

CRISPR-Cas Genome Editing for Insect Pest Stress Management in Crop Plants

 ,
, 
Abstract
:1. Introduction
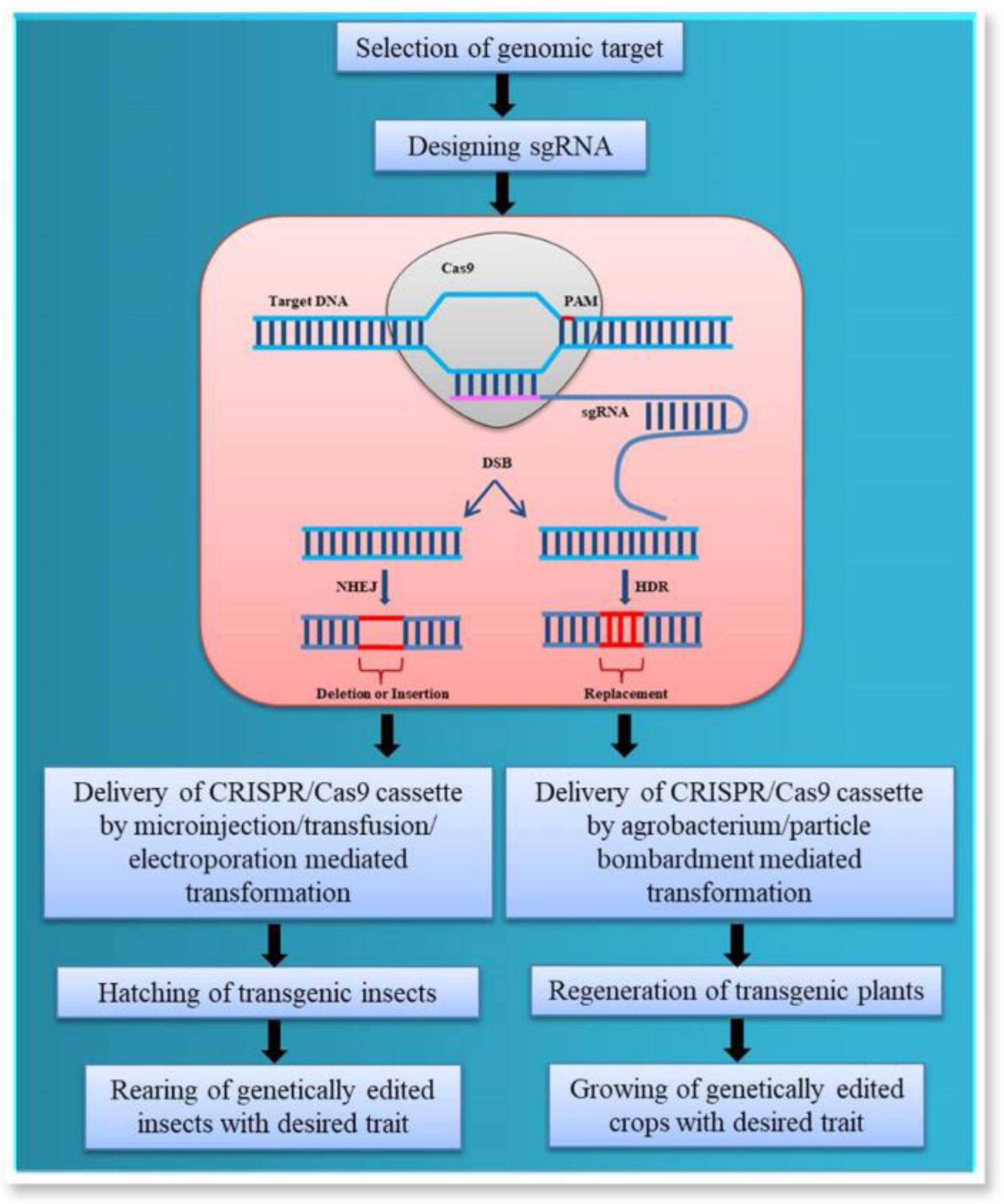
2. CRISPR-Cas System and Its Mechanism of Genome Editing
3. CRISPR-Cas Genome Editing in Agriculture for the Management of Insect Pests
3.1. CRISPR-Cas Genome Editing in Insects

{kind=link}
| Name of Insect | Target Gene | Editing | Outcome |
|---|---|---|---|
| Drosophila melanogaster | Yellow | Knockout, Knock-In | Generated designer flies [33] |
| Rosy and DSH3PX1 | Knockout, Knock-In | Executed efficient and complex genomic manipulations [33] | |
| LUBEL | Knockout | Reduced survival rate [44] | |
| Chitin synthase 1 | Substitution | Controlled insect population and resistance to various insecticides [45] | |
| Nicotinic acetylcholine receptor α6 | Substitution | Controlled insect population and resistance to various insecticides [46] | |
| Scsa | Knockout | Reduced normal growth [47] | |
| Kdr | Knockout | Reduce insecticide resistance [48] | |
| Ast, Eh, capa, Ccap, Crz, npf, Mip, mir-219, mir-315, and white | Knockout | Targeted mutagenesis [61] | |
| Yellow | Knockout | Effectively targeted mutagenesis [62] | |
| Yellow and white | Knockout | Highly efficient and varied genome-editing efficiencies [63] | |
| Yellow and rosy | Knockout, Knock-In | First report using the CRISPR/Cas9 system to mediate efficient genome engineering in Drosophila [64] | |
| Alk | Knockout | Establishing mutations [65] | |
| TpnC | Knockout | Confirmed that the myofibril assembly wasrelated to TpnC gene [66] | |
| Wntless | Knockout | Amplified the cleavage Efficiency [67] | |
| Yellow, white, and tan | Knock-In | Attaining single or multiple allelic substitutions [68] | |
| Act5C, lig4, and mus308 | Knockout, Knock-In | Genome editing in Drosophila S2 cells [69] | |
| Mod(mdg4) | Knockout | Validation of a functional gene involved in trans-splicing that influenced the development in flies [70] | |
| Fdl | Knockout | Analyzing or manipulating protein glycosylation pathways [71] | |
| Chameau, CG4221, and CG5961 mRNA | Knock-In | A problem associated with “ends-in” recombination was resolved [72] | |
| Clamp | Knockout | The expression of a sex-specific gene was regulated [73] | |
| Dα6 | Knock-In | Resistance to spinosad [46] | |
| Yellow | Knock-In | Heterozygous recessive mutation was converted to homozygous loss of function [74] | |
| Ebony, yellow, wg, wls, Lis1, and Se | Knockout, Knock-In | Non-transgenic individuals exhibited less efficient knock-in than transgenic individuals did [75] | |
| Ebony, yellow, and white | Knockout, Knock-In | Enhanced efficiency of gene targeting [76] | |
| Ebony, yellow, and vermilion | Knockout, Knock-In | Donor template and sgRNA plasmids were injected into Cas9 transgenic embryos in Drosophila [77] | |
| White and piwi | Knockout, Knock-In | Prevented off-target effects during the generation of indel mutants [77] | |
| Salm | Knock-In | Flexible modification of fly genome [78] | |
| Yellow, notch, bam, nos, ms(3)k81, and cid | Knockout | Temporally and spatially inhibited gene expression [79] | |
| Ms(3)k81, white, and yellow | Knockout, Knock-In | CRISPR-mediated genome editing was shown in Drosophila [80] | |
| EGFP and mRFP | Knockout | Induction of mutations [81] | |
| Ebony, yellow, wingless, and wnt | Knockout, Knock-In | Different patterns of expression [82] | |
| Drosophila subobscura | Yellow and white | Knockout | Gene functions were analyzed in a non-model Drosophila species [83] |
| Drosophila suzukii | White (w) and sex lethal (Sxl) | Knockout | Controlled insect population and resistance to various insecticides [27] |
| DsRed (red fluorescence protein) | Knock-In | Studied sexing and monitoring [84] | |
| White (w-) | Knockout | Absence of mating and copulation failure [85] | |
| Spodoptera exigua | Seα6 | Knockout | Resistance to insecticides [23] |
| Ryanodine receptor | Substitution | Controlled insect population and resistance to various insecticides [49] | |
| CYP9A186 gene | Knockout | Susceptibility to emamectin benzoate (EB) [50] | |
| P-glycoprotein gene | Knockout | Susceptibility to abamectin and emamectin benzoate [51] | |
| a-6-nicotinic acetylcholine receptor (nAchR) | Knockout | Resistance to spinosyn insecticides [23,65,66] | |
| Spodoptera littoralis | Orco | Knockout | Reduced survival rate [32] |
| Spodoptera litura | Abdominal-A (slabd-A) | Knockout | Defected body segmentation and pigmentation [42] |
| SlitPBP3 | Knockout | Destroyed pest insect mating [86] | |
| SlitBLOS2 | Knockout | Coloration of the integuments, a marker gene for functional studies and pest control strategies [87] | |
| Spodoptera frugiperda | Sfabd-A | Indel | Defected body segmentation [21] |
| BLOS2E93 TO | Knockout | Developed mutants [88] | |
| SfABCC2 | Edit | Resistance to Cry1F toxin of Bacillus thuringiensis [52] | |
| ABC transporters | Toxicity of two Bacillus thuringiensis Cry1 toxins to the pest [53] | ||
| ABCB1 | Knockout | Susceptibility to emamectinbenzoate, beta-cypermethrin and chlorantraniliprole [54] | |
| Helicoverpa armigera | nAchR | Knockout | Resistance to insecticides [23] |
| a-6-nicotinic acetylcholine receptor (nAchR) | Knockout | Resistance to spinosyn [23,68,69] | |
| HaCad | Knockout | Resistance to Bttoxin Cry1Ac [37] | |
| Cluster of nine P450 genes | Knockout | Identification of the key players in the insecticide metabolism [39] | |
| CYP6AE | Knockout | Regulation of detoxification enzymes [39] | |
| OR16 | Knockout | Destroyed pest insect mating [41] | |
| Tetraspanin | Knockout | Resistance to Bt toxin cry1Ac [89] | |
| HaABCA2 | Knockout | Resistance to cry2Aa and cry2Ab [55] | |
| White, ok, brown, and scarlet | Knockout | Differential distribution of eye pigments that are helpful in elucidation of biosynthetic pathway [90] | |
| NPC1b | Knockout | NPC1b wasvital for growth and dietary cholesterol uptake [91] | |
| Helicoverpapunctigera | HaABCA2 | Deletion | Resistance to both cry2Aa and cry2Ab [55] |
| Plutellaxy lostella | Abdominal-A | Knockout | Defected body segmentation [43] |
| Pxabd-A | Knockout | Providing novel ideas for pest management [43]. | |
| PxCHS1 | Knockout | Described the resistance management strategies of major agricultural pests [45] | |
| PxABCC2 PxABCC3 | Knockout | Resistance to cry1 Ac protoxin [92] | |
| nAChRα6 | Knockout | Resistance to spinosad [56] | |
| LW | Knockout | Weaker phototaxis and reduced locomotion [93] | |
| Pxdsx | Knockout | Altered expression of sex-biased genes [94] | |
| Dendrolimus punctatus | DpWnt-1 | Knockout | Defected anterior segmentation and appendage development [57] |
| Bemisia tabaci | White | Edit | Altered eye pigmentation [58] |
| Nilaparvata lugens | Cinnabar and white | Edit | Altered eye pigmentation [59] |
| Nl-cn and Nl-w | Knockout | Paved a path for gene-function interrogation [59] | |
| Ceratitis capitata | White eye (we) and paired gene (Ccprd) | Knockout | Embryonic developmental malformations [60] |
| eGFP_gRNA2, eGFP_gRNA2, 1 mM Scr7, and eGFP_gRNA2b–Cas9 complexes with ssODN_BFP donor templates | Knock-In | Conversion of eGFP to BFP [95] | |
| Bactrocera dorsalis | White and transformer | Knockout | Various phenotypic effects [96] |
| Anastrepha ludens | Astra-2 | Knockout | The mutation caused sterility [97] |
| Locust amigratoria | Orco | Knockout | Generated lossoffunction for the management of insect pests [36] |
| OfAgo1 | Knockout | Cuticle disruption [98] | |
| Cydia pomonella | CpomOR1 | Knockout, Knock-In | Affected egg production and viability [35] |
| Tetranychus urticae | Phytoene desaturase | Knockout | Functional studies [99] |
| PSST | Knockout | Pyridaben resistance [100] | |
| Leptinotarsa decemlineata | Vestigial gene (vest) | Knockout | Functionally characterized vest gene and mutagenesis [101] |
| Euschistus heros | Abnormal wing disc (awd), tyrosine hydroxylase (th), and yellow (yel) | Knockdown and knockout | Managing insect pests [102] |
| Diaphorina Citri, Homalodisca vitripennis, and Bemisia argentifolii | Thioredoxin and vermillion | Knockout | Protected food crops from different pathogens and insect vectors [103] |
| Diaphorina citri | ACP-TRX-2 | Knockout | Innovative breakthrough in gene editing [104] |
| Mythimna separata | NPC1b | Knockout | Hampered nutrient absorption [105] |
| Hyphantriacunea | Hcdsx | Knockout | Sex-specific sterility [106] |
| Agrotis ipsilon | AiTH | Knockout | Narrowing in the eggshell [107] |
| Danaus plexippus | Clk | Knockout | Defined the role of the clk gene in the control of migration behavior [108] |
| Bombyx mori | BmBLOS2 | Knockout | Generated designer flies [34] |
| BmOrco | Knockout | Impaired olfactory sensitivity [109] | |
| Tribolium castaneum | EGFP | Knockout, Knock-In | Controlled insect pests and created resistance to insecticides [110] |
| Tribolium E-cadherin | Knockout | Knockout study [110] | |
| Gryllus bimaculatus | Dop1 | Knockout | Destroyed appetitive reinforcement [111] |
| Rhopalosiphum padi | ß-1-3glucanase | Knockout | Reduced callose deposition in maize [112] |
| Ostrinia furnacalis | ABCC2 | Knockout | Resistance to Btcry1Fa toxin [113,114] |
3.2. CRISPR-Cas-Mediated Gene Drive in Insect Pest Management
3.3. CRISPR-Cas Technology in Genome Editing of Crop Plants
| Name of Crops | Target Gene | Editing | Outcome |
|---|---|---|---|
| Barley | Beta-1-3 glucanase | Alteration | Resistance to aphid infestation [112] |
| Paulownia tomentosa | PtoPDS1 and PtoPDS2 | Deletion | Enhanced endogenous defenses and increased resistance to insects [121,122]. |
| Tobacco | Anthocyanin pathway | Alteration | Discouraged insect attack [123] |
| Soybean | GmCDPK38 | Knockout | Resistance to common cutworm [124] |
| GmUGT | 1bp and 33bp deletion | Enhanced resistance to Helicoverpaarmigera and Spodopteralitura [125] | |
| Cry 8 like | Resistance to Coleopteran-Holtrichiapanallele [126] | ||
| Solanum pimpinellifolium | Six different genes | Editing | Resistance to insect pests [127]. |
| Rice | OsCYP71A1 | Deletion | Resistance to the striped stem borer and the brown plant hopper [22]. |
| Cry2A | Transgene | Resistance to leaf folder [128] | |
| Mannose-specific GNA | Transgene | Resistance toBPH (Nilaparvata lugens) and hemipteran pest [127] | |
| Cry1AC + ASAL | Transgene | Resistance to yellow stem borer, leaf folder, and BPH [129] | |
| Cry1Ab + vip3A | Transgene | Resistance toAsian stem borer and rice leaf folder [130] | |
| Cry2AX1 | Transgene | Resistance to rice leaf folder [131] | |
| Cry2Aa + cry1Ca | Transgene | Resistance to Chilo suppressalis [132] | |
| Cry2AX1 (cry2Aa + cry2Ac) | Transgene | Resistance to Lepidopteran pest [133] | |
| Cotton | Cry2AX1 | Transgene | Resistance to H. armigera [134] |
| Cry2Ab, cry1F, and cry1AC | Transgene | Resistance to Lepidopteran pest H. armigera, and S. litura [135] | |
| Cry1AC and cry2Ab | Transgene | Resistance to S. litura [136] | |
| Cry2AX | Transgene | Resistance to H. armigera [137] | |
| Cry1Aa | Transgene | Resistance to Anthamous grandis [138] | |
| Cry1Ab | Transgene | Resistance to Heliothis [139] | |
| Cry1Ab+ NptII | Transgene | Resistance to H. armigera [140] | |
| Vip3AcAaa (vip3Aa1 + vip3Ac1) | Transgene | Resistance to Lepidopteran [141] | |
| Vip3A + cry1Ab | Transgene | Resistance to Heliothis. zea and H. virescens [142] | |
| Insect gut-binding lectin from Sclerotium rolfsii | Transgene | Resistance to chewing and sucking pest [3] | |
| Cry51Aa2 | Transgene | Resistance to Lygus species [143] | |
| Cry1Be + cry1Fa | Transgene | Resistance to S. litura and O. nubialis [144] | |
| Maize | Cry1Ab/cry2Aj | Transgene | Resistance to S. exigua and Harmonia axyridis [145] |
| Mustard (Brassica juncea) | Lectin protease protein(lentil lectin-LL CPPI) | Transgene | Resistance toaphid [146] |
| Colocasia esculenta tuber agglutinin (CEA) + GNA | Transgene | Resistance tomustard aphid (Lipaphis erysimi) [147] | |
| Sugarcane | Cry2Aa+ cry1Ca Cry1Ab + cry1Ac | Transgene | Resistance to shoot borer [148] |
| Vip3A | Transgene | Resistance to sugarcane stem borer (Chilo infuscatellus) [149] | |
| Potato | Hv1a/GNA | Transgene | Resistance to peach potato aphids and grain aphids [150] |
| Galanthus nivalis agglutinin (GNA) | Transgene | Resistance to aphids [151] | |
| Wheat | Pinelliapedatisecta agglutinin (PPA) | Transgene | Resistance to aphids [152] |
| Cowpea | Arcl on APA locus of Phaeselous vulgaris | Transgene | Resistance to bruchids [153] |
| Vip3Ba1 | Transgene | Resistance to legume pod borer (Maruca vitrata) [154] | |
| Pigeon pea | Cry2Aa | Transgene | Resistance to H. armigera [155] |
| Cry2Aa | Transgene | Resistan topod borer—H. armigera [156] | |
| Cry1AC, cry2Aa | Transgene | Resistance to H. armigera [157] | |
| Chickpea | CryIIAa | Transgene | Resistance to pod borer [158] |
| Tomato | Cry1Ac | Transgene | Resistance to Tuta Absoluta—tomato leaf miner [159] |
| Remusatia vivipara (rvl 1) and Sclerotiumrolfsii(srl 1) | Transgene | Resistance to root knot nematode (Meloidogyne incognita) [160] | |
| Cry1Ab | Transgene | Resistance to T. absoluta [161] | |
| Castor | Cry1AC | Transgene | Resistance to Lepidoteran—Achaea Janata and S. litura [162] |
| Sweet potato | Cry1Aa | Transgene | Resistance to S. litura [163] |
3.4. Utilization of Crop Wild Relatives in Insect Resistance by CRISPR-Cas Technology
4. Limitations and Future Perspectives
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Douglas, A.E. Strategies for enhanced crop resistance to insect pests. Annu. Rev. Plant Boil. 2018, 69, 637–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Global Agriculture Towards 2050. In High Level Expert Forum; FAO: Rome, Italy, 2009. [Google Scholar]
- Vanti, G.L.; Katageri, I.S.; Inamdar, S.R.; Hiremathada, V.; Swamy, B.M. Potent insect gut binding lectin from Sclerotiumrolfsii impart resistance to sucking and chewing type insects in cotton. J. Biotechnol. 2018, 278, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Birkett, M.A.; Pickett, J.A. Prospects of genetic engineering for robust insect resistance. Curr. Opin. Plant Biol. 2014, 19, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taning, C.N.T.; Eynde, B.V.; Yu, N.; Ma, S.; Smagghe, G. CRISPR/Cas9 in insects: Applications, best practices and biosafety concerns. J. Insect Physiol. 2017, 98, 245–257. [Google Scholar] [CrossRef]
- Islam, T. CRISPR-Cas technology in modifying food crops. CAB Rev. 2019, 14, 1–16. [Google Scholar] [CrossRef]
- Andolfo, G.; Iovieno, P.; Frusciante, L.; Ercolano, M.R. Genome-editing technologies for enhancing plant disease resistance. Front. Plant Sci. 2016, 7, 1813. [Google Scholar] [CrossRef] [Green Version]
- Bhowmik, P.K.; Islam, M.T. CRISPR-Cas9-Mediated Gene Editing in Wheat: A Step-by-Step Protocol. In CRISPR-Cas Methods, 1st ed.; Islam, M.T., Bhowmik, P.K., Molla, K.A., Eds.; Humana Press: New York, NY, USA, 2020; Volume 1, pp. 203–222. [Google Scholar]
- Maeder, M.L.; Gersbach, C.A. Genome-editing technologies for gene and cell therapy. Mol. Ther. 2016, 24, 430–446. [Google Scholar] [CrossRef] [Green Version]
- Gilles, A.F.; Averof, M. Functional genetics for all: Engineered nucleases, CRISPR and the gene editing revolution. EvoDevo 2014, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Baltes, N.J.; Voytas, D.F. Enabling plant synthetic biology through genome engineering. Trends Biotechnol. 2015, 33, 120–131. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Valarmathi, R.; Nandini, S.; Nandhakumar, M.R. Genetically modified crops: Insect resistance. Biotechnology 2012, 11, 119–126. [Google Scholar] [CrossRef]
- Gantz, V.M.; Akbari, O.S. Gene editing technologies and applications for insects. Curr. Opin. Insect Sci. 2018, 28, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPR Finder: A web tool to identify clustered regularly 7 interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Nobel Prize in Chemistry, NobelPrize.org. Nobel Media AB. 2020. Available online: https://www.nobelprize.org/prizes/chemistry/2020/summary/ (accessed on 15 November 2022).
- Devashish, R.; Lina, A.; Archana, R.; Magnus, L. The CRISPR-Cas immune system: Biology, mechanisms and applications. Biochimie 2015, 117, 119–128. [Google Scholar]
- Courtier-Orgogozo, V.; Morizot, B.; Boëte, C. Agricultural pest control with CRISPR-based gene drive: Time for public debate: Should we use gene drive for pest control? EMBO Rep. 2017, 18, 878–880. [Google Scholar] [CrossRef]
- Haque, E.; Taniguchi, H.; Hassan, M.M.; Bhowmik, P.; Karim, M.R.; Śmiech, M.; Zhao, K.; Rahman, M.; Islam, T. Application of CRISPR/Cas9 genome editing technology for the improvement of crops cultivated in tropical climates: Recent progress, prospects, and challenges. Front. Plant Sci. 2018, 9, 617. [Google Scholar] [CrossRef] [Green Version]
- Molla, K.A.; Karmakar, S.; Islam, M.T. Wide Horizons of CRISPR-Cas-derived technologies for basic biology, agriculture, and medicine. In CRISPR-Cas Methods, 1st ed.; Islam, M.T., Bhowmik, P.K., Molla, K.A., Eds.; Humana Press: New York, NY, USA, 2020; Volume 1, pp. 1–23. [Google Scholar]
- Wu, K.; Shirk, P.D.; Taylor, C.E.; Furlong, R.B.; Shirk, B.D.; Pinheiro, D.H.; Siegfried, B.D. CRISPR/Cas9 mediated knockout of the abdominal-A homeotic gene in fall armyworm moth (Spodopterafrugiperda). PLoS ONE 2018, 13, e0208647. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.P.; Luo, T.; Fu, H.W.; Wang, L.; Tan, Y.Y.; Huang, J.Z.; Wang, Q.; Ye, G.Y.; Gatehouse, A.M.R.; Lou, Y.G.; et al. Resistance of rice to insect pests mediated by suppression of serotonin biosynthesis. Nat. Plants. 2018, 4, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Xue, Y.; Lu, W.; Ma, H.; Chen, M.; Wu, Y.; Yang, Y.; Hu, Z. Functional validation of nicotinic acetylcholine receptor (nAChR) α6as a target of spinosyns in Spodopteraexigua utilizing the CRISPR/Cas9 system. Pest Manag. Sci. 2020, 76, 2415–2422. [Google Scholar] [CrossRef]
- Marraffini, L.A. The CRISPR-Cas system of Streptococcus pyogenes: Function and applications. In Streptococcus Pyogenes: Basic Biology to Clinical Manifestation, 1st ed.; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Yin, K.Q.; Gao, C.X.; Qiu, J.L. Progress and prospects in plant genome editing. Nat. Plants 2017, 3, 17107. [Google Scholar] [CrossRef]
- Reid, W.; O’Brochta, D.A. Applications of genome editing in insects. Curr. Opin. Insect Sci. 2016, 13, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Scott, M.J. CRISPR/Cas9-mediated mutagenesis of the white and Sex lethal loci in the invasive pest, Drosophila suzukii. Biochem. Biophys. Res. Commun. 2016, 469, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Guo, Z.; Liu, Y.; Zhang, Y. Progress and prospects of CRISPR/Cas systems in insects and other arthropods. Front. Physiol. 2017, 8, 608. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Dong, Y.; Liu, X.; Yao, G.; Yu, X.; Yang, M. The current status and development of insect-resistant genetically engineered poplar in China. Front. Plant Sci. 2018, 9, 1408. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, G.; Zhu, Y.-N.; Xiang, W.W.H. Advances and perspectives in the application of CRISPR/Cas9 in insects. Zool. Res. 2016, 37, 136. [Google Scholar]
- Bortesiand, L.; Fischer, R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef]
- Koutroumpa, F.A.; Monsempes, C.; Francois, M.C.; Cian, A.D.; Royer, C.; Concordet, J.P.; Jacquin-Joly, E. Heritable genome editing with CRISPR/Cas9 induces anosmia in a crop pest moth. Sci. Rep. 2016, 6, 29620. [Google Scholar] [CrossRef]
- Gratz, S.J.; Cummings, A.M.; Nguyen, J.N.; Hamm, D.C.; Donohue, L.K.; Harrison, M.M.; Wildonger, J.; O’Connor-Giles, K.M. Genome engineering of Drosophila with the CRISPR RNA-Guided Cas9 nuclease. Genetic 2013, 194, 1029–1035. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Z.; Xu, J.; Zeng, B.; Ling, L.; You, L.; Chen, Y.; Huang, Y.; Tan, A. The CRISPR/Cas System mediates efficient genome engineering in Bombyxmori. Cell Res. 2013, 23, 1414–1416. [Google Scholar] [CrossRef] [Green Version]
- Garczynski, S.F.; Martin, J.A.; Griset, M.; Willett, L.S.; Cooper, W.R.; Swisher, K.D.; Unruh, T.R. CRISPR/Cas9 editing of the codling moth (Lepidoptera: Tortricidae) CpomOR1 gene affects egg production and viability. J. Econ. Entomol. 2017, 110, 1847–1855. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Chen, D.; Yang, P.; Jiang, F.; Wang, X.; Kang, L. CRISPR/Cas9 in locusts: Successful establishment of an olfactory deficiency line by targeting the mutagenesis of an odorant receptor co-receptor (Orco). Insect Biochem. Mol. Biol. 2016, 79, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, H.; Wang, H.; Zhao, S.; Zuo, Y.; Yang, Y.; Wu, Y. Functional validation of cadherin as a receptor of Bt toxin Cry1Ac in Helicoverpaarmigera utilizing the CRISPR/Cas9 system. Insect Biochem. Mol. Biol. 2016, 76, 11–17. [Google Scholar] [CrossRef]
- Hafeez, M.; Liu, S.; Jan, S.; Shi, L.; Fernández-Grandon, G.M.; Gulzar, A.; Ali, B.; Rehman, M.; Wang, M. Knock-down of gossypolinducing cytochrome P450 genes reduced deltamethrin sensitivity in Spodopteraexigua (Huübner). Int. J. Mol. Sci. 2019, 20, 2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Shi, Y.; Wang, L.; Liu, S.; Wu, S.; Yang, Y.; Feyereisen, R.; Wu, Y. CYP6AE gene cluster knockout in Helicoverpaarmigera reveals role in detoxification of phytochemicals and insecticides. Nat. Commun. 2018, 9, 4820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, M.C.; Domingos, A.I.; Jones, W.D.; Chiappe, M.; Amrein, H.; Vosshall, L.B. Or83b encodes a broadly expressed odorant receptor essential for Drosophila olfaction. Neuron 2004, 43, 703–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.; Liu, Y.; Ai, D. A pheromone antagonist regulates optimal mating time in the moth Helicoverpaarmigera. Curr. Biol. 2017, 27, 1610–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, H.L.; Xu, J.; Tan, A.J.; Huang, Y.P. CRISPR/Cas9-mediated targeted gene mutagenesis in Spodopteralitura. Insect Sci. 2016, 23, 469–477. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Y.; Zeng, B. CRISPR/Cas9 mediated knockout of the abdominal-A homeotic gene in the global pest, diamondback moth (Plutellaxylostella). Insect Biochem. Mol.Biol. 2016, 75, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Asaoka, T.; Almagro, J.; Ehrhardt, C. Linear ubiquitination by LUBEL has a role in Drosophila heat stress response. EMBO Rep. 2016, 17, 1624–1640. [Google Scholar] [CrossRef]
- Douris, V.; Steinbach, D.; Panteleri, R. Resistance mutation conserved between insects and mites. Proc. Natl. Acad. Sci. USA 2016, 113, 14692–14697. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, C.T.; Garrood, W.T.; Puinean, A.M. A CRISPR/Cas9 mediated point mutation in the alpha 6 subunit of the nicotinic acetylcholine receptor confers resistance to spinosad in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2016, 73, 62–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, X.; Sato-Miyata, Y.; Tsuda, M. Deficiency of succinyl-CoA synthetase alpha subunit delays development, impairs locomotor activity and reduces survival under starvation in Drosophila. Biochem. Biophys. Res. Commun. 2017, 483, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Kaduskar, B.; Kushwah, R.B.S.; Auradkar, A. Reversing insecticide resistance with allelic-drive in Drosophila melanogaster. Nat. Commun. 2022, 13, 291. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Wang, H.; Xu, Y. CRISPR/Cas9 mediated G4946E substitution in the ryanodine receptor of Spodopteraexigua confers high levels of resistance to diamide insecticides. Insect Biochem. Mol. Biol. 2017, 89, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Shi, Y.; Zhang, F.; Guan, F.; Zhang, J.; Feyereisen, R.; Fabrick, J.A.; Yang, Y.; Wu, Y. Genome mapping coupled with CRISPR gene editing reveals a P450 gene confers avermectin resistance in the beet armyworm. PLoS Genet. 2021, 17, e1009680. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.Y.; Huang, J.L.; Wang, J.; Feng, Y.; Han, T.T.; Wu, Y.D.; Yang, Y.H. Knockout of a P-glycoprotein gene increases susceptibility to abamectin and emamectin benzoate in Spodopteraexigua. Insect Mol. Biol. 2018, 27, 36–45. [Google Scholar] [CrossRef]
- Jin, M.; Tao, J.; Li, Q.; Cheng, Y.; Sun, X.; Wu, K.; Xiao, Y. Genome editing of the SfABCC2 gene confers resistance to Cry1F toxin from Bacillus thuringiensis in Spodopterafrugiperda. J. Integr. Agric. 2021, 20, 815–820. [Google Scholar] [CrossRef]
- Jin, M.; Yang, Y.; Shan, Y.; Chakrabarty, S.; Cheng, Y.; Soberon, M.; Bravo, A.; Liu, K.; Wu, K.; Xiao, Y. Two ABC transporters are differentially involved in the toxicity of two Bacillus thuringiensis Cry1 toxins to the invasive crop-pest Spodopterafrugiperda (J. E. Smith). Pest Manag. Sci. 2020, 77, 6170. [Google Scholar] [CrossRef]
- Li, Q.; Jin, M.; Yu, S.; Cheng, Y.; Shan, Y.; Wang, P.; Yuan, H.; Xiao, Y. Knockout of the ABCB1 Gene Increases Susceptibility to Emamectin Benzoate, Beta-Cypermethrin and Chlorantraniliprole in Spodopterafrugiperda. Insects 2022, 13, 137. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.; Liu, S. CRISPR/Cas9 mediated genome editing of Helicoverpaarmigera with mutations of an ABC transporter gene HaABCA2 confers resistance to Bacillus thuringiensis Cry2A toxins. Insect Biochem. Mol. Biol. 2017, 87, 147–153. [Google Scholar] [CrossRef]
- Wang, X.; Ma, Y.; Wang, F.; Yang, Y.; Wu, S.; Wu, Y. Disruption of nicotinic acetylcholine receptor α6 mediated by CRISPR/Cas9 confers resistance to spinosyns in Plutellaxylostella. Pest Manag. Sci. 2020, 76, 1618–1625. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, Q.; Zhou, X. Genome editing of Wnt-1, a gene associated with segmentation, via CRISPR/Cas9 in the pine caterpillar moth, Dendrolimuspunctatus. Front. Physiol. 2017, 7, 666. [Google Scholar] [CrossRef] [PubMed]
- Heu, C.C.; McCullough, F.M.; Luan, J.; Rasgon, J.L. CRISPR-Cas9-based genome editing in the silverleaf W\whitefly (Bemisiatabaci). Cris. J. 2020, 3, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, W.H.; Xu, N.; Yuan, X.B.; Chen, H.H.; Zhang, J.L.; Fu, S.J.; Zhang, C.X.; Xu, H.J. CRISPR/Cas9-mediated knockout of two eye pigmentation genes in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Biochem. Mol. Biol. 2018, 93, 19–26. [Google Scholar] [CrossRef]
- Meccariello, A.; Monti, S.M.; Romanelli, A.; Colonna, R.; Primo, P.; Inghilterra, M.G.; Corsano, G.D.; Ramaglia, A.; Iazzetti, G.; Chiarore, A.; et al. Highly efficient DNA-free gene disruption in the agricultural pest Ceratitis capitata by CRISPR-Cas9 ribonucleoprotein complexes. Sci. Rep. 2017, 7, 10061. [Google Scholar] [CrossRef] [Green Version]
- Kondo, S.; Ueda, R. Highly improved gene targeting by germline-specific Cas9 expression in Drosophila. Genetics 2013, 195, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Ren, M.; Wang, Z.; Zhang, B.; Rong, Y.S.; Jiao, R.; Gao, G. Highly efficient genome modifications mediated by CRISPR/Cas9 in Drosophila. Genetics 2013, 195, 289–291. [Google Scholar] [CrossRef] [Green Version]
- Bassett, A.R.; Tibbit, C.; Ponting, C.P.; Liu, J.L. Highly efficient targeted mutagenesis of Drosophila with the CRISPR/Cas9 system. Cell Rep. 2013, 4, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Gratz, S.J.; Wildonger, J.; Harrison, M.M.; O’Connor-Giles, K.M. CRISPR/Cas9-mediated genome engineering and the promise of designer flies on demand. Fly 2013, 7, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Mendoza-Garcia, P.; Hugosson, F.; Fallah, M.; Higgins, M.L.; Iwasaki, Y.; Pfeifer, K.; Wolfstetter, G.; Varshney, G.; Popichenko, D.; Gergen, J.P.; et al. The Zic family homologue Odd-paired regulates Alk expression in Drosophila. PLoS Genet 2017, 13, e1006617. [Google Scholar] [CrossRef] [Green Version]
- Chechenova, M.B.; Maes, S.; Oas, S.T.; Nelson, C.; Kiani, K.G.; Bryantsev, A.L. Functional redundancy and non-redundancy between two Troponin C isoforms in Drosophila adult muscles. Mol. Biol. Cell. 2017, 28, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Port, F.; Bullock, S.L. Augmenting CRISPR applications in Drosophila with tRNA-flanked sgRNAs. Nat. Methods 2016, 13, 852–854. [Google Scholar] [CrossRef]
- Lamb, A.M.; Walker, E.A.; Wittkopp, P.J. Tools and strategies for scarless allele replacement in Drosophila using CRISPR/Cas9. Fly 2016, 11, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunzelmann, S.; Böttcher, R.; Schmidts, I.; Förstemann, K. A comprehensive toolbox for genome editing in cultured Drosophila melanogaster cells. G3 (Bethesda) 2016, 6, 1777–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.L.; Fan, Y.J.; Wang, X.Y.; Zhang, Y.; Pu, J.; Li, L.; Xu, Y.Z. A conserved intronic U1 snRNP-binding sequence promotes trans-splicing in Drosophila. Genes Dev. 2015, 29, 760–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mabashi-Asazuma, H.; Sohn, B.H.; Kim, Y.S.; Kuo, C.W.; Khoo, K.H.; Kucharski, C.A.; Jarvis, D.L. Targeted glycoengineering extends the protein N-glycosylation pathway in the silkworm silk gland. Insect Biochem. Mol. Biol. 2015, 65, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Chen, H.; Liu, J.; Zhang, H.; Yan, Y.; Zhu, N.; Guo, Y.; Yang, B.; Chang, Y.; Dai, F.; et al. Various applications of TALEN- and CRISPR/Cas9-mediated homologous recombination to modify the Drosophila genome. Biol. Open 2014, 3, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.A.; Doherty, C.A.; Jordan, W.T., III; Bliss, J.E.; Feng, J.; Soruco, M.M.; Rieder, L.E.; Tsiarli, M.A.; Larschan, E.N. The essential Drosophila CLAMP protein differentially regulates non-coding roX RNAs in male and females. Chromosome Res. 2016, 25, 101–113. [Google Scholar] [CrossRef]
- Gantz, V.M.; Bier, E. The mutagenic chain reaction: A method for converting heterozygous to homozygous mutations. Science 2015, 348, 442–444. [Google Scholar] [CrossRef] [Green Version]
- Port, F.; Muschalik, N.; Bullock, S.L. Systematic evaluation of Drosophila CRISPR tools reveals safe and robust alternatives to autonomous gene drives in basic research. G3 2015, 5, 1493–1502. [Google Scholar] [CrossRef] [Green Version]
- Gokcezade, J.; Sienski, G.; Duchek, P. Efficient CRISPR/Cas9 plasmids for rapid and versatile genome editing in Drosophila. G3 2014, 4, 2279–2282. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Yang, Z.; Mao, D.; Chang, Z.; Qiao, H.H.; Wang, X.; Sun, J.; Hu, Q.; Cui, Y.; Liu, L.P.; et al. Performance of the Cas9 nickase system in Drosophila melanogaster. G3 2014, 4, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Koolhaas, W.H.; Schnorrer, F. A versatile two-step CRISPR-and RMCE-based strategy for efficient genome engineering in Drosophila. G3 Genes Genomes Genet 2014, 4, 2409–2418. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Wu, M.; Wen, K.; Ren, M.; Long, L.; Zhang, X.; Gao, G. CRISPR/Cas9 mediates efficient conditional mutagenesis in Drosophila. G3 2014, 4, 2167–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Z.; Ren, M.D.; Wu, M.H.; Dai, J.B.; Rong, Y.K.S.; Gao, G.J. Efficient gene knock-out and knock-in with transgenic Cas9 in Drosophila. G3 2014, 4, 925–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebo, Z.L.; Lee, H.B.; Peng, Y.; Guo, Y. A simplified and efficient germline-specific CRISPR/Cas9 system for Drosophila genomic engineering. Fly 2014, 8, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Port, F.; Chen, H.M.; Lee, T.; Bullock, S.L. Optimized CRISPR/Cas tools for efficient germline and somatic genome engineering in Drosophila. Proc. Natl. Acad. Sci. USA 2014, 111, E2967–E2976. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, R.; Murakami, H.; Ote, M.; Yamamoto, D. Clustered regulatory interspaced short palindromic repeats (CRISPR)-mediated mutagenesis and phenotype rescue by piggyBac transgenesis in a non-model Drosophila species. Insect Mol. Biol. 2016, 25, 355–361. [Google Scholar] [CrossRef]
- Ahmed, H.M.; Hildebrand, L.; Wimmer, E.A. Improvement and use of CRISPR/Cas9 to engineer a sperm-marking strain for the invasive fruit pest Drosophila suzukii. BMC Biotechnol. 2019, 19, 85. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Ziemek, J.; Schetelig, M.F. CRISPR/Cas9 mediated disruption of the white gene leads to pigmentation deficiency and copulation failure in Drosophila suzukii. J. Insect Physiol. 2020, 126, 104091. [Google Scholar] [CrossRef]
- Zhu, G.H.; Xu, J.; Cui, Z. Functional characterization of SlitPBP3 in Spodopteralitura by CRISPR/Cas9 mediated genome editing. Insect Biochem. Mol. Biol. 2017, 75, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.H.; Peng, Y.C.; Zheng, M.Y.; Zhang, X.Q.; Sun, J.B.; Huang, Y.; Dong, S.L. CRISPR/Cas9 mediated BLOS2 knockout resulting in disappearance of yellow strips and white spots on the larval integument in Spodopteralitura. J. Insect Physiol. 2017, 103, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.H.; Chereddy, S.C.; Howell, J.L.; Palli, S.R. Genome editing in the fall armyworm, Spodopterafrugiperda: Multiple sgRNA/Cas9 method for identification of knockouts in one generation. Insect Biochem. Mol. Biol. 2020, 122, 103373. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Wang, J.; Guan, F. Dominant point mutation in a tetraspainin gene associated with field-evolved resistance of cotton bollworm to transgenic Bt cotton. Proc. Natl. Acad. Sci. USA 2018, 115, 11760–11765. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Reichelt, M.; Heckel, D.G. Functional analysis of the ABCs of eye color in Helicoverpaarmigera with CRISPR/Cas9-induced mutations. Sci. Rep. 2017, 7, 40025. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.C.; Yue, X.R.; Kuang, W.Q.; Li, S.L.; Tang, R.; Zhang, Z.F.; Jing, X. NPC1b as a novel target in controlling the cotton bollworm, Helicoverpaarmigera. Pest Manag. Sci. 2020, 76, 2233–2242. [Google Scholar] [CrossRef]
- Guo, Z.; Sun, D.; Kang, S. CRISPR/Cas9-mediated knockout of both the PxABCC2 and PxABCC3 genes confers high-level resistance to Bacillus thuringiensis Cry1Ac toxin in the diamondback moth, Plutellaxylostella (L.). Insect Biochem. Mol. Biol. 2019, 107, 31–38. [Google Scholar] [CrossRef]
- Chen, E.H.; Hou, Q.L. Identification and expression analysis of cuticular protein genes in the diamondback moth, Plutellaxylostella (Lepidoptera: Plutellidae). Pestic. Biochem. Physiol. 2021, 178, 104943. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Z.; Xu, J.; Li, X.; Bi, H.; Andongma, A.A.; Huang, Y. Mutation of double sex induces sex-specific sterility of the diamondback moth Plutellaxylostella. Insect Biochem. Mol. Biol. 2019, 112, 103180. [Google Scholar] [CrossRef]
- Aumann, R.A.; Schetelig, M.F.; Häcker, I. Highly efficient genome editing by homology-directed repair using Cas9 protein in Ceratitis capitata. Insect Mol. Biol. 2018, 101, 85–93. [Google Scholar] [CrossRef]
- Zhao, S.; Xing, Z.; Liu, Z.; Liu, Y.; Liu, X.; Chen, Z.; Yan, R. Efficient somatic and germline genome engineering of Bactroceradorsalis by the CRISPR/Cas9 system. Pest Manag. Sci. 2019, 75, 1921–1932. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Handler, A.M. CRISPR/Cas9-mediated gene editing in an exogenous transgene and an endogenous sex determination gene in the Caribbean fruit fly, Anastrephasuspensa. Gene 2019, 691, 160–166. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Bi, H.L.; Wang, Y.H.; Li, X.W.; Chen, X.E.; Li, Z.Q. CRISPR/Cas9-based mutation reveals Argonaute 1 is essential for pigmentation in Ostriniafurnacalis. Insect Sci. 2019, 26, 1020–1028. [Google Scholar] [CrossRef]
- Dermauw, W.; Jonckheere, W.; Riga, M.; Livadaras, I.; Vontas, J.; Van Leeuwen, T. Targeted mutagenesis using CRISPR-Cas9 in the chelicerate herbivore Tetranychusurticae. Insect Biochem. Mol. Bio. 2020, 120, 103347. [Google Scholar] [CrossRef] [PubMed]
- Bajda, S.; Dermauw, W.; Panteleri, R.; Sugimoto, N.; Douris, V.; Tirry, L. A mutation in the PSST homologue of complex I (NADH: Ubiquinone oxidoreductase) from Tetranychusurticae is associated with resistance to METI acaricides. Insect Biochem. Mol.Biol. 2017, 80, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Gui, S.; Taning, C.N.T.; Wei, D.; Smagghe, G. First report on CRISPR/Cas9-targeted mutagenesis in the Colorado potato beetle, Leptinotarsadecemlineata. J. Insect Physiol. 2020, 121, 104013. [Google Scholar] [CrossRef]
- Cagliari, D.; Smagghe, G.; Zotti, M.; Taning, C.N.T. RNAi and CRISPR/Cas9 as Functional Genomics Tools in the Neotropical Stink Bug, Euschistusheros. Insects 2020, 11, 838. [Google Scholar] [CrossRef]
- Hunter, W.B.; Gonzalez, M.T.; Tomich, J. BAPC-assisted-CRISPR-Cas9 Delivery into Nymphs and Adults for Heritable Gene Editing (Hemiptera). FASEB J. 2019, 33 (Suppl. S1), 626.2. [Google Scholar] [CrossRef]
- Hunter, W.B.; Gonzalez, M.T.; Tomich, J. BAPC-assisted CRISPR/Cas9 System: Targeted Delivery into Adult Ovaries for Heritable Germline Gene Editing (Arthropoda: Hemiptera). BioRxiv 2018, 478743. [Google Scholar]
- Tang, R.; Li, S.; Liang, J.; Yi, H.; Jing, X.; Liu, T.X. Optimization of the application of the CRISPR/Cas9 system in Mythimnaseparata. Entomol. Exp. Appl. 2022, 170, 593–602. [Google Scholar] [CrossRef]
- Li, X.; Liu, Q.; Liu, H.; Bi, H.; Wang, Y.; Chen, X.; Chen, H. Mutation of double sex in Hyphantriacunea results in sex-specific sterility. Pest Manag. Sci. 2020, 76, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, Y.H.; Chen, X.E.; Tian, D.; Xu, X.; Li, K.; He, L. CRISPR/Cas9-mediated Tyrosine hydroxylase knockout resulting in larval lethality in Agrotisipsilon. Insect Sci. 2018, 25, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Markert, M.J.; Zhang, Y.; Enuameh, M.S.; Reppert, S.M.; Wolfe, S.A.; Merlin, C. Genomic access to monarch migration using TALEN and CRISPR/Cas9-mediated targeted mutagenesis. G3 2016, 6, 905–915. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, W.; Zeng, B. Deletion of the Bombyx mori odorant receptor co-receptor (BmOrco) impairs olfactory sensitivity in silkworms. Insect Biochem. Mol. Biol. 2017, 86, 58–67. [Google Scholar] [CrossRef]
- Gilles, A.F.; Schinko, J.B.; Averof, M. Efficient CRISPR-mediated gene targeting and transgene replacement in the beetle Triboliumcastaneum. Development 2015, 142, 2832–2839. [Google Scholar] [PubMed] [Green Version]
- Awata, H.; Watanabe, T.; Hamanaka, Y. Knockout crickets for the study of learning and memory: Dopamine receptor Dop1 mediates aversive but not appetitive reinforcement in crickets. Sci. Rep. 2015, 5, 15885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Bengtsson, T.; Olsson, N. Mutations in two aphid-regulated b-1, 3-glucanase genes by CRISPR/Cas9 do not increase barley resistance to Rhopalosiphumpadi L. Front. PlantSci. 2020, 11, 1043. [Google Scholar] [CrossRef]
- Wang, J.; Ma, H.; Zuo, Y.; Yang, Y.; Wu, Y. CRISPR-mediated gene knockout reveals nicotinic acetylcholine receptor (nAChR) subunit a6 as a target of spinosyns in Helicoverpaarmigera. Pest Manag. Sci. 2020, 76, 2925–2931. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Y.; Huang, J. CRISPR-mediated knockout of the ABCC2 gene in Ostriniafurnacalis confers high-level resistance to the Bacillus thuringiensis Cry1Fa toxin. Toxins 2020, 12, 246. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, G.R.; Whitelaw, C.B.A.; Lillico, S.G. CRISPR-based gene drives for pest control. Trends Biotechnol. 2018, 36, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Books, A. New Biotechnological Approaches to Insect Pest Management and Crop Protection; Gene Editing Approach (CRISPR-Cas System). Master’s Thesis, MS Project, Distance Master of Science in Entomology Projects, Department of Entomology, University of Nebraska, Lincoln, NE, USA, 2019; p. 44. [Google Scholar]
- Kolmer, J.A.; Bernardo, A.; Bai, G.; Hayden, M.J.; Chao, S. Adult plant leaf rust resistance derived from toropi wheat is conditioned by Lr78 and three minor QTL. Phytopathology 2018, 108, 246–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akumo, D.N.; Riedel, H.; Semtanska, I. Social and economic issues-genetically modified food. In Food Industry, InnocenzoMuzzalupo; IntechOpen: London, UK, 2013. [Google Scholar]
- Beale, M.H.; Birkett, M.A.; Bruce, T.J. Aphid alarm pheromone produced by transgenic plants affects aphid and parasitoid behavior. Proc. Natl. Acad. Sci. USA 2006, 103, 10509–10513.00FC. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathinam, M.; Mishra, P.; Mahato, A.K.; Singh, N.K.; Rao, U.; Sreevathsa, R. Comparative transcriptome analyses provide novel insights into the differential response of Pigeonpea (Cajanuscajan L.) and its wild relative (Cajanusplatycarpus (Benth.) Maesen) to herbivory by Helicoverpaarmigera (Hübner). Plant Mol. Biol. 2019, 101, 163–182. [Google Scholar] [CrossRef]
- Fan, D.; Liu, T.T.; Li, C.F. Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 2015, 5, 12217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.T.; Fan, D.; Ran, L.Y.; Jiang, Y.Z.; Liu, R. Highly efficient CRISPR/Cas9-mediated targeted mutagenesis of multiple genes in populus. Hereditas 2015, 37, 1044–1052. [Google Scholar]
- Malone, L.A.; Barraclough, E.I.; Lin-Wang, K.; Stevenson, D.E.; Allan, A.C. Effects of red-leaved transgenic tobacco expressing a MYB transcription factor on two herbivorous insects, Spodopteralitura and Helicoverpaarmigera. Entomol. Exp. Appl. 2009, 133, 117–127. [Google Scholar] [CrossRef]
- Li, X.; Hu, D.; Cai, L.; Wang, H.; Liu, X.; Du, H.; Yang, Z.; Zhang, H.; Hu, Z.; Huang, F.; et al. CALCIUM DEPENDENT PROTEIN KINASE38 regulates flowering time and common cutworm resistance in soybean. Plant Physiol. 2022, 190, 480–499. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, W.; Chen, L.; Shen, X.; Yang, H.; Fang, Y.; Ouyang, W.; Mai, S.; Chen, H.; Chen, S.; et al. CRISPR/Cas9-mediated targeted mutagenesis of GmUGT enhanced soybean resistance against leaf-chewing insects through flavonoids biosynthesis. Front. Plant Sci. 2022, 13, 802716. [Google Scholar] [CrossRef]
- Qin, D.; Liu, X.Y.; Miceli, C.; Zhang, Q.; Wang, P.W. Soybean plants expressing the Bacillus thuringiensis cry8-like gene show resistance to Holotrichiaparallela. BMC Biotechnol. 2019, 19, 66. [Google Scholar] [CrossRef] [Green Version]
- Bharathi, Y.; Kumar, S.V.; Pasalu, I.C.; Balachandran, S.M.; Reddy, V.D.; Rao, K.V. Pyramided rice lines harbouring Allium sativum(asal) and Galanthusnivalis(gna) lectin genes impart enhanced resistance against major sap sucking pests. J. Biotechnol. 2011, 152, 63–71. [Google Scholar] [CrossRef]
- Gunasekara, J.M.; Jayasekera, G.A.; Perera, K.L.; Wickramasuriya, A.M. Development of a Sri Lankan rice variety Bg 94-1 harbouringCry2A gene of Bacillus thuringiensis resistant to rice leaf folder [Cnaphalocrocismedinalis (Guenée)]. J. Natl. Sci. Found Sri Lanka. 2017, 45, 2. [Google Scholar]
- Boddupally, D.; Tamirisa, S.; Gundra, S.R.; Vudem, D.R.; Khareedu, V.R. Expression of hybrid fusion protein (Cry1Ac: ASAL) in transgenic rice plants imparts resistance against multiple insect pests. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Cheng, J.; Lin, H.; Lin, C.; Gao, J.; Shen, Z. Characterization of transgenic rice expressing fusion protein Cry1Ab/Vip3A for insect resistance. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rajadurai, G.; Kalaivani, A.; Varanavasiyappan, S.; Balakrishnan, N.; Udayasuriyan, V.; Sudhakar, D.; Natarajan, N. Generation of insect resistant marker-free transgenic rice with a novel cry2AX1 gene. Ele. J. Plant Breed. 2018, 9, 723–732. [Google Scholar] [CrossRef]
- Qiu, L.; Sun, Y.; Jiang, Z.; Yang, P.; Liu, H.; Zhou, H.; Wang, X.; Zhang, W.; Lin, Y.; Ma, W. The midgut V-ATPase subunit A gene is associated with toxicity to crystal 2Aa and crystal 1Ca-expressing transgenic rice in Chilo suppressalis. Insect Mol. Biol. 2019, 28, 520–527. [Google Scholar] [CrossRef]
- Chakraborty, M.; Reddy, P.S.; Mustafa, G.; Rajesh, G.; Narasu, V.L.; Udayasuriyan, V.; Rana, D. Transgenic rice expressing the cry2AX1 gene confers resistance to multiple lepidopteran pests. Transgenic Res. 2016, 25, 665–678. [Google Scholar] [CrossRef]
- Jadhav, M.S.; Rathnasamy, S.A.; Natarajan, B.; Duraialagaraja, S.; Varatharajalu, U. Study of expression of indigenous Bt cry2AX1 gene in T3 progeny of cotton and its efficacy against Helicoverpaarmigera (Hubner). Braz. Arch. Biol. Technol. 2020, 63, e20180428. [Google Scholar] [CrossRef]
- Katta, S.; Talakayala, A.; Reddy, M.K.; Addepally, U.; Garladinne, M. Development of transgenic cotton (Narasimha) using triple gene cry2Ab-cry1F-cry1Ac construct conferring resistance to lepidopteran pests. J. Biosci. 2020, 45, 31. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, H.A.; Asif, M.; Asad, S.; Naqvi, R.Z.; Ajaz, S.; Umer, N.; Anjum, N.; Rauf, I.; Sarwar, M.; Arshad, M.; et al. Development and evaluation of double gene transgeniccotton lines expressing Cry toxins for protection against chewing insect pests. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sakthi, A.R.; Naveenkumar, A.; Deepikha, P.S.; Balakrishnan, N.; Kumar, K.K.; Devi, E.K.; Balasubramani, V.; Arul, L.; Singh, P.K.; Sudhakar, D.; et al. Expression and inheritance of chimeric cry2AX1 gene in transgenic cotton plants generated through somatic embryogenesis. In Vitro Cell Dev. Biol. Plant 2015, 51, 379–389. [Google Scholar] [CrossRef]
- Ribeiro, T.P.; Arraes, F.B.; Lourenc¸o-Tessutti, I.T.; Silva, M.S.; Lisei-de-Sa, M.E.; Lucena, W.A.; Macedo, L.L.; Lima, J.N.; Santos Amorim, R.M.; Artico, S.; et al. Transgenic cotton expressing Cry10Aa toxin confers high resistance to the cotton boll weevil. Plant Biotechnol. J. 2017, 15, 997–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, G.A.; Bakhsh, A.; Ghazanfar, M.; Riazuddin, S.; Husnain, T. Development of transgenic cotton lines harboring a pesticidal gene (Cry1Ab). Emirates J. Food Agric. 2013, 25, 434–442. [Google Scholar] [CrossRef]
- Khan, G.A.; Bakhsha, A.; Riazuddin, S.; Husnain, T. Introduction of Cry1Ab gene into cotton (Gossypiumhirsutum) enhances resistance against Lepidopteran pest (Helicoverpaarmigera). Spanish J. Agric. Res. 2011, 1, 296–302. [Google Scholar] [CrossRef]
- Chen, W.; Liu, C.; Lu, G.; Cheng, H.; Shen, Z.; Wu, K. Effects of Vip3AcAa? Cry1Ac cotton on midgut tissue in Helicoverpaarmigera (Lepidoptera: Noctuidae). J. InsectSci. 2018, 18, 13. [Google Scholar]
- Bommireddy, P.L.; Leonard, B.R.; Temple, J.; Price, P.; Emfinger, K.; Cook, D.; Hardke, J.T. Field performance and seasonal efficacy profiles of transgenic cotton lines expressing Vip3A and VipCot against Helicoverpazea (Boddie) and Heliothisvirescens (F.). J. Cotton Sci. 2011, 15, 251–259. [Google Scholar]
- Gowda, A.; Rydel, T.J.; Wollacott, A.M.; Brown, R.S.; Akbar, W.; Clark, T.L.; Flasinski, S.; Nageotte, J.R.; Read, A.C.; Shi, X.; et al. A transgenic approach for controlling Lygus in cotton. Nat. Commun. 2016, 7, 12213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meade, T.; Narva, K.; Storer, N.P.; Sheets, J.J.; Burton, S.L.; Woosley, A.T. Inventors, and Dow AgroSciences LLC, assignee 2017. Insect resistance management with combinations of Cry1Be and Cry1F proteins. U.S. Patent 9,453,556, 28 February 2018. [Google Scholar]
- Chang, X.; Lu, Z.; Shen, Z.; Peng, Y.; Ye, G. Bitrophic and tritrophic effects of transgenic cry1Ab/cry2Aj maize on the beneficial, nontarget Harmoniaaxyridis (Coleoptera: Coccinellidae). Environ. Entomol. 2017, 46, 1171–1176. [Google Scholar] [CrossRef]
- Rani, S.; Sharma, V.; Hada, A.; Bhattacharya, R.C.; Koundal, K.R. Fusion gene construct preparation with lectin and protease inhibitor genes against aphids and efficient genetic transformation of Brassica juncea using cotyledon explants. Acta Physiol. Plant. 2017, 39, 115. [Google Scholar] [CrossRef]
- Das, A.; Ghosh, P.; Das, S. Expression of Colocasia esculentatuber agglutinin in Indian mustard providesresistance against Lipaphiserysimi and the expressed protein is non-allergenic. Plant Cell Rep. 2018, 37, 849–863. [Google Scholar] [CrossRef]
- Koerniati, S.; Sukmadjaja, D.; Samudra, I.M. C synthetic gene of CryIAb-CryIAc fusion to generate resistant sugarcane to shoot or stem borer. InIOP Conf. Ser. Earth Environ. Sci. 2020, 418, 012069. [Google Scholar] [CrossRef]
- Riaz, S.; Nasir, I.A.; Bhatti, M.U.; Adeyinka, O.S.; Toufiq, N.; Yousaf, I.; Tabassum, B. Resistance to Chiloinfuscatellus(Lepidoptera: Pyraloidea) in transgenic lines of sugarcane expressing Bacillus thuringiensis derived Vip3A protein. Mol. Biol. Rep. 2020, 47, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nakasu, E.Y.; Edwards, M.G.; Fitches, E.; Gatehouse, J.A.; Gatehouse, A.M. Transgenic plants expressing x- ACTX-Hv1a and snowdrop lectin (GNA) fusion protein show enhanced resistance to aphids. Front. Plant Sci. 2014, 28, 673. [Google Scholar] [CrossRef] [Green Version]
- Mi, X.; Liu, X.; Yan, H.; Liang, L.; Zhou, X.; Yang, J.; Si, H.; Zhang, N. Expression of the Galanthus nivalis agglutinin (GNA) gene in transgenic potato plants confers resistance to aphids. ComptesRendusBiol. 2017, 340, 7–12. [Google Scholar]
- Duan, X.; Hou, Q.; Liu, G.; Pang, X.; Niu, Z.; Wang, X.; Zhang, Y.; Li, B.; Liang, R. Expression of Pinelliapedatisectalectin gene in transgenic wheat enhances resistance to wheat aphids. Molecules 2018, 23, 748. [Google Scholar] [CrossRef] [Green Version]
- Grazziotin, M.A.; Cabral, G.B.; Ibrahim, A.B.; Machado, R.B.; Aragão, F.J. Expression of the Arcelin 1 gene from Phaseolus vulgaris L. in cowpea seeds (Vigna unguiculata L.) confers bruchid resistance. Ann. Appl. Biol. 2020, 176, 268–274. [Google Scholar] [CrossRef]
- Bett, B.; Gollasch, S.; Moore, A.; James, W.; Armstrong, J.; Walsh, T.; Harding, R.; Higgins, T.J. Transgenic cowpeas (Vigna unguiculata L. Walp) expressing Bacillusthuringiensis Vip3Ba protein are protected against the Maruca pod borer (Marucavitrata). Plant Cell Tiss. Org.Cult. 2017, 131, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Baburao, T.M.; Sumangala, B. Development and molecular characterization of transgenic Pigeon pea carrying cry2Aa for pod borer resistance. J. Pharm. Phytochem. 2018, 75, 1581–1585. [Google Scholar]
- Singh, S.; Kumar, N.R.; Maniraj, R.; Lakshmikanth, R.; Rao, K.Y.; Muralimohan, N.; Arulprakash, T.; Karthik, K.; Shashibhushan, N.B.; Vinutha, T.; et al. Expression of Cry2Aa, a Bacillus thuringiensis insecticidal protein in transgenic pigeon pea confers resistance to gram pod borer, Helicoverpaarmigera. Sci. Rep. 2018, 8, 8820. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, G.; Ganguly, S.; Purohit, A.; Chaudhuri, R.K.; Das, S.; Chakraborti, D. Transgenic pigeonpea events expressing Cry1Ac and Cry2Aa exhibit resistance to Helicoverpaarmigera. Plant Cell Rep. 2017, 36, 1037–1051. [Google Scholar] [CrossRef]
- Sawardekar, S.V.; Katageri, I.S.; Salimath, P.M.; Kumar, P.A.; Kelkar, V.G. Standardization of in-vitro genetictransformation technique in chickpea (Cicer arietinum L.) for pod-borer resistance. Adv. Agric. Res. Technol. J. 2017, 1, 2. [Google Scholar]
- Selale, H.; Dağlı, F.; Mutlu, N.; Doğanlar, S.; Frary, A. Cry1Ac-mediated resistance to tomato leaf miner (Tutaabsoluta) in tomato. Plant Cell Tiss. Org. Cult. 2017, 131, 65–73. [Google Scholar] [CrossRef]
- Bhagat, Y.S.; Bhat, R.S.; Kolekar, R.M.; Patil, A.C.; Lingaraju, S.; Patil, R.V.; Udikeri, S.S. Remusatia viviparalectin and Sclerotiumrolfsiilectin interfere with the development and gall formation activity of Meloidogyne incognita in transgenic tomato. Transgenic Res. 2019, 28, 299–315. [Google Scholar] [CrossRef]
- Soliman, H.I.; Abo-El-Hasan, F.M.; El-Seedy, A.S.; Mabrouk, Y.M. Agrobacterium-mediated transformation oftomato (Lycopersicon esculentum mill.) using a syntheticcry1ab gene for enhanced resistance against Tutaabsoluta (Meyrick). J. Microbiol. Biotech. Food Sci. 2017, 7, 67–74. [Google Scholar] [CrossRef]
- Muddanuru, T.; Polumetla, A.K.; Maddukuri, L.; Mulpuri, S. Development and evaluation of transgenic castor (Ricinus communis L.) expressing the insecticidal protein Cry1Aa of Bacillus thuringiensis against lepidopteran insect pests. Crop Protec. 2019, 119, 113–125. [Google Scholar] [CrossRef]
- Zhong, Y.; Ahmed, S.; Deng, G.; Fan, W.; Zhang, P.; Wang, H. Improved insect resistance against Spodopteralitura in transgenic sweet potato by overexpressing Cry1Aa toxin. Plant Cell Rep. 2019, 38, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Rakha, M.; Bouba, N.; Ramasamy, S.; Regnard, J.L.; Hanson, P. Evaluation of wild tomato accessions (Solanum spp.) for resistance to two-spotted spider mite (Tetranychusurticae Koch) based on trichome type and acylsugar content. Genet. Resour. Crop Evol. 2017, 64, 1011–1022. [Google Scholar] [CrossRef] [Green Version]
- Zsoögoön, A.; Cćermak, T.; Naves, E.R. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhu, L.; Hull, J.J. Transcriptome analysis reveals a comprehensive insect resistance response mechanism in cotton to infestation by the phloem feeding insect Bemisiatabaci (whitefly). Plant Biotechnol. J. 2016, 14, 1956–1975. [Google Scholar] [CrossRef]
- Unckless, R.L.; Clark, A.G.; Messer, P.W. Evolution of resistance against CRISPR/Cas9 gene drive. Genetics 2017, 205, 827–841. [Google Scholar] [CrossRef]
- Hammond, A.M.; Kyrou, K.; Bruttini, M. The creation and selection of mutations resistant to a gene drive over multiple generations in the malaria mosquito. PLoS Genet 2017, 13, e1007039. [Google Scholar] [CrossRef]
- Marshall, J.M.; Buchman, A.; Sanchez, C.H.M.; Akbari, O.S. Overcoming evolved resistance to population-suppressing homing-based gene drives. Sci. Rep. 2017, 7, 3776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, S.L. Unintended consequences of 21st century technology for agricultural pest management. EMBO Rep. 2017, 18, 1478. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, B.; Spalding, M.H.; Weeks, D.P.; Yang, B. High-efficiency TALEN-based gene editing produces disease-resistant rice. Nat.Biotechnol. 2012, 30, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.K.; Mahmud, N.U.; Islam, T. Impacts of Bt brinjal on economic benefit of farmers and environmental sustainability in Bangladesh. In Bacilli in Agrobiotechnology; Islam, M.T., Rahman, M., Pandey, P., Eds.; Bacilli in Climate Resilient Agriculture and Bioprospecting; Springer: Cham, Switzerland, 2022; pp. 539–560. [Google Scholar]
- Islam, M.T.; Bhowmik, P.K.; Molla, K.A. CRISPR-Cas Methods; Springer Nature: Berlin/Heidelberg, Germany, 2020; Volume 1, pp. 1–278. [Google Scholar]
- Islam, M.T.; Molla, K.A. CRISPR-Cas Methods; Springer Nature: Berlin/Heidelberg, Germany, 2021; Volume 2, pp. 1–397. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, T.T.; Maliha, I.J.; Khan, A.A.M.; Chakraborty, M.; Uddin, M.S.; Amin, M.R.; Islam, T. CRISPR-Cas Genome Editing for Insect Pest Stress Management in Crop Plants. Stresses 2022, 2, 493-514. https://doi.org/10.3390/stresses2040034
Moon TT, Maliha IJ, Khan AAM, Chakraborty M, Uddin MS, Amin MR, Islam T. CRISPR-Cas Genome Editing for Insect Pest Stress Management in Crop Plants. Stresses. 2022; 2(4):493-514. https://doi.org/10.3390/stresses2040034
Chicago/Turabian StyleMoon, Tasfia Tasnim, Ishrat Jahan Maliha, Abdullah Al Moin Khan, Moutoshi Chakraborty, Md Sharaf Uddin, Md Ruhul Amin, and Tofazzal Islam. 2022. "CRISPR-Cas Genome Editing for Insect Pest Stress Management in Crop Plants" Stresses 2, no. 4: 493-514. https://doi.org/10.3390/stresses2040034