
Involvement of Nitric Oxide in Methyl Jasmonate-Mediated Regulation of Water Metabolism in Wheat Plants under Drought Stress


Abstract
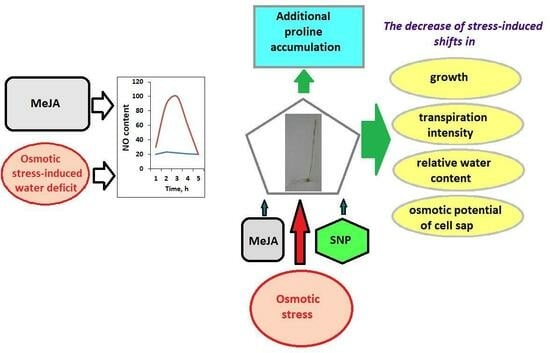
:
1. Introduction
2. Results
2.1. Growth Parameters
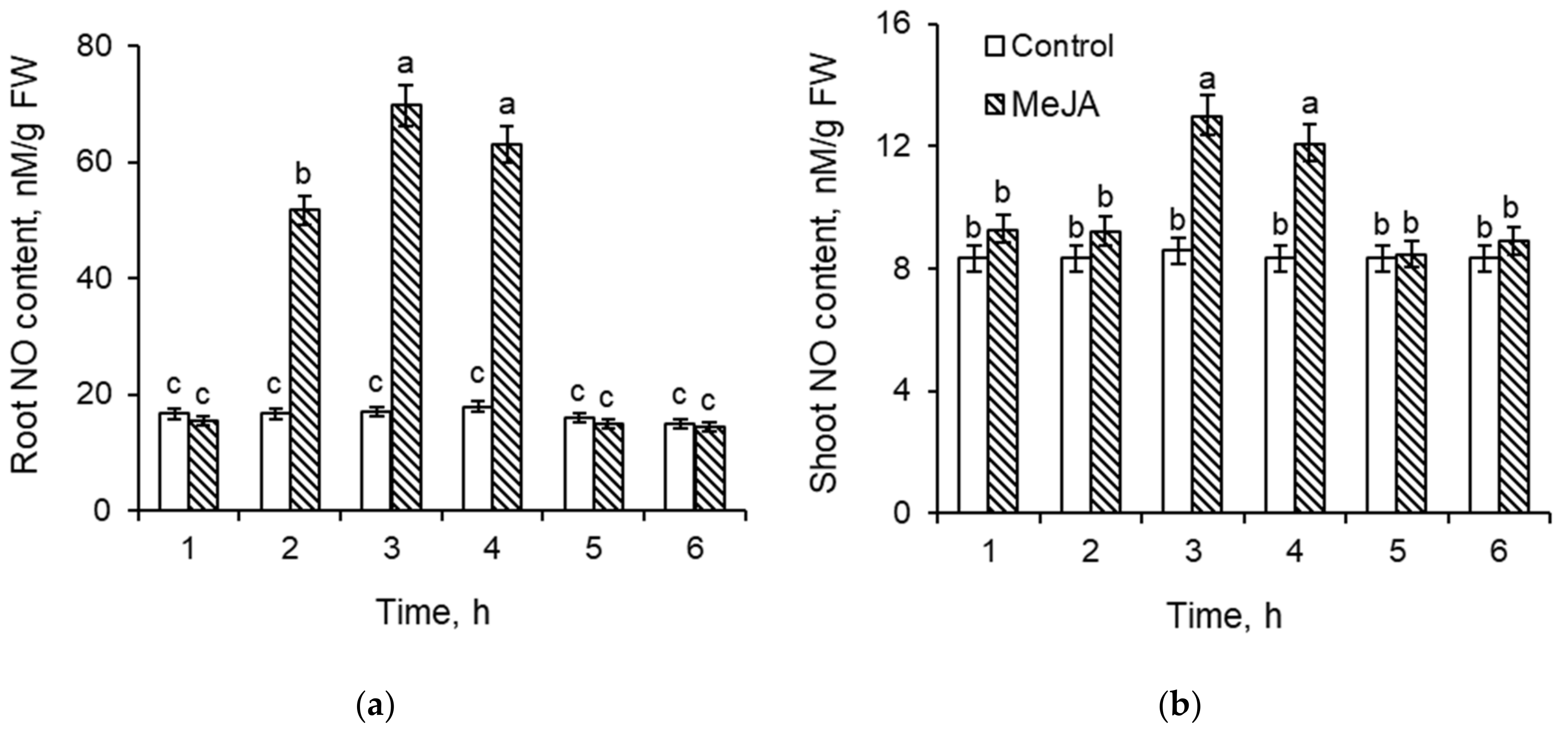
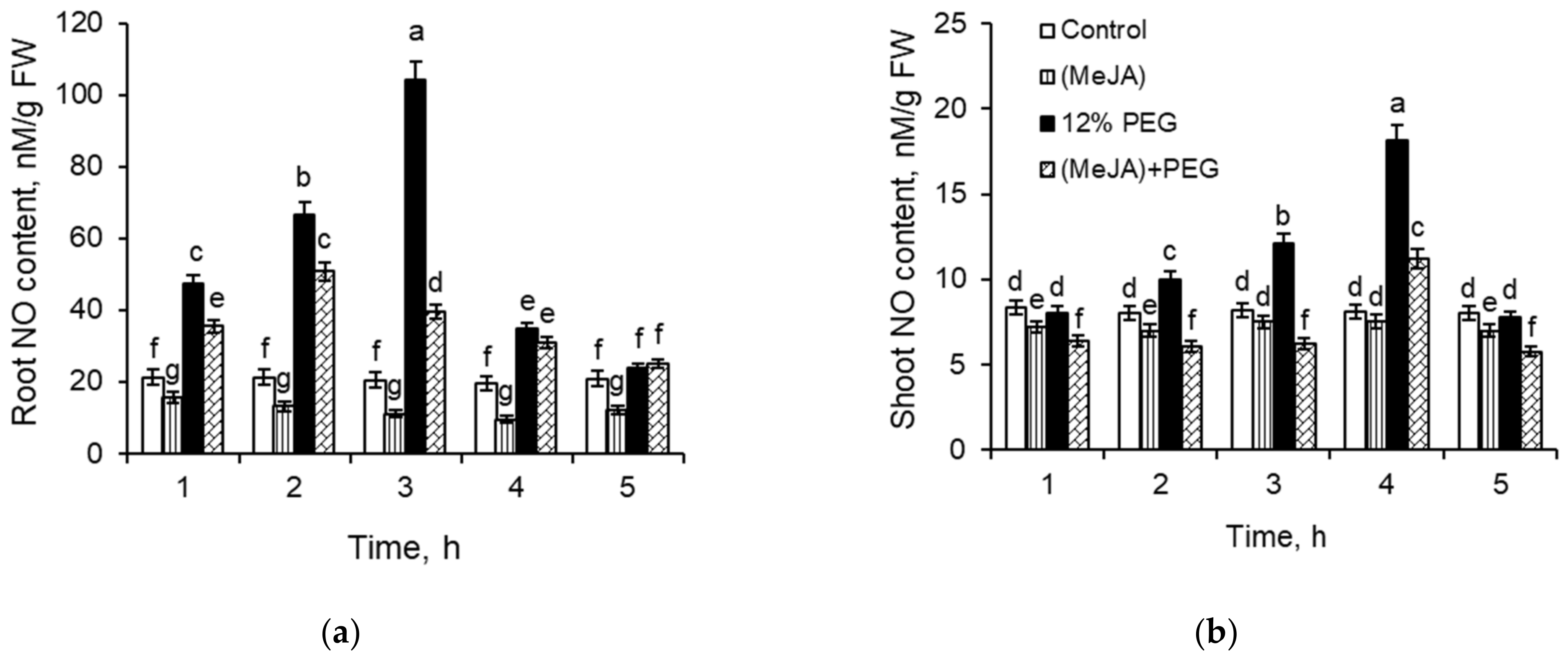
2.2. NO Content
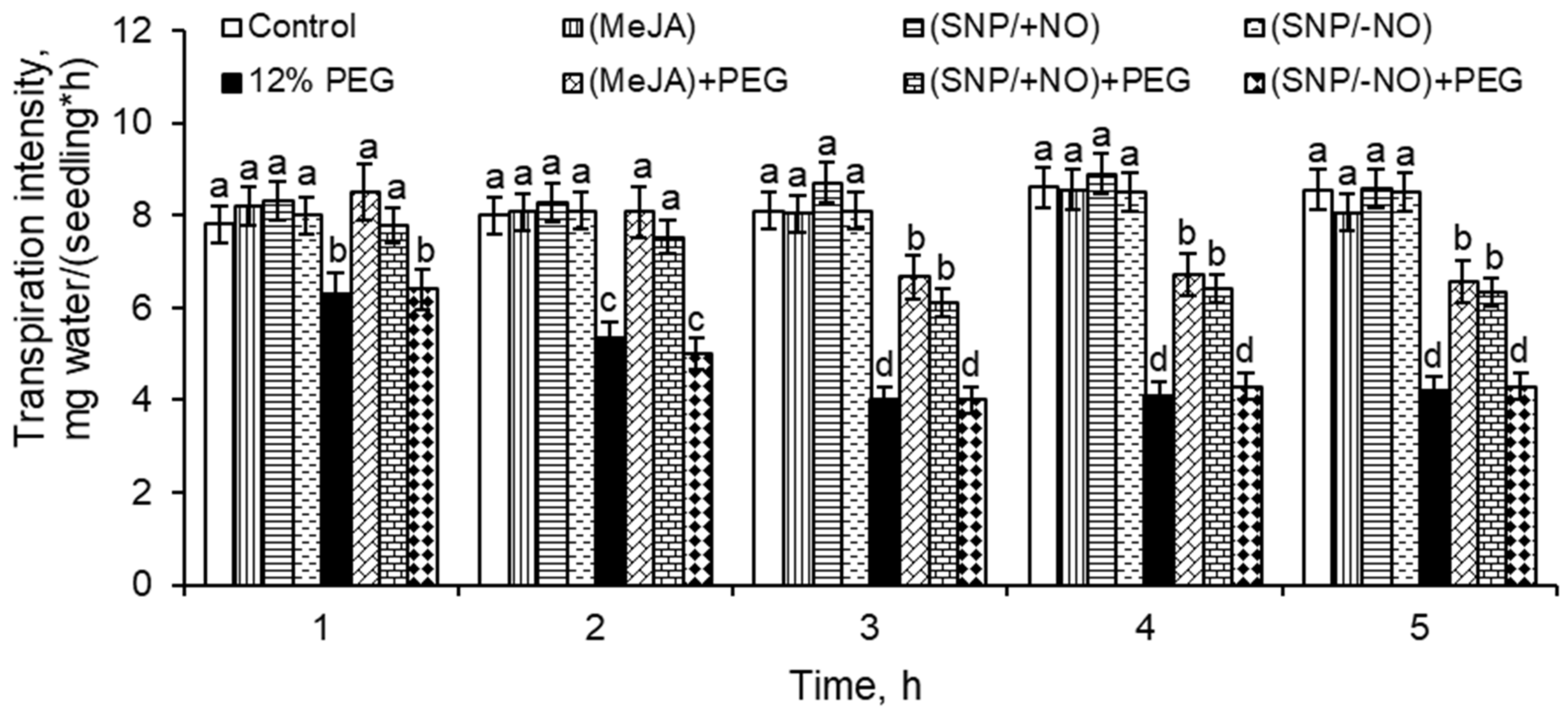
2.3. Transpiration Intensity
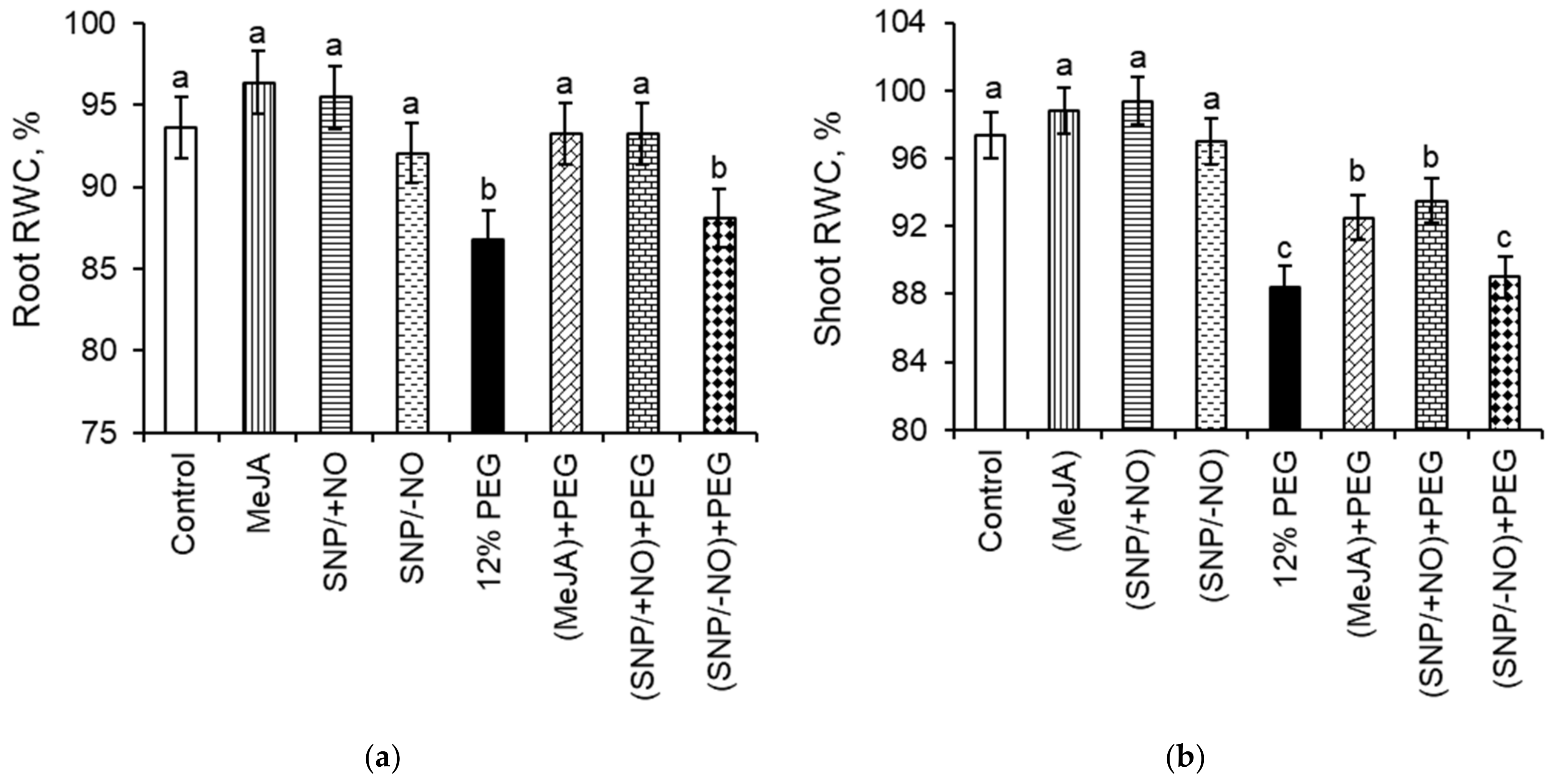
2.4. Relative Water Content (RWC)
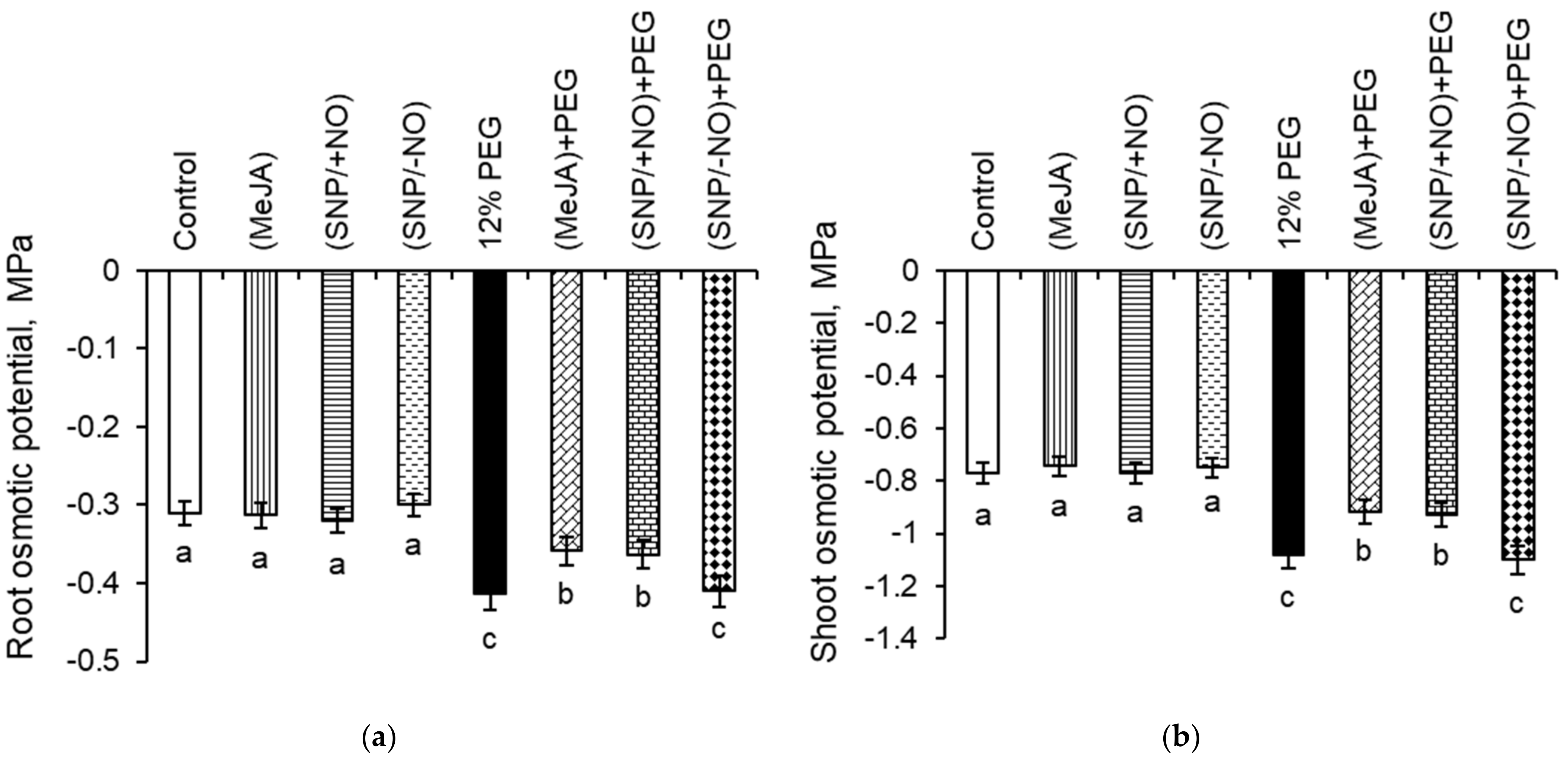
2.5. Osmotic Potential Estimation
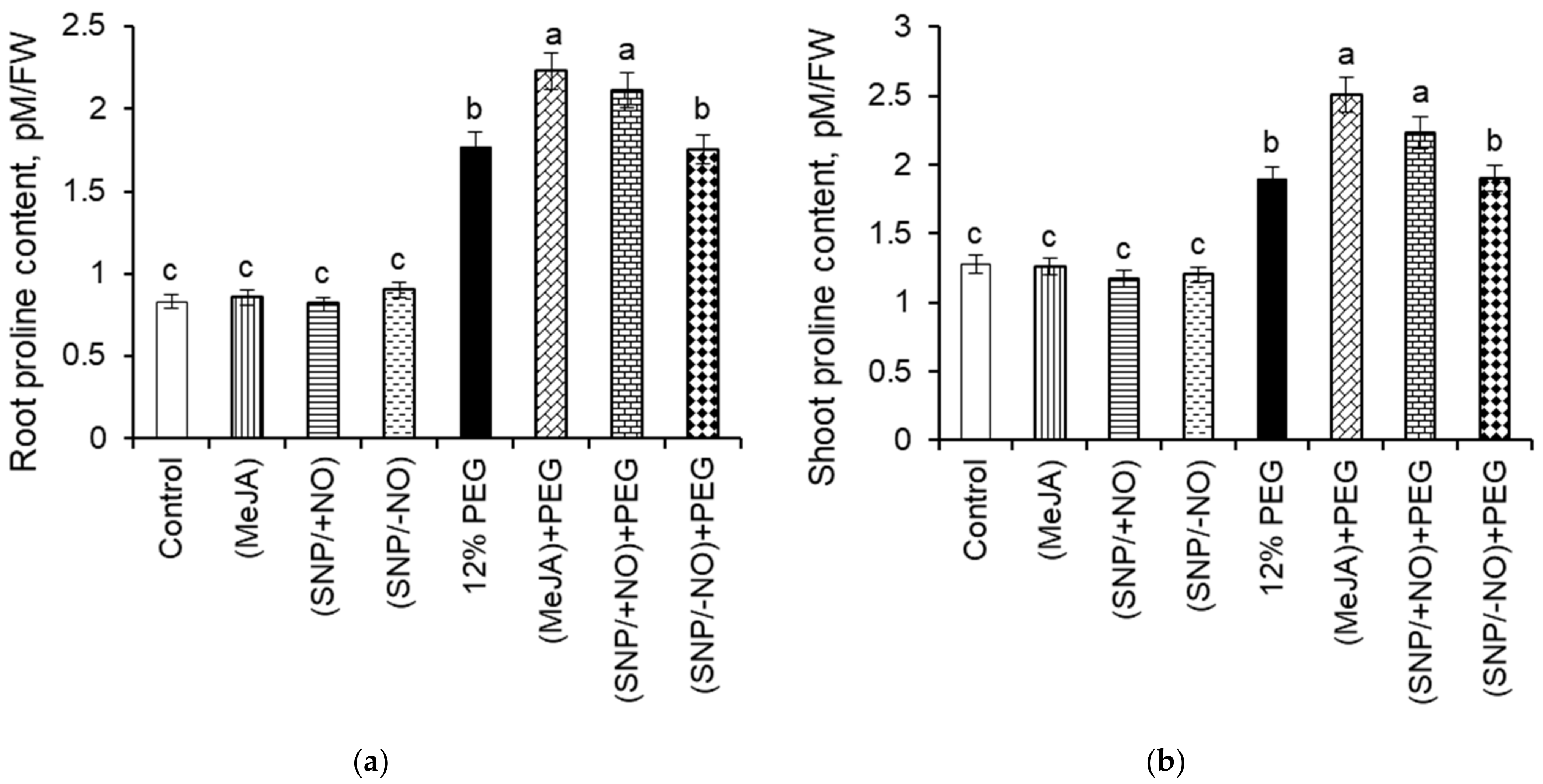
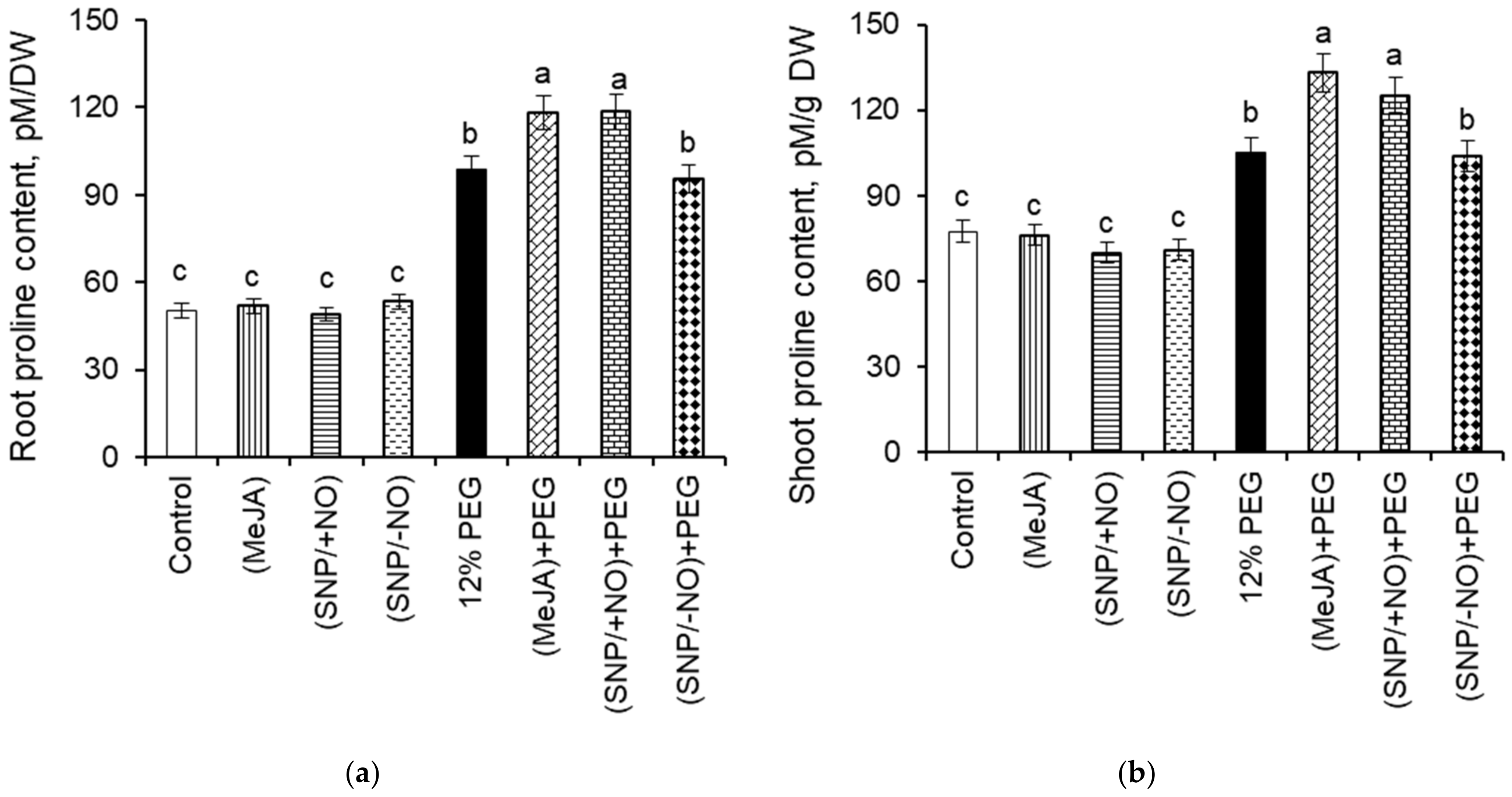
2.6. Proline Content
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. NO Measurement
4.3. Measurement of the Relative Water Content (RWC)
4.4. Estimation of the Transpiration Intensity
4.5. Osmotic Potential Estimation
4.6. Proline Accumulation
4.7. Estimation of Shoot Height, Root Length, and Leaf Area
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of Food and Agriculture 2020. Overcoming Water Challenges in Agriculture; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Mansoor, S.; Kour, N.; Manhas, S.; Zahid, S.; Wani, O.A.; Sharma, V.; Wijaya, L.; Alyemeni, M.N.; Alsahli, A.A.; El-Serehy, H.A.; et al. Biochar as a Tool for Effective Management of Drought and Heavy Metal Toxicity. Chemosphere 2021, 271, 129458. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guo, Y.; Cui, Q.; Zhang, Z.; Yan, X.; Ahammed, G.J.; Yang, X.; Yang, J.; Wei, C.; Zhang, X. Alkanes (C29 and C31)-mediated Intracuticular Wax Accumulation Contributes to Melatonin- and ABA-induced Drought Tolerance in Watermelon. J. Plant Growth Regul. 2020, 39, 1441–1450. [Google Scholar] [CrossRef]
- Bravo, L.A.; Gallardo, J.; Navarrete, A.; Olave, N.; Martínez, J.; Alberdi, M.; Close, T.J.; Corcuera, L.J. Cryoprotective Activity of a Cold-Induced Dehydrin Purified from Barley. Physiol. Plant. 2003, 118, 262–269. [Google Scholar] [CrossRef]
- Yang, T.; Yao, S.; Hao, L.; Zhao, Y.; Lu, W.; Xiao, K. Wheat bHLH-Type Transcription Factor Gene TabHLH1 is Crucial in Mediating Osmotic Stresses Tolerance through Modulating Largely the ABA-Associated Pathway. Plant Cell Rep. 2016, 35, 2309–2323. [Google Scholar] [CrossRef]
- Filippou, P.; Bouchagier, P.; Skotti, E.; Fotopoulos, V. Proline and Reactive Oxygen/Nitrogen Species Metabolism is Involved in the Tolerant Response of the Invasive Plant Species Ailanthus altissima to Drought and Salinity. Environ. Exp. Bot. 2014, 97, 1–10. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous Application of Nitric Oxide Modulates Osmolyte Metabolism, Antioxidants, Enzymes of Ascorbate-Glutathione Cycle and Promotes Growth under Cadmium Stress in Tomato. Protoplasma 2018, 255, 79–93. [Google Scholar] [CrossRef]
- De Ollas, C.; Dodd, I.C. Physiological Impacts of ABA–JA Interactions Under Water-limitation. Plant Mol. Biol. 2016, 91, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Riemann, M.; Dhakarey, R.; Hazman, M.; Miro, B.; Kohli, A.; Nick, P. Exploring Jasmonates in the Hormonal Network of Drought and Salinity Responses. Front. Plant Sci. 2015, 6, 1077. [Google Scholar] [CrossRef] [Green Version]
- Saibi, W.; Brini, F. Superoxide Dismutase (SOD) and Abiotic Stress Tolerance in Plants: An Overview. In Superoxide Dismutase; Editor Magliozzi, S., Ed.; Nova Science Publishers: New York, NY, USA, 2018; pp. 101–142. [Google Scholar]
- Daszkowska-Golec, A.; Szarejko, I. Open or Close the Gate—Stomata Action under the Control of Phytohormones in Drought Stress Conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef]
- Hsu, Y.Y.; Chao, Y.; Kao, C.H. Methyl Jasmonate-Induced Lateral Root Formation in Rice: The Role of Heme Oxygenase and Calcium. J. Plant Physiol. 2013, 170, 63–69. [Google Scholar] [CrossRef]
- Sánchez-Romera, B.; Ruiz-Lozano, J.M.; Li, G.; Luu, D.; Martínez-Ballesta, M.; Carvajal, M.; Zamarreño, A.M.; García-Mina, J.M.; Maurel, C.; Aroca, R. Enhancement of Root Hydraulic Conductivity by Methyl Jasmonate and the Role of Calcium and Abscisic Acid in this Process. Plant Cell Environ. 2014, 37, 995–1008. [Google Scholar] [CrossRef]
- Holland, C.K.; Jez, J.M. Structural Biology of Jasmonic Acid Metabolism and Responses in Plants. In Plant Structural Biology: Hormonal Regulations; Hejátko, J., Hakoshima, T., Eds.; Springer Nature Singapore Publishers: Cham, Switzerland, 2018; pp. 67–82. [Google Scholar] [CrossRef]
- De Ollas, C.; Arbona, V.; Gomez-Cadenas, A. Jasmonoyl Isoleucine Accumulation is Needed for Abscisic Acid Build-Up in Roots of Arabidopsis under Water Stress Conditions. Plant Cell Environ. 2015, 38, 2157–2170. [Google Scholar] [CrossRef] [Green Version]
- De Domenico, S.; Bonsegna, S.; Horres, R.; Pastor, V.; Taurino, M.; Poltronieri, P.; Imtiaz, M.; Kahl, G.; Flors, V.; Winter, P.; et al. Transcriptomic Analysis of Oxylipin Biosynthesis Genes and Chemical Profiling Reveal an Early Induction of Jasmonates in Hickpea Roots Under Drought Stress. Plant Physiol. Biochem. 2012, 61, 115–122. [Google Scholar] [CrossRef]
- Sirhindi, G.; Mir, M.A.; Abd-Allah, E.F.; Ahmad, P.; Gucel, S. Jasmonic Acid Modulates the Physio-Biochemical Attributes, Antioxidant Enzyme Activity, and Gene Expression in Glycine max under Nickel Toxicity. Front. Plant Sci. 2016, 7, 591. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y. Methyl Jasmonate Reduces Water Stress in Strawberry. J. Plant Growth Regul. 1999, 18, 127–134. [Google Scholar] [CrossRef]
- De Ollas, C.; Arbona, V.; Gómez-Cadenas, A.; Dodd, I.C. Attenuated Accumulation of Jasmonates Modifies Stomatal Responses to Water Deficit. J. Exp. Bot. 2018, 69, 2103–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, G.; Choi, Y.D. Drought Stress Promotes Xylem Differentiation by Modulating the Interaction Between Cytokinin and Jasmonic Acid. Plant Signal. Behav. 2018, 13, e1451707. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Lee, S.; Chang, S.H.; Kim, J.; Choi, Y.D. Jasmonic Acid Modulates Xylem Development by Controlling Polar Auxin Transport in Vascular Tissues. Plant Biotechnol. Rep. 2018, 12, 265–271. [Google Scholar] [CrossRef]
- Hung, K.T.; Kao, C.H. Nitric Oxide Acts as an Antioxidant and Delays Methyl Jasmonate-Induced Senescence of Rice Leaves. J. Plant Physiol. 2004, 161, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Avalbaev, A.; Yuldashev, R.; Fedorova, K.; Somov, K.; Vysotskaya, L.; Allagulova, C.; Shakirova, F. Exogenous Methyl Jasmonate Regulates Cytokinin Content by Modulating Cytokinin Oxidase Activity in Wheat Seedlings under Salinity. J. Plant Physiol. 2016, 191, 101–110. [Google Scholar] [CrossRef]
- Tavallali, V.; Karimi, S. Methyl Jasmonate Enhances Salt Tolerance of Almond Rootstocks by Regulating Endogenous Phytohormones, Antioxidant Activity and Gasexchange. J. Plant Physiol. 2019, 234–235, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Albertos, P.; Mateos, I.; Sanchez-Vicente, I.; Lechon, T.; Fernández-Marcos, M.; Lorenzo, O. Nitric Oxide (NO) and Phytohormones Crosstalk during Early Plant Development. J. Exp. Bot. 2015, 66, 2857–2868. [Google Scholar] [CrossRef] [PubMed]
- Santisree, P.; Bhatnagar-Mathur, P.; Sharma, K.K. NO to Drought-Multifunctional Role of Nitric Oxide in Plant Drought: Do We Have All the Answers? Plant Sci. 2015, 239, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Sami, F.; Faizan, M.; Faraz, A.; Siddiqui, H.; Yusuf, M.; Hayat, S. Nitric Oxide-Mediated Integrative Alterations in Plant Metabolism to Confer Abiotic Stress Tolerance, NO Crosstalk With Phytohormones and NO-Mediated Post Translational Modifications in Modulating Diverse Plant Stress. Nitric Oxide 2018, 73, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.J.; Liu, J.H. Nitric Oxide is Involved in Dehydration/Drought Tolerance in Poncirus trifoliata Seedlings through Regulation of Antioxidant Systems and Stomatal Response. Plant Cell Rep. 2012, 31, 145–154. [Google Scholar] [CrossRef]
- García-Mata, C.; Lamattina, L. Nitric Oxide Induces Stomatal Closure and Enhances the Adaptive Plant Responses Against Drought Stress. Plant Physiol. 2001, 126, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- García-Mata, C.; Lamattina, L. Gasotransmitters are Emerging As New Guard Cell Signaling Molecules and Regulators of Leaf Gas Exchange. Plant Sci. 2013, 201, 66–73. [Google Scholar] [CrossRef]
- Ahmad, P.; Abass Ahanger, M.; Nasser Alyemeni, M.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of Sodium Chloride Toxicity in Solanum lycopersicum L. by Supplementation of Jasmonic Acid and Nitric Oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef]
- Yastreb, T.O.; Kolupaev, Y.E.; Shkliarevskyi, M.A.; Dmitriev, A.P. Participation of Jasmonate Signaling Components in the Development of Arabidopsis thaliana’s Salt Resistance Induced by H2S and NO Donors. Russ. J. Plant Physiol. 2020, 67, 827–834. [Google Scholar] [CrossRef]
- Tan, J.; Zhao, H.; Hong, J.; Han, Y.; Li, H.; Zhao, W. Effects of Exogenous Nitric Oxide on Photosynthesis, Antioxidant Capacity and Proline Accumulation in Wheat Seedlings Subjected to Osmotic Stress. World J. Agric. Sci. 2008, 4, 307–313. [Google Scholar]
- Paul, S.; Roychoudhury, A. Regulation of Physiological Aspects in Plants by Hydrogen Sulfide and Nitric Oxide Under Challenging Environment. Physiol. Plant. 2020, 168, 374–393. [Google Scholar] [CrossRef] [Green Version]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.G.; Yun, B.W. Nitric Oxide Regulates Plant Responses to Drought, Salinity, and Heavy Metal Stress. EEB 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Role of Nitric Oxide in Tolerance of Plants to Abiotic Stress. Protoplasma 2011, 248, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.; Li, X.; Zhang, H.; Zhang, G.; Liu, Y.; Zhou, Y.; Xia, X.; Chen, Z.; Yu, J. Guard Cell Hydrogen Peroxide and Nitric Oxide Mediate Elevated CO2-Induced Stomatal Movement in Tomato. New Phytol. 2015, 208, 342–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, P.; Rasool, S.; Gul, A.; Sheikh, S.A.; Akram, N.A.; Ashraf, M.; Kazi, A.M.; Gucel, S. Jasmonates: Multifunctional Roles in Stress Tolerance. Front. Plant Sci. 2016, 7, 813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Stettmaier, K.; Hutzler, C.; Mueller, M.; Durner, J. Nitric Oxide is Induced by Wounding and Influences Jasmonic Acid Signaling in Arabidopsis thaliana. Planta 2004, 218, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.; Barros, R.; Bright, J.; Desikan, R.; Hancock, J.; Harrison, J.; Morris, P.; Ribeiro, D.; Wilson, I. Nitric Oxide, Stomatal Closure, and Abiotic Stress. J. Exp. Bot. 2008, 59, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Salt and Drought Stress Signal Transduction in Plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluskiewicz, B.; Baba, W.; Brestic, M. Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef]
- Petrov, P.; Petrova, A.; Dimitrov, I.; Tashev, T.; Olsovska, K.; Brestic, M.; Misheva, S. Relationships between Leaf Morpho-Anatomy, Water Status and Cell Membrane Stability in Leaves of Wheat Seedlings Subjected to Severe Soil Drought. J. Agron. Crop Sci. 2018, 204, 219–227. [Google Scholar] [CrossRef]
- Lubyanova, A.R.; Bezrukova, M.V.; Shakirova, F.M. Interaction between Signal Pathways upon Formation of Plant Defense in Response to Environmental Stress Factors. Russ. J. Plant Physiol. 2021, 68, 989–1002. [Google Scholar] [CrossRef]
- Bhat, J.A.; Ahmad, P.; Corpas, F.J. Main Nitric Oxide (NO) Hallmarks to Relieve Arsenic Stress in Higher Plants. J. Hazard. Mater. 2021, 406, 124289. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Li, X.; Wan, H.; Zhou, G.; Cheng, Y. SlWRKY81 Reduces Drought Tolerance by Attenuating Proline Biosynthesis in Tomato. Sci. Hortic. 2020, 270, 109444. [Google Scholar] [CrossRef]
- Farmer, E.E.; Ryan, C.A. Interplant Communication: Airborne Methyl Jasmonate Induces Synthesis of Proteinase Inhibitors in Plant Leaves. Proc. Natl. Acad. Sci. USA 1990, 87, 7713–7716. [Google Scholar] [CrossRef] [Green Version]
- Evans, N.H. Modulation of Guard Cell Plasma Membrane Potassium Currents by Methyl Jasmonate. Plant Physiol. 2003, 131, 8–11. [Google Scholar] [CrossRef] [Green Version]
- Mur, L.A.; Prats, E.; Pierre, S.; Hall, M.A.; Hebelstrup, K.H. Integrating Nitric Oxide into Salicylic Acid and Jasmonic Acid/Ethylene Plant Defense Pathways. Front. Plant Sci. 2013, 4, 215. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Chen, G.; Zhang, C. Interaction between Reactive Oxygen Species and Nitric Oxide in Drought-Induced Abscisic Acid Synthesis in Root Tips of Wheat Seedlings. Funct. Plant Biol. 2001, 28, 1055–1061. [Google Scholar] [CrossRef]
- Solórzano, E.; Corpas, F.J.; González-Gordo, S.; Palma, J.M. Reactive Oxygen Species (ROS) Metabolism and Nitric Oxide (NO) Content in Roots and Shoots of Rice (Oryza sativa L.) Plants under Arsenic-Induced Stress. Agronomy 2020, 10, 1014. [Google Scholar] [CrossRef]
- Batista, P.F.; Costa, A.C.; Müller, C.; de Oliveira Silva-Filho, R.; da Silva, F.B.; Merchant, A.; Mendes, G.C.; Nascimento, K.J.T. Nitric Oxide Mitigates the Effect of Water Deficit in Crambe abyssinica. Plant Physiol. Biochem. 2018, 129, 310–322. [Google Scholar] [CrossRef]
- Montilla-Bascón, G.; Rubiales, D.; Hebelstrup, K.H.; Mandon, J.; Harren, F.J.M.; Cristescu, S.M.; Mur, L.A.J.; Prats, E. Reduced Nitric Oxide Levels During Drought Stress Promote Drought Tolerance in Barley and is Associated with Elevated Polyamine Biosynthesis. Sci. Rep. 2017, 7, 13311. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Gan, L.; Wu, X.; Zhong, Y. Exogenously Applied Nitric Oxide Enhances the Drought Tolerance in Hulless Barley. Plant Prod. Sci. 2015, 18, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Maslennikova, D.R.; Allagulova, C.R.; Fedorova, K.A.; Plotnikov, A.A.; Avalbaev, A.M.; Shakirova, F.M. Cytokinins Contribute to Realization of Nitric Oxide Growth-Stimulating and Protective Effects on Wheat Plants. Russ. J. Plant Physiol. 2017, 64, 665–671. [Google Scholar] [CrossRef]
- Bezrukova, M.V.; Kudoyarova, G.R.; Lubyanova, A.R.; Maslennikova, D.R.; Shakirova, F.M. Influence of 24-Epibrassinolide on Water Exchange of Wheat Varieties Various in Dry Resistance Under Osmotic Stress. Russ. J. Plant Physiol. 2021, 68, 307–314. [Google Scholar] [CrossRef]
- Sharipova, G.V.; Veselov, D.S.; Kudoyarova, G.R.; Timergalin, M.D.; Wilkinson, S. Effect of Ethylene Perception Inhibitor on Growth, Water Relations, and Abscisic Acid Content in Wheat Plants Under Water Deficit. Russ. J. Plant Physiol. 2012, 59, 573–580. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, S.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stressed Studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Root Length, cm | Shoot Height, cm | Leaf Area, cm2 |
|---|---|---|---|
| Control | 7.22 ± 0.80 | 15.75 ± 0.93 | 3.04 ± 0.55 |
| (MeJA) | 10.07 ± 1.32 (140) * | 16.80 ± 1.51 (107) | 3.43 ± 0.61 (113) ns |
| (SNP/+NO) | 9.00 ± 1.29 (125) * | 16.49 ± 1.45 (105) | 3.36 ± 0.43 (111) ns |
| (SNP/−NO) | 7.36 ± 0.74 (102) ns | 15.77 ± 1.76 (100) | 3.07 ± 0.43 (101) ns |
| 12% PEG | 6.71 ± 0.92 (93) ns | 14.48 ± 1.84 (92) | 2.83 ± 0.61 (93) ns |
| (MeJA)+PEG | 8.34 ± 1.43 (116) ns | 16.13 ± 2.08 (102) | 3.19 ± 0.58 (105) ns |
| (SNP/+NO)+PEG | 8.10 ± 1.23 (112) ns | 16.05 ± 1.45 (102) | 3.17 ± 0.38 (104) ns |
| (SNP/−NO)+PEG | 7.19 ± 1.37 (100) ns | 14.50 ± 1.38 (92) | 2.88 ± 0.39 (95) ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lubyanova, A.R.; Bezrukova, M.V.; Shakirova, F.M. Involvement of Nitric Oxide in Methyl Jasmonate-Mediated Regulation of Water Metabolism in Wheat Plants under Drought Stress. Stresses 2022, 2, 477-492. https://doi.org/10.3390/stresses2040033
Lubyanova AR, Bezrukova MV, Shakirova FM. Involvement of Nitric Oxide in Methyl Jasmonate-Mediated Regulation of Water Metabolism in Wheat Plants under Drought Stress. Stresses. 2022; 2(4):477-492. https://doi.org/10.3390/stresses2040033
Chicago/Turabian StyleLubyanova, Alsu R., Marina V. Bezrukova, and Farida M. Shakirova. 2022. "Involvement of Nitric Oxide in Methyl Jasmonate-Mediated Regulation of Water Metabolism in Wheat Plants under Drought Stress" Stresses 2, no. 4: 477-492. https://doi.org/10.3390/stresses2040033