
Alleviating Salt Stress in Tomatoes through Seed Priming with Polyethylene Glycol and Sodium Chloride Combination



Abstract
:1. Introduction
2. Results
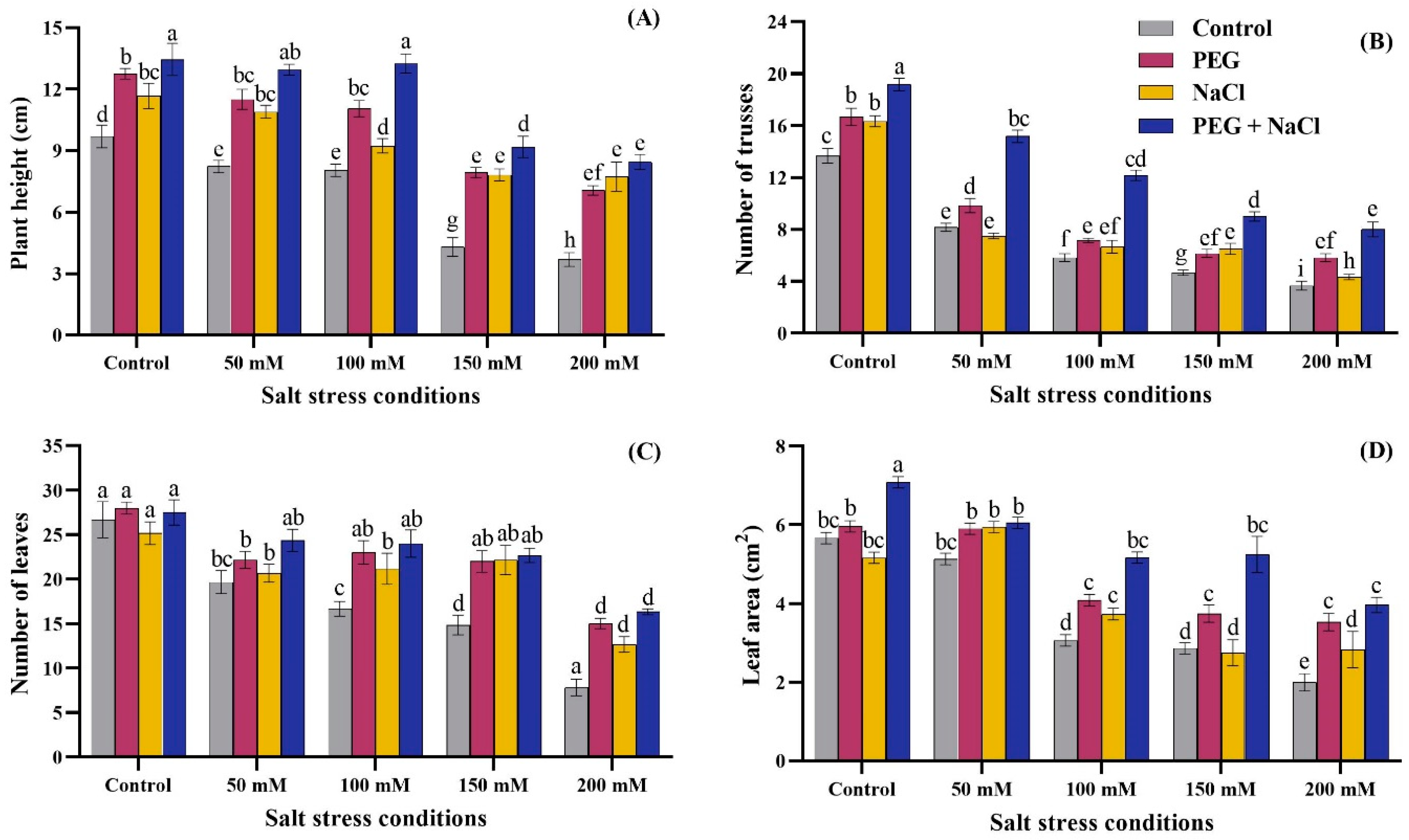
2.1. Growth Parameters
2.2. Photosynthetic Attributes
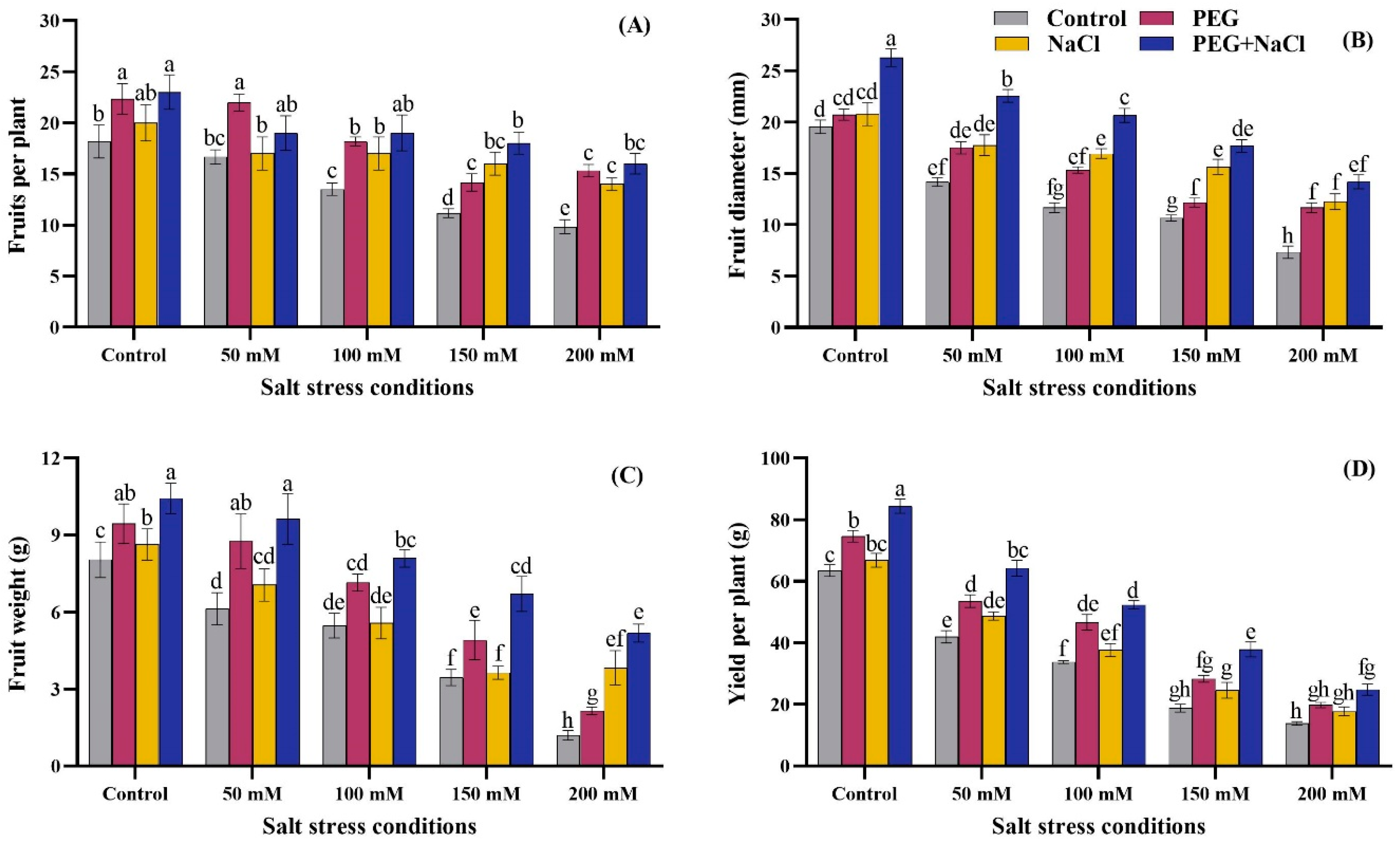
2.3. Yield Attributes
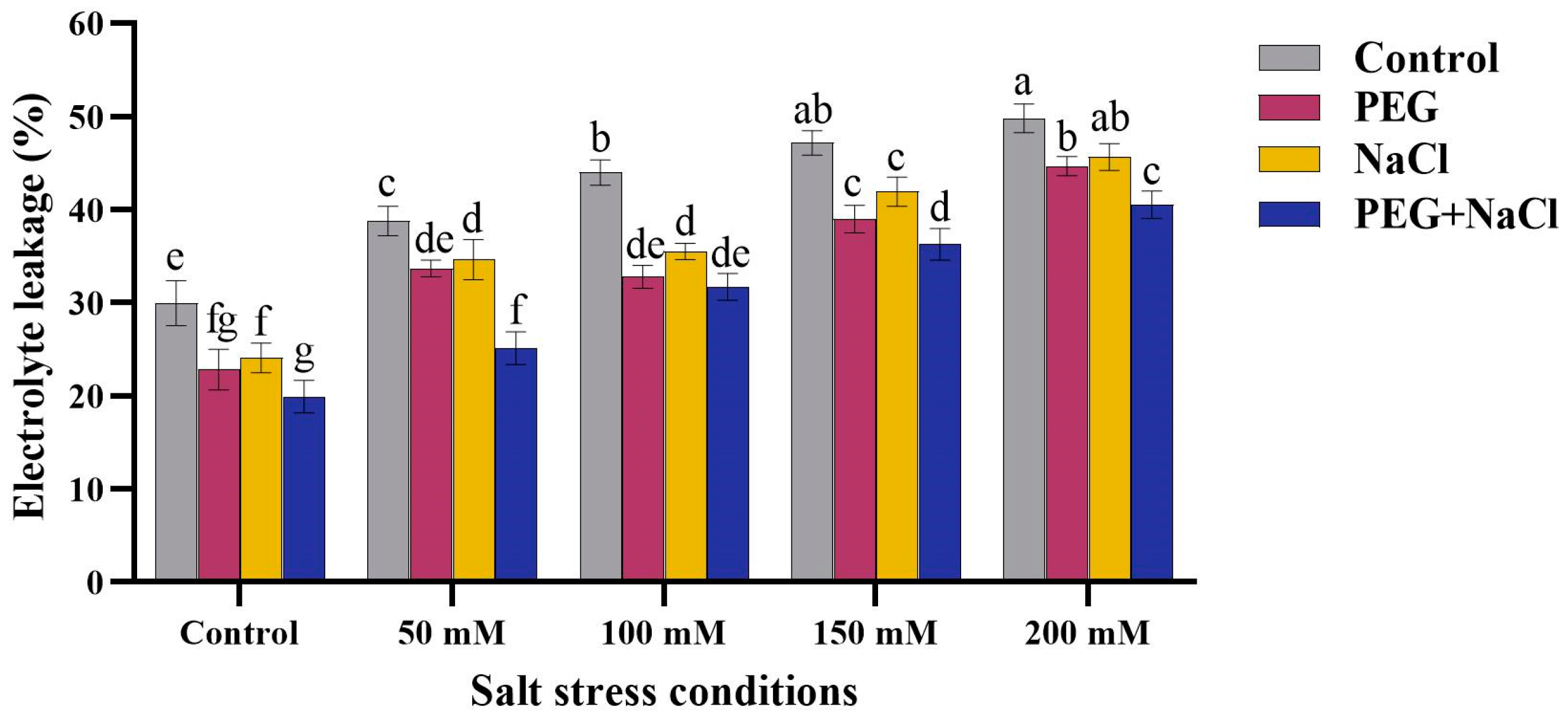
2.4. Electrolyte Leakage
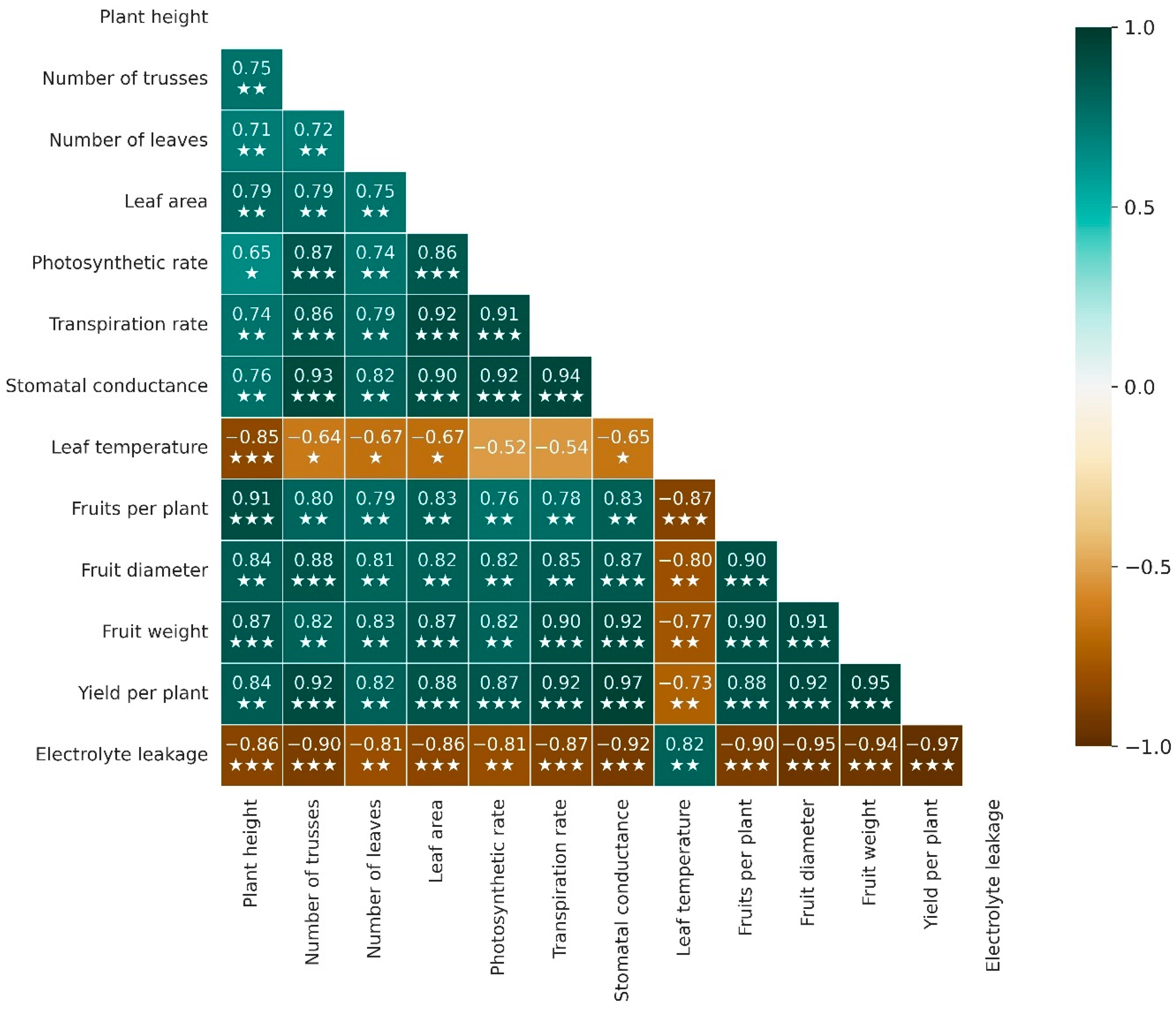
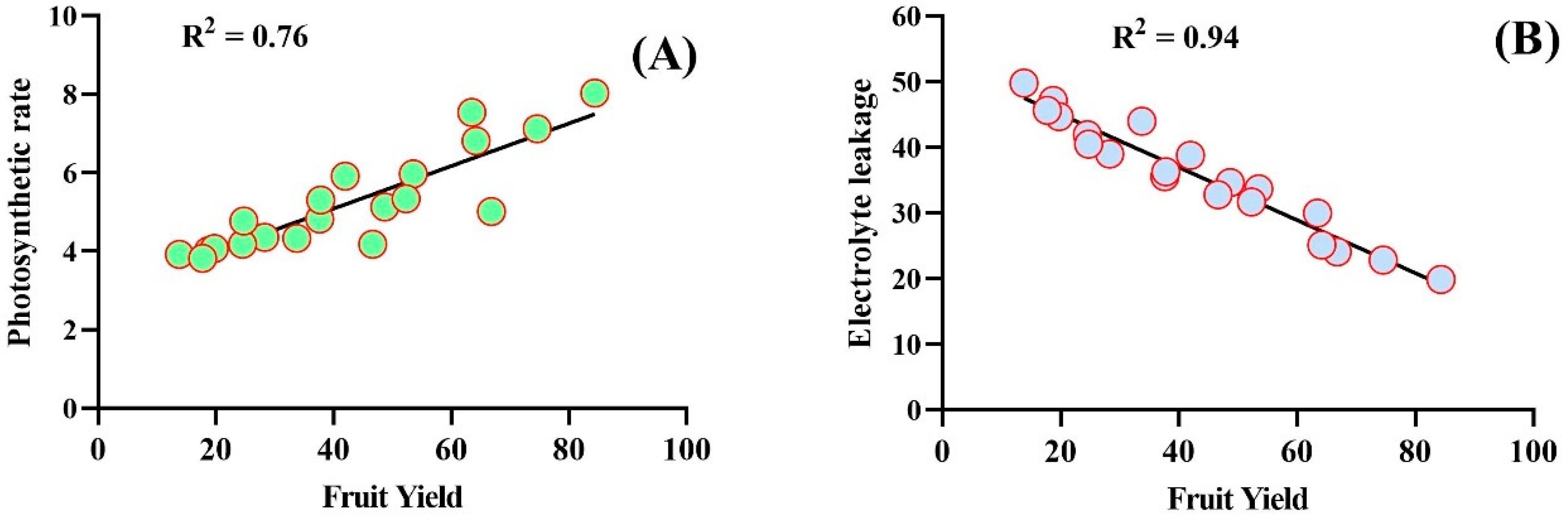
2.5. Correlation Anaylsis
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Cultivation and Application of Treatments
4.3. Growth Parameters
4.4. Photosynthetic Traits
4.5. Fruit Yield Attributes
4.6. Determination of Electrolyte Leakage
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, L.Y.; De Zhu, H.; De Yin, K.; Du, J.D.; Zhang, Y.X. Seed Priming Mitigates the Effects of Saline-Alkali Stress in Soybean Seedlings. Chil. J. Agric. Res. 2017, 77, 118–125. [Google Scholar] [CrossRef]
- Mohammadi Alagoz, S.; Hadi, H.; Toorchi, M.; Pawłowski, T.A.; Asgari Lajayer, B.; Price, G.W.; Farooq, M.; Astatkie, T. Morpho-Physiological Responses and Growth Indices of Triticale to Drought and Salt Stresses. Sci. Rep. 2023, 13, 8896. [Google Scholar] [CrossRef] [PubMed]
- Habibi, N.; Aryan, S.; Amin, M.W.; Sanada, A.; Terada, N.; Koshio, K. Potential Benefits of Seed Priming under Salt Stress Conditions on Physiological, and Biochemical Attributes of Micro-Tom Tomato Plants. Plants 2023, 12, 2187. [Google Scholar] [CrossRef] [PubMed]
- Habibi, N.; Fakoor, M.Y.; Faqiri, S.M.; Sharaf, Z.; Hotak, M.S.; Danishyar, N.; Haris, M.M.; Osmani, K.S.; Shinohara, T.; Terada, N.; et al. Enhancing Salinity Tolerance in Tomatoes at the Reproductive Stage by Increasing Pollen Viability. Bionatura 2023, 8, 25. [Google Scholar] [CrossRef]
- Habibi, N.; Sediqui, N.; Terada, N.; Sanada, A.; Koshio, K. Effects of Salinity on Growth, Physiological and Biochemical Responses of Tomato. J. ISSAAS 2021, 27, 14–28. [Google Scholar]
- Oho, K.; Habibi, N.; Marie, T.; Silva, B.; Terada, N.; Sanada, A.; Shinohara, T.; Gemma, H.; Koshio, K. Elucidation of physicochemical changes in fruit development of “sabara” jaboticaba (Plinia cauliflora (Mart.) Kausel). J. ISSAAS 2022, 28, 34–50. [Google Scholar]
- Kamanga, R.M.; Ndakidemi, P.A. Cultivation of Tomato under Dehydration and Salinity Stress: Unravelling the Physiology and Alternative Tolerance Options. In Tomato—From Cultivation to Procession Technology; IntechOpen: London, UK, 2022; pp. 1–17. [Google Scholar]
- Kaveh, H.; Nemati, H.; Farsi, M.; Vatandoost Jartoodeh, S. How Salinity Affect Germination and Emergence of Tomato Lines. J. Biol. Environ. Sci. 2011, 5, 159–163. [Google Scholar]
- Raziq, A.; Wang, Y.; Mohi Ud Din, A.; Sun, J.; Shu, S.; Guo, S. A Comprehensive Evaluation of Salt Tolerance in Tomato (Var. Ailsa Craig): Responses of Physiological and Transcriptional Changes in RBOH’s and ABA Biosynthesis and Signalling Genes. Int. J. Mol. Sci. 2022, 23, 1603. [Google Scholar] [CrossRef]
- Loudari, A.; Mayane, A.; Zeroual, Y.; Colinet, G.; Oukarroum, A. Photosynthetic Performance and Nutrient Uptake under Salt Stress: Differential Responses of Wheat Plants to Contrasting Phosphorus Forms and Rates. Front. Plant Sci. 2022, 13, 1038672. [Google Scholar] [CrossRef]
- Saghafi, D.; Ghorbanpour, M.; Shirafkan Ajirloo, H.; Asgari Lajayer, B. Enhancement of Growth and Salt Tolerance in Brassica napus L. Seedlings by Halotolerant Rhizobium Strains Containing ACC-Deaminase Activity. Plant Physiol. Rep. 2019, 24, 225–235. [Google Scholar] [CrossRef]
- Parra, M.; Albacete, A.; Martínez-Andújar, C.; Pérez-Alfocea, F. Increasing Plant Vigour and Tomato Fruit Yield under Salinity by Inducing Plant Adaptation at the Earliest Seedling Stage. Environ. Exp. Bot. 2007, 60, 77–85. [Google Scholar] [CrossRef]
- Agius, C.; von Tucher, S.; Rozhon, W. The Effect of Salinity on Fruit Quality and Yield of Cherry Tomatoes. Horticulturae 2022, 8, 59. [Google Scholar] [CrossRef]
- Biswas, S.; Seal, P.; Majumder, B.; Biswas, A.K. Efficacy of Seed Priming Strategies for Enhancing Salinity Tolerance in Plants: An Overview of the Progress and Achievements. Plant Stress 2023, 9, 100186. [Google Scholar] [CrossRef]
- ul Sahar, N.; Khatoon, N.; Mangrio, A.M.; Rind, N.A.; Rafiq, M.; Dahot, M.U. The Halopriming of Seeds Improves the Germination, Growth, Physiological and Phytochemical Attributes of Tomato under Saline Conditions. Emir. J. Food Agric. 2023, 35, 48–58. [Google Scholar] [CrossRef]
- Alam, M.S.; Tester, M.; Fiene, G.; Mousa, M.A.A. Early Growth Stage Characterization and the Biochemical Responses for Salinity Stress in Tomato. Plants 2021, 10, 712. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Zhao, S.; Arif, S.; Zhao, Y.; Ming, L.C.; Huang, D. Seed Priming Technology as a Key Strategy to Increase Crop Plant Production under Adverse Environmental Conditions. J. Agric. Hortic. Res. 2022, 5, 27–46. [Google Scholar] [CrossRef]
- Nakaune, M.; Tsukazawa, K.; Uga, H.; Asamizu, E.; Imanishi, S.; Matsukura, C.; Ezura, H. Low Sodium Chloride Priming Increases Seedling Vigor and Stress Tolerance to Ralstonia Solanacearum in Tomato. Plant Biotechnol. 2012, 29, 9–18. [Google Scholar] [CrossRef]
- Bousba, R.; Bounar, R.; Sedrati, N.; Lekhal, R.; Hamla, C.; Rached-Kanouni, M. Effects of Osmotic Stress Induced by Polyethylene Glycol (Peg) 6000 and Mannitol on Seed Germination and Seedling Growth of Durum Wheat. J. Bioresour. Manag. 2021, 8, 57–66. [Google Scholar] [CrossRef]
- Alharbi, K.; Al-Osaimi, A.A.; Alghamdi, B.A. Sodium Chloride (NaCl)-Induced Physiological Alteration and Oxidative Stress Generation in Pisum sativum (L.): A Toxicity Assessment. ACS Omega 2022, 7, 20819–20832. [Google Scholar] [CrossRef]
- Wang, R.; Li, C.; Zeng, L.; Liu, L.; Xi, J.; Li, J. Polyethylene Glycol Priming Enhances the Seed Germination and Seedling Growth of Scutellaria Baicalensis Georgi under Salt Stress. Plants 2024, 13, 565. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of Reactive Oxygen Species and Antioxidant Defense in Plants under Salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef]
- Hossain, M.S.; Alam, M.U.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Al Mahmud, J.; Fujita, M. Use of Iso-Osmotic Solution to Understand Salt Stress Responses in Lentil (Lens culinaris Medik.). S. Afr. J. Bot. 2017, 113, 346–354. [Google Scholar] [CrossRef]
- El-Saifi, S.K.; Ahmed, H.M.; Hasan, S.M.; Morsi, M.M.; El-Shatoury, R.S. Seed Priming Influences Seed Germination and Seedling Growth of Tomato under Different Salinity Levels. J. Plant Prod. 2010, 1, 159–170. [Google Scholar] [CrossRef]
- Mirabi, E.; Hasanabadi, M. Effect of Seed Priming on Some Characteristic of Seedling and Seed Vigor of Tomato (Lycopersicun esculentum). J. Adv. Lab. Res. Biol. 2012, 3, 237–240. [Google Scholar]
- Rouhi, H.R.; Aboutalebian, M.A.; Karimi, F.; Saman, M. Change in Several Antioxidant Enzymes Activity of Berseem Clover (Trifolium alexandrinum L.) by Priming Change in Several Antioxidant Enzymes Activity of Berseem Clover (Trifolium alexandrinum L.) by Priming. Int. J. Agric. Sci. 2012, 2, 237–243. [Google Scholar]
- Tiwari, M.; Singh, P. Plant Defense Priming: A New Tool for Sustainable Global Food Security. In Agricultural Innovations and Sustainability; Agrobios Research: Rajasthan, India, 2021; pp. 133–153. [Google Scholar]
- Sheteiwy, M.S.; Ulhassan, Z.; Qi, W.; Lu, H.; AbdElgawad, H.; Minkina, T.; Sushkova, S.; Rajput, V.D.; El-Keblawy, A.; Jośko, I.; et al. Association of Jasmonic Acid Priming with Multiple Defense Mechanisms in Wheat Plants under High Salt Stress. Front. Plant Sci. 2022, 13, 886862. [Google Scholar] [CrossRef] [PubMed]
- Devika, O.S.; Singh, S.; Sarkar, D.; Barnwal, P.; Suman, J.; Rakshit, A. Seed Priming: A Potential Supplement in Integrated Resource Management under Fragile Intensive Ecosystems. Front. Sustain. Food Syst. 2021, 5, 654001. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hossain, M.; Hossain, K.F.B.; Sikder, M.T.; Shammi, M.; Rasheduzzaman, M.; Hossain, M.A.; Alam, A.M.; Uddin, M.K. Effects of NaCl-Salinity on Tomato (Lycopersicon esculentum Mill.) Plants in a Pot Experiment. Open Agric. 2018, 3, 578–585. [Google Scholar] [CrossRef]
- Bacha, H.; Tekaya, M.; Drine, S.; Guasmi, F.; Touil, L.; Enneb, H.; Triki, T.; Cheour, F.; Ferchichi, A. Impact of Salt Stress on Morpho-Physiological and Biochemical Parameters of Solanum lycopersicum cv. Microtom Leaves. S. Afr. J. Bot. 2017, 108, 364–369. [Google Scholar] [CrossRef]
- Rafique, N.; Hammad Raza, S.; Qasim, M.; Iqbal, N. Pre-sowing application of ascorbic acid and salicylic acid to seed of pumpkin and seedling response to salt. Pak. J. Bot. 2011, 43, 2677–2682. [Google Scholar]
- Munns, R. Comparative Physiology of Salt and Water Stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, Z.; Yuan, L.; Yin, C.; Cheng, J.; Wang, L.; Huang, J.; Zhang, H. Osmopriming Improves Tomato Seed Vigor under Aging and Salinity Stress. Afr. J. Biotechnol. 2012, 11, 6305–6311. [Google Scholar] [CrossRef]
- Pradhan, N.; Prakash, P.; Tiwari, S.K. Osmopriming of Tomato Genotypes with Polyethylene Glycol 6000 Induces Tolerance to Salinity Stress Osmopriming of Tomato Genotypes with Polyethylene Glycol 6000 Induces. Trends Biosci. 2015, 7, 4412–4417. [Google Scholar]
- Ali, S.; Ullah, S.; Khan, M.N.; Khan, W.M.; Razak, S.A.; Wahab, S.; Hafeez, A.; Khan Bangash, S.A.; Poczai, P. The Effects of Osmosis and Thermo-Priming on Salinity Stress Tolerance in Vigna radiata L. Sustainability 2022, 14, 12924. [Google Scholar] [CrossRef]
- Hussein, M.; El-Gereadly, H.M.; EL-Desuki, M. Role of Puterscine in Resistance to Salinity of Pea Plants (Pisum sativum L.). J. Appl. Sci. Res. 2006, 2, 598–604. [Google Scholar]
- Zhang, P.; Senge, M.; Dai, Y. Effects of Salinity Stress at Different Growth Stages on Tomato Growth, Yield, and Water-Use Efficiency. Commun. Soil Sci. Plant Anal. 2017, 48, 624–634. [Google Scholar] [CrossRef]
- Cano, E.A.; Bolarín, M.C.; Pérez-Alfocea, F.; Caro, M. Effect of NaCl Priming on Increased Salt Tolerance in Tomato. J. Hortic. Sci. 1991, 66, 621–628. [Google Scholar] [CrossRef]
- Alves, R.d.C.; Rossatto, D.R.; da Silva, J.d.S.; Checchio, M.V.; de Oliveira, K.R.; Oliveira, F.d.A.; de Queiroz, S.F.; da Cruz, M.C.P.; Gratão, P.L. Seed Priming with Ascorbic Acid Enhances Salt Tolerance in Micro-Tom Tomato Plants by Modifying the Antioxidant Defense System Components. Biocatal. Agric. Biotechnol. 2021, 31, 101927. [Google Scholar] [CrossRef]
- Delian, E.; Badulescu, L.; Bădulescu, L.; Dobrescu, A.; Chira, L.; Lagunovschi-Luchian, V. A Brief Overview of Seed Priming Benefits in Tomato. Rom. Biotechnol. Lett. 2017, 22, 12505–12513. [Google Scholar]
- Jakab, G.; Ton, J.; Flors, V.; Zimmerli, L.; Métraux, J.P.; Mauch-Mani, B. Enhancing Arabidopsis Salt and Drought Stress Tolerance by Chemical Priming for Its Abscisic Acid Responses. Plant Physiol. 2005, 139, 267–274. [Google Scholar] [CrossRef]
- Anwar, A.; Yu, X.; Li, Y. Seed Priming as a Promising Technique to Improve Growth, Chlorophyll, Photosynthesis and Nutrient Contents in Cucumber Seedlings. Not. Bot. Horti Agrobot. Cluj Napoca 2020, 48, 116–127. [Google Scholar] [CrossRef]
- Jisha, K.C.; Puthur, J.T. Halopriming of Seeds Imparts Tolerance to NaCl and PEG Induced Stress in Vigna radiata (L.) Wilczek Varieties. Physiol. Mol. Biol. Plants 2014, 20, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Michel, B.E.; Kaufmann, M.R. The Osmotic Potential of Polyethylene Glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef] [PubMed]
- Viera, W.; Samaniego, I.; Camacho, D.; Habibi, N.; Ron, L.; Sediqui, N.; Álvarez, J.; Viteri, P.; Sotomayor, A.; Merino, J.; et al. Phytochemical Characterization of a Tree Tomato (Solanum betaceum Cav.) Breeding Population Grown in the Inter-Andean Valley of Ecuador. Plants 2022, 11, 268. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity | Priming | E | Gs | (°C) | |
|---|---|---|---|---|---|
| Control | Control | 7.68 ± 0.15 ab | 0.41 ± 0.024 c–e | 0.37 ± 0.01 b | 27.05 ± 1.05 c–e |
| PEG | 7.73 ± 0.54 ab | 0.45 ± 0.023 bc | 0.39 ± 0.014 b | 20.46 ± 0.69 gh | |
| NaCl | 5.38 ± 0.33 ef | 0.44 ± 0.012 cd | 0.36 ± 0.019 b | 22.43 ± 0.68 f–h | |
| PEG plus NaCl | 8.49 ± 0.44 a | 0.52 ± 0.014 a | 0.46 ± 0.011 a | 19.97 ± 1.59 h | |
| *** | *** | ** | *** | ||
| 50 mM | Control | 6.06 ± 0.42 c–e | 0.35 ± 0.012 fg | 0.27 ± 0.015 cd | 30.75 ± 1.05 bc |
| PEG | 6.48 ± 0.45 cd | 0.41 ± 0.013 de | 0.28 ± 0.006 c | 24.33 ± 0.48 e–g | |
| NaCl | 5.5 ± 0.35 d–f | 0.38 ± 0.012 ef | 0.26 ± 0.014 c–e | 25.76 ± 0.68 d–f | |
| PEG plus NaCl | 7.14 ± 0.38 bc | 0.49 ± 0.024 ab | 0.36 ± 0.016 b | 22.52 ± 1.12 f–h | |
| *** | *** | *** | *** | ||
| 100 mM | Control | 4.57 ± 0.25 f–k | 0.22 ± 0.02 hi | 0.2 ± 0.013 fg | 29.63 ± 0.85 b–d |
| PEG | 4.86 ± 0.59 f–i | 0.25 ± 0.012 h | 0.22 ± 0.009 d–g | 24.07 ± 2.97 e–g | |
| NaCl | 4.64 ± 0.19 f–k | 0.23 ± 0.013 h | 0.19 ± 0.013 fg | 27.21 ± 1.2 c–e | |
| PEG plus NaCl | 5.24 ± 0.35 e–h | 0.34 ± 0.012 fg | 0.27 ± 0.008 c | 22.53 ± 2.11 f–h | |
| ** | *** | *** | *** | ||
| 150 mM | Control | 4.3 ± 0.33 g–k | 0.19 ± 0.013 i–k | 0.15 ± 0.018 hi | 32.77 ± 1.41 ab |
| PEG | 4.74 ± 0.34 f–j | 0.25 ± 0.014 h | 0.18 ± 0.015 gh | 25.32 ± 0.76 ef | |
| NaCl | 4.06 ± 0.11 i–k | 0.21 ± 0.015 h–j | 0.15 ± 0.013 h–j | 27.22 ± 0.9 c–e | |
| PEG plus NaCl | 5.27 ± 0.24 e–g | 0.32 ± 0.011 g | 0.22 ± 0.017 e–g | 23.2 ± 1.17 e–h | |
| ** | *** | *** | *** | ||
| 200 mM | Control | 3.56 ± 0.35 k | 0.15 ± 0.022 k | 0.09 ± 0.02 k | 35.1 ± 1.72 a |
| PEG | 4.18 ± 0.15 h–k | 0.18 ± 0.021 jk | 0.13 ± 0.011 ij | 24.5 ± 0.87 e–g | |
| NaCl | 3.71 ± 0.27 jk | 0.17 ± 0.011 jk | 0.1 ± 0.023 jk | 25.6 ± 1.21 d–f | |
| PEG plus NaCl | 4.69 ± 0.35 f–j | 0.24 ± 0.031 h | 0.23 ± 0.014 d–f | 23.3 ± 1.18 e–h | |
| *** | ** | *** | *** |
| Salt Stress Treatments | Priming Treatments | Amount |
|---|---|---|
| 0 mM | Control | 0 MPa |
| 0 mM | PEG | −1.2 MPa |
| 0 mM | NaCl | 200 mM |
| 0 mM | PEG + NaCl | −1.2 MPa plus 200 mM |
| 50 mM | Control | 0 MPa |
| 50 mM | PEG | −1.2 MPa |
| 50 mM | NaCl | 200 mM |
| 50 mM | PEG + NaCl | −1.2 MPa plus 200 mM |
| 100 mM | Control | 0 MPa |
| 100 mM | PEG | −1.2 MPa |
| 100 mM | NaCl | 200 mM |
| 100 mM | PEG + NaCl | −1.2 MPa plus 200 mM |
| 150 mM | Control | 0 MPa |
| 150 mM | PEG | −1.2 MPa |
| 150 mM | NaCl | 200 mM |
| 150 mM | PEG + NaCl | −1.2 MPa plus 200 mM |
| 200 mM | Control | 0 MPa |
| 200 mM | PEG | −1.2 MPa |
| 200 mM | NaCl | 200 mM |
| 200 mM | PEG + NaCl | −1.2 MPa plus 200 mM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habibi, N.; Terada, N.; Sanada, A.; Koshio, K. Alleviating Salt Stress in Tomatoes through Seed Priming with Polyethylene Glycol and Sodium Chloride Combination. Stresses 2024, 4, 210-224. https://doi.org/10.3390/stresses4020012
Habibi N, Terada N, Sanada A, Koshio K. Alleviating Salt Stress in Tomatoes through Seed Priming with Polyethylene Glycol and Sodium Chloride Combination. Stresses. 2024; 4(2):210-224. https://doi.org/10.3390/stresses4020012
Chicago/Turabian StyleHabibi, Nasratullah, Naoki Terada, Atsushi Sanada, and Kaihei Koshio. 2024. "Alleviating Salt Stress in Tomatoes through Seed Priming with Polyethylene Glycol and Sodium Chloride Combination" Stresses 4, no. 2: 210-224. https://doi.org/10.3390/stresses4020012