
Toxoplasma gondii in Marine Life of Italian Coasts, What Do We Know So Far?


{kind=link}
Abstract
:1. Introduction
2. A Closer Look into Italy
3. Distribution of T. gondii in Italy
4. Toxoplasma gondii: From Land to Sea
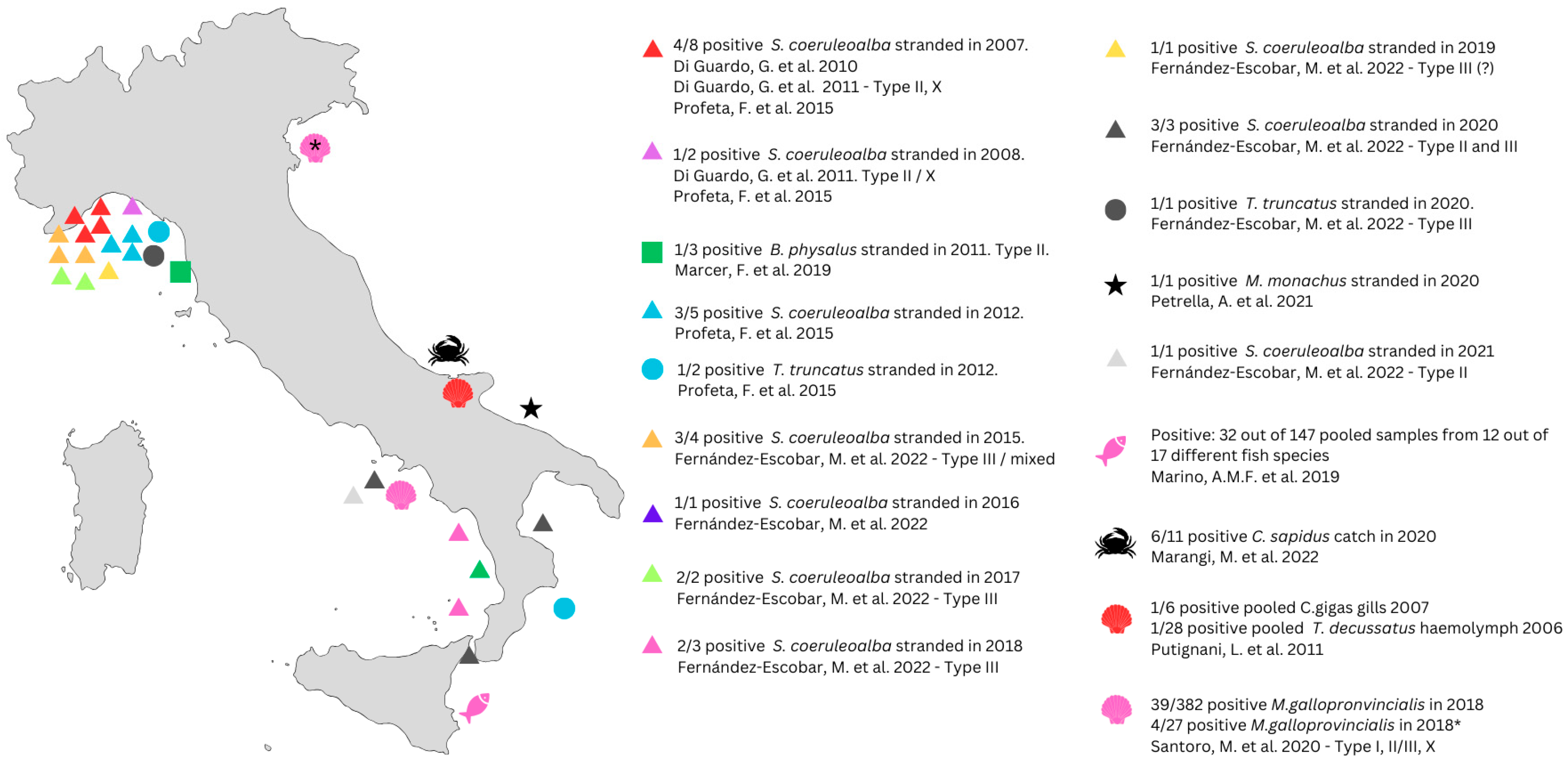
5. Toxoplasma gondii in the Italian Sea Environment
5.1. Marine Mammals
5.2. Fish
5.3. Shellfish

6. Concluding Remarks
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morozińska-Gogol, J. The presence of Toxoplasma gondii in the terrestrial and marine environments and its importance for public health. Ann. Parasitol. 2021, 67, 137–149. [Google Scholar] [PubMed]
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Lélu, M.; Villena, I.; Dardé, M.-L.; Aubert, D.; Geers, R.; Dupuis, E.; Marnef, F.; Poulle, M.-L.; Gotteland, C.; Dumètre, A.; et al. Quantitative Estimation of the Viability of Toxoplasma gondii Oocysts in Soil. Appl. Environ. Microbiol. 2012, 78, 5127–5132. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasma gondii oocyst survival under defined temperatures. J. Parasitol. 1998, 84, 862–865. [Google Scholar] [CrossRef] [PubMed]
- Pereira, K.S.; Franco, R.M.B.; Leal, D.A.G. Chapter 1—Transmission of Toxoplasmosis (Toxoplasma gondii) by Foods. In Advances in Food and Nutrition Research; Taylor, S.L., Ed.; Academic Press: Cambridge, MA, USA, 2010; pp. 1–19. Available online: https://www.sciencedirect.com/science/article/pii/S1043452610600010 (accessed on 8 August 2023).
- Robert-Gangneux, F.; Dardé, M.-L. Epidemiology of and Diagnostic Strategies for Toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Trop. 2001, 78, 103–116. [Google Scholar] [CrossRef]
- Baker, R.E.; Mahmud, A.S.; Miller, I.F.; Rajeev, M.; Rasambainarivo, F.; Rice, B.L.; Takahashi, S.; Tatem, A.J.; Wagner, C.E.; Wang, L.-F.; et al. Infectious disease in an era of global change. Nat. Rev. Microbiol. 2022, 20, 193–205. [Google Scholar] [CrossRef]
- Shapiro, K. Climate and coastal habitat change: A recipe for a dirtier ocean. Mar. Pollut. Bull. 2012, 64, 1079–1080. [Google Scholar] [CrossRef]
- Di Guardo, G.; Mazzariol, S. Cetacean morbillivirus: A Land-to-Sea Journey and Back? Virol. Sin. 2019, 34, 240–242. [Google Scholar] [CrossRef]
- Gulland, F.M.D.; Hall, A.J.; Ylitalo, G.M.; Colegrove, K.M.; Norris, T.; Duignan, P.J.; Halaska, B.; Whitehouse, K.A.; Lowenstine, L.J.; Deming, A.C.; et al. Persistent Contaminants and Herpesvirus OtHV1 Are Positively Associated With Cancer in Wild California Sea Lions (Zalophus californianus). Front. Mar. Sci. 2020, 7, 1093. Available online: https://www.frontiersin.org/articles/10.3389/fmars.2020.602565 (accessed on 4 October 2023). [CrossRef]
- Nymo, I.H.; Tryland, M.; Godfroid, J. A review of Brucella infection in marine mammals, with special emphasis on Brucella pinnipedialis in the hooded seal (Cystophora cristata). Vet. Res. 2011, 42, 93. [Google Scholar] [CrossRef]
- Thomas, N.; White, C.L.; Saliki, J.; Schuler, K.; Lynch, D.; Nielsen, O.; Dubey, J.P.; Knowles, S. Canine distemper virus in the sea otter (Ehydra lutris) population in Washington Estate, USA. J. Wildl. Dis. 2020, 56, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Osterhaus, A.D.; Rimmelzwaan, G.F.; Martina, B.E.; Bestebroer, T.M.; Fouchier, R.A. Influenza B virus in seals. Science 2000, 288, 1051–1053. [Google Scholar] [CrossRef]
- Fayer, R.; Dubey, J.P.; Lindsay, D.S. Zoonotic protozoa: From land to sea. Trends Parasitol. 2004, 20, 531–536. [Google Scholar] [CrossRef]
- Bossart, G.D. Marine Mammals as Sentinel Species for Oceans and Human Health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Doney, S.C. The growing human footprint on coastal and open-ocean biogeochemistry. Science 2010, 328, 1512–1516. [Google Scholar] [CrossRef] [PubMed]
- Hylkema, A.; Kitson-Walters, K.; Kramer, P.R.; Patterson, J.T.; Roth, L.; Sevier, M.L.B.; Vega-Rodriguez, M.; Warham, M.M.; Williams, S.M.; Lang, J.C. The 2022 Diadema antillarum die-off event: Comparisons with the 1983–1984 mass mortality. Front. Mar. Sci. 2023, 9, 2654. Available online: https://www.frontiersin.org/articles/10.3389/fmars.2022.1067449 (accessed on 8 August 2023). [CrossRef]
- Kreuder, C.; Miller, M.A.; Jessup, D.A.; Lowenstine, L.J.; Harris, M.D.; Ames, J.A.; Carpenter, T.E.; Conrad, P.A.; Mazet, J.A.K. Patterns of mortality in southern sea otters (Enhydra lutris nereis) from 1998–2001. J. Wildl. Dis. 2003, 39, 495–509. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D′Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef]
- Harvell, D.; Aronson, R.; Baron, N.; Connell, J.; Dobson, A.; Ellner, S.; Gerber, L.; Kim, K.; Kuris, A.; McCallum, H.; et al. The rising tide of ocean diseases: Unsolved problems and research priorities. Front. Ecol. Environ. 2004, 2, 375–382. [Google Scholar] [CrossRef]
- Benenson, M.W.; Takafuji, E.T.; Lemon, S.M.; Greenup, R.L.; Sulzer, A.J. Oocyst-transmitted toxoplasmosis associated with ingestion of contaminated water. N. Engl. J. Med. 1982, 307, 666–669. [Google Scholar] [CrossRef]
- Aramini, J.J.; Stephen, C.; Dubey, J.P.; Engelstoft, C.; Schwantje, H.; Ribble, C.S. Potential contamination of drinking water with Toxoplasma gondii oocysts. Epidemiol. Infect. 1999, 122, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumètre, A.; de Wit, L.A.; VanWormer, E.; Villena, I. Environmental transmission of Toxoplasma gondii: Oocysts in water, soil and food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef] [PubMed]
- Simone, M. La Dinamica Demografica del 2022; Istituto Nazionale di Statistica (ISTAT): Rome, Italy, Comunicato Stampa. Available online: https://www.istat.it/it/archivio/282316 (accessed on 20 October 2023).
- Mirto, A.P.M. Attività Antropiche e Salute delle Coste. Indicatori Territoriali di Rischio e Sostenibilità per Aree Costiere e Insulari in Italia e nell’Unione Europea; Istituto Nazionale di Statistica (ISTAT): Rome, Italy, 2022; ISBN 978-88-458-2084-7. Available online: https://www.istat.it/it/archivio/274891 (accessed on 20 October 2023).
- Keesing, F.; Ostfeld, R.S. Impacts of biodiversity and biodiversity loss on zoonotic diseases. Proc. Natl. Acad. Sci. USA 2021, 118, e2023540118. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.G.; Wilson, S.; Alavi, N.; Lapen, D.R. Human density is associated with the increased prevalence of a generalist zoonotic parasite in mammalian wildlife. Proc. R. Soc. B Biol. Sci. 2021, 288, 20211724. [Google Scholar] [CrossRef] [PubMed]
- Conrad, P.; Miller, M.; Kreuder, C.; James, E.; Mazet, J.; Dabritz, H.; Jessup, D.; Gulland, F.; Grigg, M. Transmission of Toxoplasma: Clues from the study of sea otters as sentinels of Toxoplasma gondii flow into the marine environment. Int. J. Parasitol. 2005, 35, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the public health hazards to be covered by inspection of meat from farmed game. EFSA J. 2013, 11, 3264. [Google Scholar] [CrossRef]
- World Health Organization. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015. World Health Organization: Geneva, Switzerland, 2015. Available online: https://apps.who.int/iris/handle/10665/199350 (accessed on 20 October 2023).
- Battisti, E.; Zanet, S.; Trisciuoglio, A.; Bruno, S.; Ferroglio, E. Circulating genotypes of Toxoplasma gondii in Northwestern Italy. Vet. Parasitol. 2018, 253, 43–47. [Google Scholar] [CrossRef]
- Rocchigiani, G.; Nardoni, S.; D′Ascenzi, C.; Nicoloso, S.; Picciolli, F.; Papini, R.A.; Mancianti, F. Seroprevalence of Toxoplasma gondii and Neospora caninum in red deer from Central Italy. Ann. Agric. Environ. Med. 2016, 23, 699–701. [Google Scholar] [CrossRef]
- Dini, F.; Morselli, S.; Marangoni, A.; Taddei, R.; Maioli, G.; Roncarati, G.; Balboni, A.; Dondi, F.; Lunetta, F.; Galuppi, R. Spread of Toxoplasma gondii among animals and humans in Northern Italy: A retrospective analysis in a One-Health framework. Food Waterborne Parasitol. 2023, 32, e00197. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Zanzani, S.A.; Villa, L.; Manfredi, M.T. Toxoplasma gondii infection in meat-producing small ruminants: Meat juice serology and genotyping. Parasitol. Int. 2020, 76, 102060. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Marino, A.M.F.; Garippa, G.; Rossi, L.; Mignone, W.; Dini, V.; Giunta, R.P.; Luini, M.; Villa, L.; Zanzani, S.A.; et al. Toxoplasma gondii seroprevalence in beef cattle raised in Italy: A multicenter study. Parasitol. Res. 2020, 119, 3893–3898. [Google Scholar] [CrossRef] [PubMed]
- Vesco, G.; Buffolano, W.; La Chiusa, S.; Mancuso, G.; Caracappa, S.; Chianca, A.; Villari, S.; Currò, V.; Liga, F.; Petersen, E. Toxoplasma gondii infections in sheep in Sicily, southern Italy. Vet. Parasitol. 2007, 146, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Fusco, G.; Rinaldi, L.; Guarino, A.; Proroga, Y.T.R.; Pesce, A.; Giuseppina, D.M.; Cringoli, G. Toxoplasma gondii in sheep from the Campania region (Italy). Vet. Parasitol. 2007, 149, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Ciuca, L.; Borriello, G.; Bosco, A.; D’Andrea, L.; Cringoli, G.; Ciaramella, P.; Maurelli, M.P.; Di Loria, A.; Rinaldi, L.; Guccione, J. Seroprevalence and Clinical Outcomes of Neospora caninum, Toxoplasma gondii and Besnoitia besnoiti Infections in Water Buffaloes (Bubalus bubalis). Animals 2020, 10, 532. [Google Scholar] [CrossRef] [PubMed]
- Sgroi, G.; Viscardi, M.; Santoro, M.; Borriello, G.; D′Alessio, N.; Boccia, F.; Pacifico, L.; Fioretti, A.; Veneziano, V.; Fusco, G. Genotyping of Toxoplasma gondii in wild boar (Sus scrofa) in southern Italy: Epidemiological survey and associated risk for consumers. Zoonoses Public Health 2020, 67, 805–813. [Google Scholar] [CrossRef]
- Dubey, J.P.; Miller, N.L.; Frenkel, J.K. The Toxoplasma gondii oocysts from cat feces. J. Exp. Med. 1970, 132, 636–662. [Google Scholar] [CrossRef] [PubMed]
- FEDIAF. European Pet Food, Annual Report 2023. Available online: www.europeanpetfood.org (accessed on 15 November 2023).
- Genchi, M.; Vismarra, A.; Zanet, S.; Morelli, S.; Galuppi, R.; Cringoli, G.; Lia, R.; Diaferia, M.; di Regalbono, A.F.; Venegoni, G.; et al. Prevalence and risk factors associated with cat parasites in Italy: A multicenter study. Parasites Vectors 2021, 14, 475. [Google Scholar] [CrossRef]
- Carvelli, A.; Iacoponi, F.; Scaramozzino, P. A Cross-Sectional Survey to Estimate the Cat Population and Ownership Profiles in a Semirural Area of Central Italy. BioMed Res. Int. 2016, 2016, e3796872. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Feline toxoplasmosis and coccidiosis: A survey of domiciled and stray cats. J. Am. Vet. Med. Assoc. 1973, 162, 873–877. [Google Scholar]
- Papini, R.; Sbrana, C.; Rosa, B.; Saturni, A.M.; Sorrentino, A.M.; Cerretani, M.; Raffaelli, G.; Guidi, G. Serological survey of Toxoplasma gondii infections in stray cats from Italy. Rev. Med. Vet. 2006, 157, 193–196. [Google Scholar]
- Vignani, D.; Budano, F. Istat, Istituto Nazionale di Statistica. Temperatura e Precipitazione delle Città Capoluogo Negli Anni 1971–2021. Report, 17 May 2023. Available online: www.istat.it (accessed on 20 October 2023).
- Horn, J.A.; Mateus-Pinilla, N.; Warner, R.E.; Heske, E.J. Home range, habitat use, and activity patterns of free-roaming domestic cats. J. Wildl. Manag. 2011, 75, 1177–1185. [Google Scholar] [CrossRef]
- Marino, A.M.F.; Giunta, R.P.; Salvaggio, A.; Castello, A.; Alfonzetti, T.; Barbagallo, A.; Aparo, A.; Scalzo, F.; Reale, S.; Buffolano, W.; et al. Toxoplasma gondii in edible fishes captured in the Mediterranean basin. Zoonoses Public Health 2019, 66, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Cabezón, O.; Hall, A.; Vincent, C.; Pabón, M.; García-Bocanegra, I.; Dubey, J.; Almería, S. Seroprevalence of Toxoplasma gondii in North-eastern Atlantic harbor seal (Phoca vitulina vitulina) and grey seal (Halichoerus grypus). Vet. Parasitol. 2011, 179, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Michael, S.A.; Howe, L.; Chilvers, B.L.; Morel, P.; Roe, W.D. Seroprevalence of Toxoplasma gondii in mainland and sub-Antarctic New Zealand sea lion (Phocarctos hookeri) populations. N. Z. Vet. J. 2016, 64, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, G.; Fichi, G.; Comentale, A.; Ricci, E.; Mancusi, C.; Perrucci, S. Dolphins Stranded along the Tuscan Coastline (Central Italy) of the “Pelagos Sanctuary”: A Parasitological Investigation. Pathogens 2020, 9, 612. [Google Scholar] [CrossRef]
- VanWormer, E.; Carpenter, T.E.; Singh, P.; Shapiro, K.; Wallender, W.W.; Conrad, P.A.; Largier, J.L.; Maneta, M.P.; Mazet, J.A.K. Coastal development and precipitation drive pathogen flow from land to sea: Evidence from a Toxoplasma gondii and felid host system. Sci. Rep. 2016, 6, 29252. [Google Scholar] [CrossRef]
- Cole, R.A.; Lindsay, D.S.; Howe, D.K.; Roderick, C.L.; Dubey, J.P.; Thomas, N.J.; Baeten, L.A. Biological and Molecular Characterizations of Toxoplasma gondii Strains Obtained from Southern Sea Otters (Enhydra lutris nereis). J. Parasitol. 2000, 86, 526–530. [Google Scholar] [CrossRef]
- Zhang, E.; Kim, M.; Rueda, L.; Rochman, C.; VanWormer, E.; Moore, J.; Shapiro, K. Association of zoonotic protozoan parasites with microplastics in seawater and implications for human and wildlife health. Sci. Rep. 2022, 12, 6532. [Google Scholar] [CrossRef]
- De-la-Torre, G.E. Microplastics: An emerging threat to food security and human health. J. Food Sci. Technol. 2020, 57, 1601–1608. [Google Scholar] [CrossRef]
- Di Guardo, G.; Mazzariol, S. Toxoplasma gondii: Clues from stranded dolphins. Vet. Pathol. 2013, 50, 737. [Google Scholar] [CrossRef]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H.; Grigg, M.E. Recent epidemiologic and clinical importance of Toxoplasma gondii infections in marine mammals: 2009–2020. Vet. Parasitol. 2020, 288, 109296. [Google Scholar] [CrossRef] [PubMed]
- Giorda, F.; Crociara, P.; Iulini, B.; Gazzuola, P.; Favole, A.; Goria, M.; Serracca, L.; Dondo, A.; Crescio, M.I.; Audino, T.; et al. Neuropathological Characterization of Dolphin Morbillivirus Infection in Cetaceans Stranded in Italy. Animals 2022, 12, 452. [Google Scholar] [CrossRef] [PubMed]
- Pintore, M.D.; Mignone, W.; Di Guardo, G.; Mazzariol, S.; Ballardini, M.; Florio, C.L.; Goria, M.; Romano, A.; Caracappa, S.; Giorda, F.; et al. Neuropathologic findings in cetaceans stranded in Italy (2002–2014). J. Wildl. Dis. 2018, 54, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Ahmadpour, E.; Rahimi, M.T.; Ghojoghi, A.; Rezaei, F.; Hatam-Nahavandi, K.; Oliveira, S.M.R.; Pereira, M.d.L.; Majidiani, H.; Siyadatpanah, A.; Elhamirad, S.; et al. Toxoplasma gondii Infection in Marine Animal Species, as a Potential Source of Food Contamination: A Systematic Review and Meta-Analysis. Acta Parasit. 2022, 67, 592–605. [Google Scholar] [CrossRef]
- Dumètre, A.; Dardé, M.L. How to detect Toxoplasma gondii oocysts in environmental samples? FEMS Microbiol. Rev. 2003, 27, 651–661. [Google Scholar] [CrossRef]
- Marangi, M.; Lago, N.; Mancinelli, G.; Antonio, O.L.; Scirocco, T.; Sinigaglia, M.; Specchiulli, A.; Cilenti, L. Occurrence of the protozoan parasites Toxoplasma gondii and Cyclospora cayetanensis in the invasive Atlantic blue crab Callinectes sapidus from the Lesina Lagoon (SE Italy). Mar. Pollut. Bull. 2022, 176, 113428. [Google Scholar] [CrossRef]
- Marangi, M.; Giangaspero, A.; Lacasella, V.; Lonigro, A.; Gasser, R.B. Multiplex PCR for the detection and quantification of zoonotic taxa of Giardia, Cryptosporidium and Toxoplasma in wastewater and mussels. Mol. Cell. Probes 2015, 29, 122–125. [Google Scholar] [CrossRef]
- Santoro, M.; Viscardi, M.; Boccia, F.; Borriello, G.; Lucibelli, M.G.; Auriemma, C.; Anastasio, A.; Veneziano, V.; Galiero, G.; Baldi, L.; et al. Parasite Load and STRs Genotyping of Toxoplasma gondii Isolates from Mediterranean Mussels (Mytilus galloprovincialis) in Southern Italy. Front. Microbiol. 2020, 11, 355. Available online: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00355 (accessed on 8 August 2023). [CrossRef]
- Putignani, L.; Mancinelli, L.; Del Chierico, F.; Menichella, D.; Adlerstein, D.; Angelici, M.; Marangi, M.; Berrilli, F.; Caffara, M.; di Regalbono, D.F.; et al. Investigation of Toxoplasma gondii presence in farmed shellfish by nested-PCR and real-time PCR fluorescent amplicon generation assay (FLAG). Exp. Parasitol. 2011, 127, 409–417. [Google Scholar] [CrossRef]
- Di Guardo, G.; Proietto, U.; Di Francesco, C.E.; Marsilio, F.; Zaccaroni, A.; Scaravelli, D.; Mignone, W.; Garibaldi, F.; Kennedy, S.; Forster, F.; et al. Cerebral Toxoplasmosis in Striped Dolphins (Stenella coeruleoalba) Stranded Along the Ligurian Sea Coast of Italy. Vet. Pathol. 2010, 47, 245–253. [Google Scholar] [CrossRef]
- Di Guardo, G.; Di Cesare, A.; Otranto, D.; Casalone, C.; Iulini, B.; Mignone, W.; Tittarelli, C.; Meloni, S.; Castagna, G.; Forster, F.; et al. Genotyping of Toxoplasma gondii isolates in meningo-encephalitis affected striped dolphins (Stenella coeruleoalba) from Italy. Vet. Parasitol. 2011, 183, 31–36. [Google Scholar] [CrossRef]
- Profeta, F.; Di Francesco, C.E.; Marsilio, F.; Mignone, W.; Di Nocera, F.; De Carlo, E.; Lucifora, G.; Pietroluongo, G.; Baffoni, M.; Cocumelli, C.; et al. Retrospective seroepidemiological investigations against Morbillivirus, Toxoplasma gondii and Brucella spp. in cetaceans stranded along the Italian coastline (1998–2014). Res. Vet. Sci. 2015, 101, 89–92. [Google Scholar] [CrossRef]
- Mazzariol, S.; Marcer, F.; Mignone, W.; Serracca, L.; Goria, M.; Marsili, L.; Di Guardo, G.; Casalone, C. Dolphin Morbillivirus and Toxoplasma gondii coinfection in a Mediterranean fin whale (Balaenoptera physalus). BMC Vet. Res. 2012, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Marcer, F.; Marchiori, E.; Centelleghe, C.; Ajzenberg, D.; Gustinelli, A.; Meroni, V.; Mazzariol, S. Parasitological and pathological findings in fin whales Balaenoptera physalus stranded along Italian coastlines. Dis. Aquat. Organ. 2019, 133, 25–37. [Google Scholar] [CrossRef]
- Fernández-Escobar, M.; Giorda, F.; Mattioda, V.; Audino, T.; Di Nocera, F.; Lucifora, G.; Varello, K.; Grattarola, C.; Ortega-Mora, L.M.; Casalone, C.; et al. Toxoplasma gondii Genetic Diversity in Mediterranean Dolphins. Pathogens 2022, 11, 909. [Google Scholar] [CrossRef] [PubMed]
- Petrella, A.; Mazzariol, S.; Padalino, I.; Di Francesco, G.; Casalone, C.; Grattarola, C.; Di Guardo, G.; Smoglica, C.; Centelleghe, C.; Gili, C. Cetacean Morbillivirus and Toxoplasma gondii Co-infection in Mediterranean Monk Seal Pup, Italy. Emerg. Infect. Dis. 2021, 27, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Heim, R.R. An overview of weather and climate extremes—Products and trends. Weather. Clim. Extrem. 2015, 10, 1–9. [Google Scholar] [CrossRef]
- Jones, J.L.; Dargelas, V.; Roberts, J.; Press, C.; Remington, J.S.; Montoya, J.G. Risk factors for Toxoplasma gondii infection in the United States. Clin. Infect. Dis. 2009, 49, 878–884. [Google Scholar] [CrossRef]
- Grattarola, C.; Giorda, F.; Iulini, B.; Pintore, M.D.; Pautasso, A.; Zoppi, S.; Goria, M.; Romano, A.; Peletto, S.; Varello, K.; et al. Meningoencephalitis and Listeria monocytogenes, Toxoplasma gondii and Brucella spp. coinfection in a dolphin in Italy. Dis. Aquat. Organ. 2016, 118, 169–174. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez-Fernandez, V.; Bruschi, F. Toxoplasma gondii in Marine Life of Italian Coasts, What Do We Know So Far? Parasitologia 2023, 3, 364-373. https://doi.org/10.3390/parasitologia3040035
Rodriguez-Fernandez V, Bruschi F. Toxoplasma gondii in Marine Life of Italian Coasts, What Do We Know So Far? Parasitologia. 2023; 3(4):364-373. https://doi.org/10.3390/parasitologia3040035
Chicago/Turabian StyleRodriguez-Fernandez, Veronica, and Fabrizio Bruschi. 2023. "Toxoplasma gondii in Marine Life of Italian Coasts, What Do We Know So Far?" Parasitologia 3, no. 4: 364-373. https://doi.org/10.3390/parasitologia3040035