xylem sap: Composition and in vitro growth of a virulent strain of Pseudomonas syringae pv. actinidiae. Acta Hortic. 2015, 1095, 123–128. [Google Scholar] [CrossRef]
- Kisaki, G.; Tanaka, S.; Ishihara, A.; Igarashi, C.; Morimoto, T.; Hamano, K.; Endo, A.; Sugita-Konishi, S.; Tabuchi, M.; Gomi, K.; et al. Evaluation of various cultivars of Actinidia species and breeding source Actinidia rufa for resistance to Pseudomonas syringae pv. actinidiae biovar 3. J. Gen. Plant Pathol. 2018, 84, 399–406. [Google Scholar] [CrossRef]
- Saei, A.; Hoeata, K.; Krebs, A.; Sutton, P.; Herrick, J.; Wood, M.; Gea, L. The status of Pseudomonas syringae pv. actinidiae (Psa) in the New Zealand kiwifruit breeding programme in relation to ploidy level. Acta Hortic. 2018, 1218, 293–298. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Vasconcelos, M.W.; Gaspar, M.; Balestra, G.M.; Mazzaglia, A.; Carvalho, S.M.P. Early pathogen recognition and antioxidant system activation contributes to Actinidia arguta tolerance against Pseudomonas syringae pathovars actinidiae and actinidifoliorum. Front. Plant Sci. 2020, 11, 1022. [Google Scholar] [CrossRef]
- Wang, F.M.; Mo, Q.H.; Ye, K.Y.; Gong, H.J.; Qi, B.B.; Liu, P.P.; Jiang, Q.S.; Li, J.W. Evaluation of the wild Actinidia germplasm for resistance to Pseudomonas syringae pv. actinidiae. Plant Pathol. 2020, 69, 979–989. [Google Scholar] [CrossRef]
- Thomidis, T.; Goumas, D.E.; Zotos, A.; Triantafyllidis, V.; Kokotos, E. Susceptibility of twenty-three kiwifruit cultivars to Pseudomonas syringae pv. actinidiae. Eng. Proc. 2021, 9, 33. [Google Scholar] [CrossRef]
- Cotruţ, R.; Renzi, M.; Taratufolo, M.C.; Mazzaglia, A.; Balestra, G.M.; Stănică, F. Actinidia arguta ploidy level variation in relation to Pseudomonas syringae pv. actinidiae susceptibility. Lucr. Ştiinţifice 2013, 56, 1–12. [Google Scholar]
- Wang, F.-M.; Li, J.-W.; Ye, K.-Y.; Gong, H.-J.; Liu, P.-P.; Jiang, Q.-S.; Qi, B.-B.; Mo, Q.-H. Preliminary report on the improved resistance towards Pseudomonas syringae pv. actinidiae of cultivated kiwifruit (Actinidia chinensis) when grafted onto wild Actinidia guilinensis rootstock in vitro. J. Plant Pathol. 2021, 103, 51–54. [Google Scholar] [CrossRef]
- Chapman, J.R.; Taylor, R.K.; Weir, B.S.; Romberg, M.K.; Vanneste, J.L.; Luck, J.; Alexander, B.J. Phylogenetic relationships among global populations of Pseudomonas syringae pv. actinidiae. Phytopathology 2012, 102, 1034–1044. [Google Scholar] [CrossRef]
- Cunty, A.; Poliakoff, F.; Rivoal, C.; Cesbron, S.; Fischer-Le Saux, M.; Lemaire, C.; Jacques, M.A.; Manceau, C.; Vanneste, J.L. Characterization of Pseudomonas syringae pv. actinidiae (Psa) isolated from France and assignment of Psa biovar 4 to a de novo pathovar: Pseudomonas syringae pv. actinidifoliorum pv. nov. Plant Pathol. 2015, 64, 582–596. [Google Scholar] [CrossRef]
- Fujikawa, T.; Sawada, H. Genome analysis of the kiwifruit canker pathogen Pseudomonas syringae pv. actinidiae biovar 5. Sci. Rep. 2016, 6, 21399. [Google Scholar] [CrossRef] [PubMed]
- Sawada, H.; Kondo, K.; Nakaune, R. Novel biovar (biovar 6) of Pseudomonas syringae pv. actinidiae causing bacterial canker of kiwifruit (Actinidia deliciosa) in Japan. Jpn. J. Phytopathol. 2016, 82, 101–115. [Google Scholar] [CrossRef]
- Sawada, H.; Miyoshi, T.; Ide, Y. Novel MLSA group (Psa5) of Pseudomonas syringae pv. actinidiae causing bacterial canker of kiwifruit (Actinidia chinensis) in Japan. Jpn. J. Phytopathol. 2014, 80, 171–184. [Google Scholar] [CrossRef]
- McCann, H.C.; Li, L.; Liu, Y.; Li, D.; Pan, H.; Zhong, C.; Rikkerink, E.H.A.; Templeton, M.D.; Straub, C.; Colombi, E.; et al. Origin and evolution of the kiwifruit canker pandemic. Genome Biol. Evol. 2017, 9, 932–944. [Google Scholar] [CrossRef]
- Fujikawa, T.; Hatomi, H.; Sawada, H. Draft genome sequences of 10 strains of Pseudomonas syringae pv. actinidiae biovar 1, a major kiwifruit bacterial canker pathogen in Japan. Microbiol. Resour. Announc. 2020, 9, e00759-20. [Google Scholar] [CrossRef]
- Genka, H.; Baba, T.; Tsuda, M.; Kanaya, S.; Mori, H.; Yoshida, T.; Noguchi, M.T.; Tsuchiya, K.; Sawada, H. Comparative analysis of argK-tox clusters and their flanking regions in phaseolotoxin-producing Pseudomonas syringae pathovars. J. Mol. Evol. 2006, 63, 401–414. [Google Scholar] [CrossRef]
- Gnanamanickam, S.S. Plant-Associated Bacteria, 1st ed.; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]
- Cellini, A.; Donati, I.; Farneti, B.; Khomenko, I.; Buriani, G.; Biasioli, F.; Cristescu, S.M.; Spinelli, F. A breach in plant defences: Pseudomonas syringae pv. actinidiae targets ethylene signalling to overcome Actinidia chinensis pathogen responses. Int. J. Mol. Sci. 2021, 22, 4375. [Google Scholar] [CrossRef]
- Donati, I.; Buriani, G.; Cellini, A.; Mauri, S.; Costa, G.; Spinelli, F. New insights on the bacterial canker of kiwifruit (Pseudomonas syringae pv. actinidiae). J. Berry Res. 2014, 4, 53–67. [Google Scholar] [CrossRef]
- Vandelle, E.; Colombo, T.; Regaiolo, A.; Maurizio, V.; Libardi, T.; Puttilli, M.-R.; Danzi, D.; Polverari, A. Transcriptional profiling of three Pseudomonas syringae pv. actinidiae biovars reveals different responses to apoplast-like conditions related to strain virulence on the host. Mol. Plant-Microbe Interact. 2021, 34, 376–396. [Google Scholar] [CrossRef]
- Bundalovic-Torma, C.; Lonjon, F.; Desveaux, D.; Guttman, D.S. Diversity, evolution, and function of Pseudomonas syringae effectoromes. Annu. Rev. Phytopathol. 2022, 60, 211–236. [Google Scholar] [CrossRef]
- Ishiga, T.; Sakata, N.; Nguyen, V.T.; Ishiga, Y. Flood inoculation of seedlings on culture medium to study interactions between Pseudomonas syringae pv. actinidiae and kiwifruit. J. Gen. Plant Pathol. 2020, 86, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, J.; Yoon, M.; Applegate, E.R.; Stroud, E.A.; Templeton, M.D. AvrE1 and HopR1 from Pseudomonas syringae pv. actinidiae are additively required for full virulence on kiwifruit. Mol. Plant Pathol. 2020, 21, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, Q.; Zhi, T.; Fan, R.; Xie, T.; Zhao, Z.; Long, Y.; Li, Z. Genetic causes of non-pathogenic Pseudomonas syringae pv. actinidiae isolates in kiwifruit orchards. Front. Microbiol. 2021, 12, 650099. [Google Scholar] [CrossRef] [PubMed]
- Hemara, L.M.; Jayaraman, J.; Sutherland, P.W.; Montefiori, M.; Arshed, S.; Chatterjee, A.; Chen, R.; Andersen, M.; Mesarich, C.H.; van der Linden, O.; et al. Effector loss drives adaptation of Pseudomonas syringae pv. actinidiae to Actinidia arguta. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wang, N.; Han, N.; Tian, R.; Chen, J.; Gao, X.; Wu, Z.; Liu, Y.; Huang, L. Role of the type VI secretion system in the pathogenicity of Pseudomonas syringae pv. actinidiae, the causative agent of kiwifruit bacterial canker. Front. Microbiol. 2021, 12, 627785. [Google Scholar] [CrossRef] [PubMed]
- Firrao, G.; Torelli, E.; Polano, C.; Ferrante, P.; Ferrini, F.; Martini, M.; Marcelletti, S.; Scortichini, M.; Ermacora, P. Genomic structural variations affecting virulence during clonal expansion of Pseudomonas syringae pv. actinidiae biovar 3 in Europe. Front. Microbiol. 2018, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, M.; Liu, W.; Nie, J.; Huang, L. Pseudomonas syringae pv. actinidiae effector HopAU1 interacts with calcium-sensing receptor to activate plant immunity. Int. J. Mol. Sci. 2022, 23, 508. [Google Scholar] [CrossRef]
- Andolfi, A.; Ferrante, P.; Petriccione, M.; Cimmino, A.; Evidente, A.; Scortichini, M. Production of phytotoxic metabolites by Pseudomonas syringae pv. actinidiae, the causal agent of bacterial canker of kiwifruit. J. Plant Pathol. 2014, 96, 169–175. [Google Scholar] [CrossRef]
- McAtee, P.A.; Brian, L.; Curran, B.; van der Linden, O.; Nieuwenhuizen, N.J.; Chen, X.; Henry-Kirk, R.A.; Stroud, E.A.; Nardozza, S.; Jayaraman, J.; et al. Re-programming of Pseudomonas syringae pv. actinidiae gene expression during early stages of infection of kiwifruit. BMC Genom. 2018, 19, 822. [Google Scholar] [CrossRef]
- Luti, S.; Campigli, S.; Ranaldi, F.; Paoli, P.; Pazzagli, L.; Marchi, G. Lscβ and lscγ, two novel levansucrases of Pseudomonas syringae pv. actinidiae biovar 3, the causal agent of bacterial canker of kiwifruit, show different enzymatic properties. Int. J. Biol. Macromol. 2021, 179, 279–291. [Google Scholar] [CrossRef]
- Cellini, A.; Buriani, G.; Correia, C.; Fiorentini, L.; Vandelle, E.; Polverari, A.; Santos, C.; Vanneste, J.L.; Spinelli, F. Host-specific signal perception by PsaR2 LuxR solo induces Pseudomonas syringae pv. actinidiae virulence traits. Microbiol. Res. 2022, 260, 127048. [Google Scholar] [CrossRef] [PubMed]
- Cellini, A.; Donati, I.; Fiorentini, L.; Vandelle, E.; Polverari, A.; Venturi, V.; Buriani, G.; Vanneste, J.L.; Spinelli, F. N-Acyl homoserine lactones and Lux solos regulate social behaviour and virulence of Pseudomonas syringae pv. actinidiae. Microb. Ecol. 2020, 79, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Purahong, W.; Orrù, L.; Donati, I.; Perpetuini, G.; Cellini, A.; Lamontanara, A.; Michelotti, V.; Tacconi, G.; Spinelli, F. Plant microbiome and its link to plant health: Host species, organs and Pseudomonas syringae pv. actinidiae infection shaping bacterial phyllosphere communities of kiwifruit plants. Front. Plant Sci. 2018, 9, 1563. [Google Scholar] [CrossRef] [PubMed]
- Ares, A.; Pereira, J.; Garcia, E.; Costa, J.; Tiago, I. The leaf bacterial microbiota of female and male kiwifruit plants in distinct seasons: Assessing the impact of Pseudomonas syringae pv. actinidiae. Phytobiomes J. 2021, 5, 275–287. [Google Scholar] [CrossRef]
- Ferrante, P.; Scortichini, M. Redefining the global populations of Pseudomonas syringae pv. actinidiae based on pathogenic, molecular and phenotypic characteristics. Plant Pathol. 2015, 64, 51–62. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Machado, J.; Balestra, G.M.; Mazzaglia, A.; Vasconcelos, M.W.; Carvalho, S.M.P. Exploring the expression of defence-related genes in Actinidia spp. after infection with Pseudomonas syringae pv. actinidiae and pv. actinidifoliorum: First steps. Eur. J. Hortic. Sci. 2019, 84, 206–212. [Google Scholar] [CrossRef]
- Vanneste, J.L.; Yu, J.; Cornish, D.A.; Tanner, D.J.; Windner, R.; Chapman, J.R.; Taylor, R.K.; Mackay, J.F.; Dowlut, S. Identification, virulence, and distribution of two biovars of Pseudomonas syringae pv. actinidiae in New Zealand. Plant Dis. 2013, 97, 708–719. [Google Scholar] [CrossRef]
- Ochoa-Díaz, M.M.; Daza-Giovannetty, S.; Gómez-Camargo, D. Bacterial genotyping methods: From the basics to modern. Methods Mol. Biol. 2018, 1734, 13–20. [Google Scholar] [CrossRef]
- Ko, S.-J.; Lee, Y.-H.; Cha, K.-H.; Park, K.-B.; Park, I.-J.; Kim, Y.-C. An improved method for testing pathogenicity of Pseudomonas syringae pv. actinidiae causing bacterial canker of kiwifruit. Res. Plant Dis. 2002, 8, 250–253. [Google Scholar] [CrossRef]
- Hirano, S.S.; Upper, C.D. Population biology and epidemiology of Pseudomonas syringae. Annu. Rev. Phytopathol. 1990, 28, 155–177. [Google Scholar] [CrossRef]
- Vanneste, J.; Reglinski, T.; Yu, J.; Cornish, D. Multiplication and movement of Pseudomonas syringae pv. actinidiae in kiwifruit plants. Acta Hortic. 2015, 1095, 117–122. [Google Scholar] [CrossRef]
- Li, Y.; Cheng, H.; Fang, S.; Qian, Z. Prevalent forecast of kiwifruit bacterial canker caused by Pseudomonas syringae pv. actinidiae. Ying Yong Sheng Tai Xue Bao 2001, 12, 355–358. [Google Scholar] [PubMed]
- Antoniacci, L.; Bugiani, R.; Rossi, R.; Calzolari, A.; Alessandrini, A.; Gozzi, R.; Spinelli, F.; Cellini, A.; Mauri, S.; Donati, I. Validation of New Zealand Psa forecasting model in Emilia Romagna Region, Italy. Acta Hortic. 2019, 1243, 71–78. [Google Scholar] [CrossRef]
- Choi, S.; Jayaraman, J.; Segonzac, C.; Park, H.J.; Park, H.; Han, S.W.; Sohn, K.H. Pseudomonas syringae pv. actinidiae type III effectors localized at multiple cellular compartments activate or suppress innate immune responses in Nicotiana benthamiana. Front. Plant Sci. 2017, 8, 2157. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, P.; Scortichini, M. Frost promotes the pathogenicity of Pseudomonas syringae pv. actinidiae in Actinidia chinensis and A. deliciosa plants. Plant Pathol. 2014, 63, 12–19. [Google Scholar] [CrossRef]
- Kiwifruit Vine Health, K. Psa-V Best Practice Guide; Kiwifruit Vine Health (KVH): Mount Maunganui, New Zealand, 2018. [Google Scholar]
- Chiabrando, V.; Giacalone, G. Kiwifruit under plastic covering: Impact on fruit quality and on orchard microclimate. J. Food Nutr. Agric. 2018, 1, 1–6. [Google Scholar] [CrossRef]
- Black, M.Z.; Casonato, S.; Bent, S.J. Opportunities for environmental modification to control Pseudomonas syringae pv. actinidiae in kiwifruit. Acta Hortic. 2015, 1105, 253–260. [Google Scholar] [CrossRef]
- Currie, M.; Martin, P.; Blattmann, M.; Gordon, B.; Patterson, K. Final Report on Growing Tolerant Cultivars in a Psa Environment, VI1296. A Report Prepared for ZESPRI Group Limited; Plant & Food Research: Auckland, New Zealand, 2014. [Google Scholar]
- Gaskin, R.; Manktelow, D.; Cook, S.m.; May, B.; Horgan, D.; van Leeuwen, R. Novel Technologies to Deliver Protectant Sprays to Strung Canopies. A Report Prepared for ZESPRI Group Limited; Plant Protection Chemistry NZ: Rotorua, New Zealand, 2016. [Google Scholar]
- Vanneste, J.l.L.; Moffat, B.; Oldham, J. Survival of Pseudomonas syringae pv actinidiae on Cryptomeria japonica a nonhost plant used as shelter belts in kiwifruit orchards. N. Z. Plant Prot. 2012, 65, 1–7. [Google Scholar] [CrossRef]
- Pattemore, D.E.; Goodwin, R.M.; McBrydie, H.M.; Hoyte, S.M.; Vanneste, J.L. Evidence of the role of honey bees (Apis mellifera) as vectors of the bacterial plant pathogen Pseudomonas syringae. Australas. Plant Pathol. 2014, 43, 571–575. [Google Scholar] [CrossRef]
- Gupta, N.; Debnath, S.; Sharma, S.; Sharma, P.; Purohit, J. Role of nutrients in controlling the plant diseases in sustainable agriculture. In Agriculturally Important Microbes for Sustainable Agriculture: Applications in Crop Production and Protection; Meena, V.S., Mishra, P.K., Bisht, J.K., Pattanayak, A., Eds.; Springer: Singapore, 2017; Volume 2, pp. 217–262. [Google Scholar]
- Cesco, S.; Tolotti, A.; Nadalini, S.; Rizzi, S.; Valentinuzzi, F.; Mimmo, T.; Porfido, C.; Allegretta, I.; Giovannini, O.; Perazzolli, M.; et al. Plasmopara viticola infection affects mineral elements allocation and distribution in Vitis vinifera leaves. Sci. Rep. 2020, 10, 18759. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Zeng, Y.; Liu, P. Metabolic profiling reveals local and systemic responses of kiwifruit to Pseudomonas syringae pv. actinidiae. Plant Direct 2020, 4, e00297. [Google Scholar] [CrossRef] [PubMed]
- Groen, S.C.; Whiteman, N.K. The evolution of ethylene signaling in plant chemical ecology. J. Chem. Ecol. 2014, 40, 700–716. [Google Scholar] [CrossRef] [PubMed]
- Nunes da Silva, M.; Fernandes, A.P.; Vasconcelos, M.W.; Valente, L.C.M.; Carvalho, S.M.P. Influence of the nitrogen source on the tolerance of Actinidia chinensis to Pseudomonas syringae pv. actinidiae. Acta Hortic. 2022, 1332, 103–110. [Google Scholar] [CrossRef]
- Holmes, A. Effect of Soil Nutrition and Composition on the Susceptibility of Hayward and Hort16A to Pseudomonas syringae pv. Actinidiae. ZESPRI Innovation Project V11270 Final Report; ZESPRI: Tauranga, New Zealand, 2012. [Google Scholar]
- Gupta, N.; Bajpai, M.S.; Majumdar, R.; Mishra, P.K. Response of iodine on antioxidant levels of Glycine max L. grown under Cd stress. Adv. Biol. Res. 2015, 9, 40–48. [Google Scholar] [CrossRef]
- Gu, G.; Yang, S.; Yin, X.; Long, Y.; Ma, Y.; Li, R.; Wang, G. Sulfur induces resistance against canker caused by Pseudomonas syringae pv. actinidae via phenolic components Increase and morphological structure modification in the kiwifruit stems. Int. J. Mol. Sci. 2021, 22, 12185. [Google Scholar] [CrossRef]
- Zhang, Z.; Long, Y.; Yin, X.; Yang, S. Sulfur-induced resistance against Pseudomonas syringae pv. actinidiae via triggering salicylic acid signaling pathway in kiwifruit. Int. J. Mol. Sci. 2021, 22, 12710. [Google Scholar] [CrossRef]
- Cellini, A.; Fiorentini, L.; Buriani, G.; Yu, J.; Donati, I.; Cornish, D.A.; Novak, B.; Costa, G.; Vanneste, J.L.; Spinelli, F. Elicitors of the salicylic acid pathway reduce incidence of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidae. Ann. Appl. Biol. 2014, 165, 441–453. [Google Scholar] [CrossRef]
- Wurms, K.V.; Hardaker, A.J.; Ah Chee, A.; Bowen, J.; Phipps, J.; Taylor, J.; Jensen, D.; Cooney, J.; Wohlers, M.; Reglinski, T. Phytohormone and putative defense gene expression differentiates the response of ‘Hayward’ kiwifruit to Psa and Pfm infections. Front. Plant Sci. 2017, 8, 1366. [Google Scholar] [CrossRef]
- Michelotti, V.; Lamontanara, A.; Buriani, G.; Orrù, L.; Cellini, A.; Donati, I.; Vanneste, J.L.; Cattivelli, L.; Tacconi, G.; Spinelli, F. Comparative transcriptome analysis of the interaction between Actinidia chinensis var. chinensis and Pseudomonas syringae pv. actinidiae in absence and presence of acibenzolar-S-methyl. BMC Genom. 2018, 19, 585. [Google Scholar] [CrossRef]
- Wang, T.; Wang, G.; Jia, Z.-H.; Pan, D.-L.; Zhang, J.-Y.; Guo, Z.-R. Transcriptome analysis of kiwifruit in response to Pseudomonas syringae pv. actinidiae infection. Int. J. Mol. Sci. 2018, 19, 373. [Google Scholar] [CrossRef]
- Wang, T.; Jia, Z.H.; Zhang, J.Y.; Liu, M.; Guo, Z.R.; Wang, G. Identification and analysis of NBS-LRR genes in Actinidia chinensis genome. Plants 2020, 9, 1350. [Google Scholar] [CrossRef] [PubMed]
- Tahir, J.; Hoyte, S.; Bassett, H.; Brendolise, C.; Chatterjee, A.; Templeton, K.; Deng, C.; Crowhurst, R.; Montefiori, M.; Morgan, E.; et al. Multiple quantitative trait loci contribute to resistance to bacterial canker incited by Pseudomonas syringae pv. actinidiae in kiwifruit (Actinidia chinensis). Hortic. Res. 2019, 6, 101. [Google Scholar] [CrossRef] [PubMed]
- Nunes da Silva, M.; Carvalho, S.M.P.; Rodrigues, A.M.; Gómez-Cadenas, A.; António, C.; Vasconcelos, M.W. Defence-related pathways, phytohormones and primary metabolism are key players in kiwifruit plant tolerance to Pseudomonas syringae pv. actinidiae. Plant Cell Environ. 2022, 45, 528–541. [Google Scholar] [CrossRef] [PubMed]
- Wurms, K.; Gould, E.; Chee, A.A.; Taylor, J.; Curran, B.; Reglinski, T. Elicitor induction of defence genes and reduction of bacterial canker in kiwifruit. N. Z. Plant Prot. 2017, 70, 272–284. [Google Scholar] [CrossRef]
- Song, Y.; Sun, L.; Lin, M.; Chen, J.; Qi, X.; Hu, C.; Fang, J. Comparative transcriptome analysis of resistant and susceptible kiwifruits in response to Pseudomonas syringae pv. actinidiae during early infection. PLoS ONE 2019, 14, e0211913. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Liu, Y.; Zhang, D.; Ni, M.; Jia, B.; Heng, W.; Fang, Z.; Zhu, L.-W.; Liu, P. Transcriptomic and proteomic profiling reveal the key role of AcMYB16 in the response of Pseudomonas syringae pv. actinidiae in kiwifruit. Front. Plant Sci. 2021, 12, 756330. [Google Scholar] [CrossRef]
- Jing, Z.; Liu, Z. Genome-wide identification of WRKY transcription factors in kiwifruit (Actinidia spp.) and analysis of WRKY expression in responses to biotic and abiotic stresses. Genes Genom. 2018, 40, 429–446. [Google Scholar] [CrossRef]
- Michelotti, V.; Urbinati, G.; Gentile, A.; Lucioli, S.; Caboni, E.; Tacconi, G. Preliminary results on the development of a genome editing protocol in Actinidia chinensis var. chinensis as Psa resistance approach. Acta Hortic. 2022, 1332, 111–116. [Google Scholar] [CrossRef]
- Miao, L.; Genjia, T.; Yao, L.; Lian, X. Resistance mechanism of kiwifruit cultivars to Pseudomonas syringae pv. actinidiae. Acta Phytopathol. Sin. 2005, 32, 37–42. [Google Scholar]
- Petriccione, M.; Di Cecco, I.; Arena, S.; Scaloni, A.; Scortichini, M. Proteomic changes in Actinidia chinensis shoot during systemic infection with a pandemic Pseudomonas syringae pv. actinidiae strain. J. Proteom. 2013, 78, 461–476. [Google Scholar] [CrossRef]
- Petriccione, M.; Mastrobuoni, F.; Zampella, L.; Scortichini, M. Reference gene selection for normalization of RT-qPCR gene expression data from Actinidia deliciosa leaves infected with Pseudomonas syringae pv. actinidiae. Sci. Rep. 2015, 5, 16961. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Ethylene and 1-aminocyclopropane-1-carboxylate (ACC) in plant–bacterial interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Nunes da Silva, M.; Vasconcelos, M.W.; Pinto, V.; Balestra, G.M.; Mazzaglia, A.; Gomez-Cadenas, A.; Carvalho, S.M.P. Role of methyl jasmonate and salicylic acid in kiwifruit plants further subjected to Psa infection: Biochemical and genetic responses. Plant Physiol. Biochem. 2021, 162, 258–266. [Google Scholar] [CrossRef]
- Reglinski, T.; Vanneste, J.; Wurms, K.; Gould, E.; Spinelli, F.; Rikkerink, E. Using fundamental knowledge of induced resistance to develop control strategies for bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidiae. Front. Plant Sci. 2013, 4, 24. [Google Scholar] [CrossRef]
- Liu, L.; Sonbol, F.-M.; Huot, B.; Gu, Y.; Withers, J.; Mwimba, M.; Yao, J.; He, S.Y.; Dong, X. Salicylic acid receptors activate jasmonic acid signalling through a non-canonical pathway to promote effector-triggered immunity. Nat. Commun. 2016, 7, 13099. [Google Scholar] [CrossRef] [Green Version]
- Reglinski, T.; Wurms, K.; Vanneste, J.; Ah Chee, A.; Yu, J.; Oldham, J.; Cornish, D.; Cooney, J.; Jensen, D.; Trower, T. Transient changes in defence gene expression and phytohormone cInduced by acibenzolar-S-methyl in glasshouse and orchard grown kiwifruit. Front. Agron. 2022, 3, 831172. [Google Scholar] [CrossRef]
- Stroud, E.A.; Rikkerink, E.H.A.; Jayaraman, J.; Templeton, M.D. Actigard™ induces a defence response to limit Pseudomonas syringae pv. actinidiae in Actinidia chinensis var. chinensis ‘Hort16A’ tissue culture plants. Sci. Hortic. 2022, 295, 110806. [Google Scholar] [CrossRef]
- Sun, L.-M.; Fang, J.-B.; Zhang, M.; Qi, X.-J.; Lin, M.-M.; Chen, J.-Y. Molecular cloning and functional analysis of the NPR1 homolog in kiwifruit (Actinidia eriantha). Front. Plant Sci. 2020, 11, 551201. [Google Scholar] [CrossRef]
- Melotto, M.; Underwood, W.; Koczan, J.; Nomura, K.; He, S.Y. Plant stomata function in innate immunity against bacterial invasion. Cell 2006, 126, 969–980. [Google Scholar] [CrossRef]
- Ton, J.; Flors, V.; Mauch-Mani, B. The multifaceted role of ABA in disease resistance. Trends Plant Sci. 2009, 14, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Munemasa, S.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Involvement of endogenous abscisic acid in methyl jasmonate-induced stomatal closure in Arabidopsis. Plant Physiol. 2011, 156, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Cellini, A.; Fiorentini, L.; Dharmaraj, K.; Vanneste, J.L.; Cristescu, S.M.; Harren, F.J.M.; Costa, G.; Spinelli, F. Evidences of Ethylene as a Virulence Factor for Pseudomonas syringae pv. actinidiae and Field Measures to Limit the Kiwifruit Plant Bacterial Canker. In Proceedings of the IXth International Conference on the Plant Hormone Ethylene, Rotorua, New Zealand, 19–23 March 2012. [Google Scholar]
- Collina, M.; Donati, I.; Bertacchini, E.; Brunelli, A.; Spinelli, F. Greenhouse assays on the control of the bacterial canker of kiwifruit (Pseudomonas syringae pv. actinidiae). J. Berry Res. 2016, 6, 407–415. [Google Scholar] [CrossRef]
- Lu, Y.; Yao, J. Chloroplasts at the crossroad of photosynthesis, pathogen infection and plant defense. Int. J. Mol. Sci. 2018, 19, E3900. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Y.; Li, L.; Li, D.; Zhang, Q.; Guo, Y.; Wang, S.; Zhong, C.; Huang, H. Whole transcriptome sequencing of Pseudomonas syringae pv. actinidiae-infected kiwifruit plants reveals species-specific interaction between long non-coding RNA and coding genes. Sci. Rep. 2017, 7, 4910. [Google Scholar] [CrossRef]
- Kaji, R.; Yariuchi, R.; Fujii, Y.; Taniguchi, S.; Uji, Y.; Suzuki, G.; Kashihara, K.; Kisaki, G.; Suezawa, K.; Ohtani, M. Expression analysis of defense-related genes in wild kiwifruit (Actinidia rufa) tolerant to bacterial canker. J. Gen. Plant Pathol. 2021, 87, 361–365. [Google Scholar] [CrossRef]
- Shelp, B.J.; Aghdam, M.S.; Flaherty, E.J. γ-Aminobutyrate (GABA) regulated plant defense: Mechanisms and opportunities. Plants 2021, 10, 1939. [Google Scholar] [CrossRef]
- Miao, L.; Genjia, T.; Yao, L.; Kejian, D.; Zhulong, C.; Yun, L. Relationships between the contents of phenolics, soluble proteins in plants of kiwifruit cultivars and their resistance to kiwifruit bacterial canker by Pseudomonas syringae pv. actinidiae. Plant Prot. 2009, 35, 37–41. [Google Scholar]
- Cheng, C.-H. Inheritance of resistance to Pseudomonas syringae pv. actinidiae and genetic correlations with fruit characters in a diploid Actinidia chinensis (kiwifruit) population. Euphytica 2014, 198, 305–315. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes da Silva, M.; Santos, M.G.; Vasconcelos, M.W.; Carvalho, S.M.P. Mitigation of Emergent Bacterial Pathogens Using Pseudomonas syringae pv. actinidiae as a Case Study—From Orchard to Gene and Everything in Between. Crops 2022, 2, 351-377. https://doi.org/10.3390/crops2040025
Nunes da Silva M, Santos MG, Vasconcelos MW, Carvalho SMP. Mitigation of Emergent Bacterial Pathogens Using Pseudomonas syringae pv. actinidiae as a Case Study—From Orchard to Gene and Everything in Between. Crops. 2022; 2(4):351-377. https://doi.org/10.3390/crops2040025
Chicago/Turabian StyleNunes da Silva, Marta, Miguel G. Santos, Marta W. Vasconcelos, and Susana M. P. Carvalho. 2022. "Mitigation of Emergent Bacterial Pathogens Using Pseudomonas syringae pv. actinidiae as a Case Study—From Orchard to Gene and Everything in Between" Crops 2, no. 4: 351-377. https://doi.org/10.3390/crops2040025