1. Introduction
The brown ear tick,
Rhipicephalus appendiculatus, is the primary vector of the apicomplexan parasite,
Theileria parva, which is the causative agent of East Coast fever (ECF) in cattle, and of Corridor Disease (CD) in buffalo [
1].
Rhipicephalus appendiculatus is a hard tick with a three-host life cycle [
2]. Adults mainly attach to the ears of domestic and wild ruminants, whereas ticks in the immature stages (larvae and nymphs) attach to the ears, head, and neck of most ruminant species, as well as carnivores, equids, and hares [
3]. Their geographical distribution is widespread in eastern, central, and southern Africa, where the climatic conditions are suitable for their seasonal activity [
4,
5]. For instance, larvae and nymphs of
R. appendiculatus prefer the dry and warm seasons, while adults prefer the more wet and rainy seasons [
6].
East Coast fever is one of the most lethal tick-borne diseases of cattle in sub-Saharan Africa. It is a severe lympho-proliferative disease caused by an acute infection of
T. parva; however, it remains asymptomatic in wild buffalos [
7]. Kalume et al. showed that mortality can reach up to 100% in exotic cattle, which is a constraining factor for improving sustainable livestock production, the pillar of durable development in African countries, especially for subsistence farming [
4]. Currently, the most effective method to protect cattle against acute disease caused by
T. parva, is the vaccination-based infection and treatment method (ITM) [
8]. This involves inoculating cattle with live sporozoites and simultaneously treating with long-acting oxytetracyclines to control the severity of the disease [
8]. The production of this vaccine requires many animals to be utilized during various steps: in the first step, cattle are used to generate
T. parva-infected ticks by feeding nymphs on infected cattle (pick up phase). Engorged nymphs are incubated to molt to adults. Step two, donor rabbits are used to partially feed the infected molted adult ticks [
9]. The partial blood feeding (forced interrupted feeding on day 4 after tick attachment on rabbit) allows for the maturation of sporoblasts into sporozoites in the salivary glands of infected
R. appendiculatus adults, and prevents loss of sporozoites [
9].
Several artificial feeding systems have been used to feed arthropods, with different adaptations for hard/soft ticks, mosquitoes, and sand flies thus avoiding the use of experimental animals [
10,
11,
12]. In fact, the development and standardization of laboratory methods to feed arthropods artificially is desirable, since these “in vitro” assays not only contribute to the objectives of the 3R (Reduction, Refinement and Replacement) principles in animal experimentation [
13], but also provide researchers with a rapid and inexpensive tool to investigate the transmission of tick-borne pathogens. These methods will also aid in testing the effectiveness of anti-tick vaccines and topical or systemic acaricides [
14].
For ixodid ticks, two artificial devices have been developed: membrane-based systems and capillary tube feeding. For instance, Purnell and Joyner (1967) successfully infected
R. appendiculatus ticks with
T. parva by fitting capillary tubes filled with infected blood loosely over the ticks’ mouthparts. Furthermore, a goldbeater’s skin was successfully used for feeding
R. appendiculatus until repletion [
15,
16]. In 1996, Young et al. used artificial feeding to study pathogenic transmission of
T. parva and
Ehrlichia ruminantium for
R. appendiculatus and
Amblyomma variegatum ticks, respectively. They used a modified goldbeater’s skin and the skin of a rabbit or bovine as feeding membranes, depending on the length of the tick’s mouthparts [
17]. Furthermore, Musyoki et al. compared the transmission of
T. parva by
R. appendiculatus nymphs by feeding them in vitro and in vivo. Consequently, parameters such as the mean attachment rate, feeding duration, and engorgement weights of membrane-fed ticks were not significantly different from nymphs fed on cattle [
18]. In another study [
19], the effect of the blood anticoagulant in the feeding process of
A. variegatum was assessed using defibrinated, heparinized or Ethylenediaminetetraacetic acid (EDTA) treated blood. The results showed that the feeding process was disturbed or completely stopped after 48h for ticks feeding on blood containing EDTA. Thus, the anticoagulant type could influence the feeding process of hematophagous arthropod vectors [
20] within an in vitro feeding system.
This study assessed the attachment and feeding of adult R. appendiculatus ticks, using an in vitro tick feeding system by comparing the impact of different blood anticoagulating factors on the feeding process.
2. Results
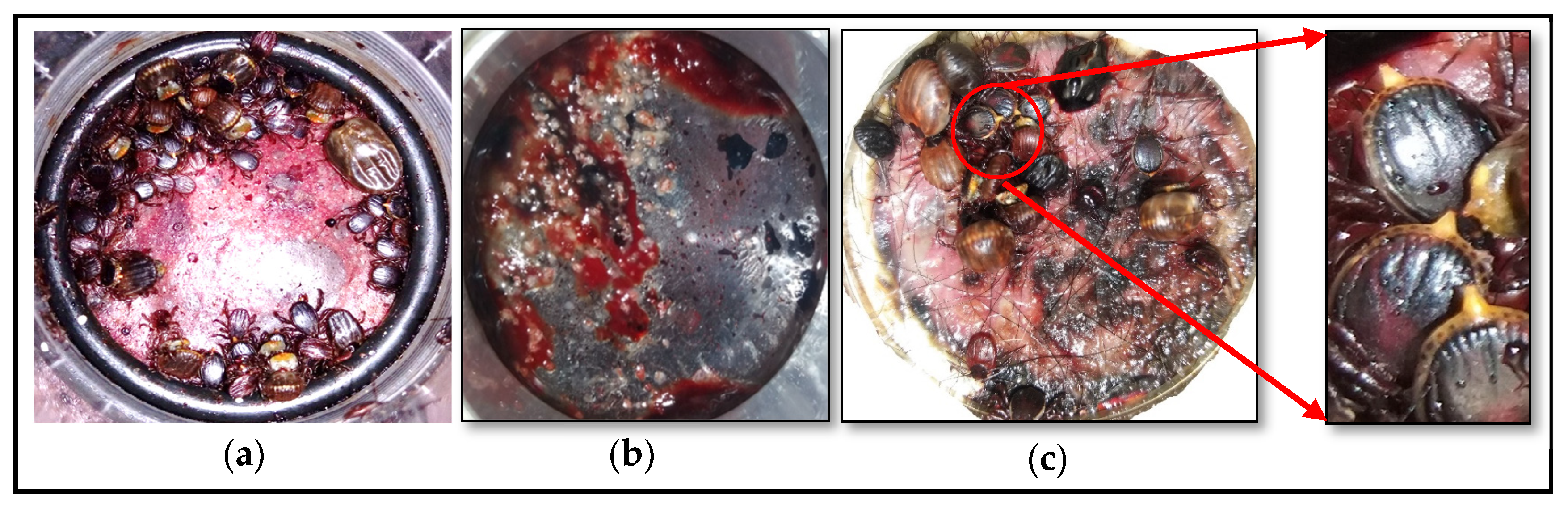
During artificial in vitro feeding, ticks retained normal behavior, such as movement and host-seeking on the membrane. The first attached ticks were observed in all feeders, within the first 12 h after adding the ticks to the tick receptacle. Cement formation was observed at the tick attachment sites after 24 h on the blood receptacle side of the membrane, indicating tick mouthpart insertion through, and tick attachment to, the membrane (
Figure 1b). Upon first tick attachment to the feeding membrane, the remaining ticks started to attach to the same attachment area, forming aggregates similar to their behaviour on animal hosts (
Figure 1a). The attachment rate 4 days after adding the ticks to the tick receptacle ranged from 76.7% to 83.3% (24.0 ± 1.0), 68.0% to 86.7% (45.5 ± 4.8), and 67.0% to 77.0% (42.5 ± 2.17) for lithium heparin, citrate phosphate dextrose, and defibrinated blood, respectively (
Table 1). No significant difference (χ2 = 0.74;
p = 0.69) were observed between the three blood anticoagulant methods.
The start of tick engorgement, which was assessed by the presence of feces, as well as the extension of caudal appendage for males (
Figure 1c), was noted in all feeders within 24 to 48 h after adding the ticks to the tick receptacle.
Moreover, feeding until repletion occurred in ticks that fed on heparinized and defibrinated blood. Ticks showed normal feeding behavior with the initial slow-feeding phase followed by the rapid-feeding phase that occurred during the final 12 to 36 h after adding the ticks to the tick receptacle, whereafter the ticks started to detach. The female ticks fed to repletion within 7 to 10 days after adding the ticks to the tick receptacle. In the feeders that contained citrate phosphate, the feeding progress to the fast-feeding phase seemed to be interrupted, since the ticks remained in a semi-engorged status and not feeding to full repletion.
No dead ticks were observed for any of the feeders during the first 48h after tick placement in the tick receptacle. However, after Day 3, some non-attached ticks were found dead (less than 5%). In some feeders, blood leakage occurred on Day 2 or 3 after starting the system. This could have been due to the thickness of the membrane being too thin and/or the pressure exerted by the ticks on the membrane when feeding in clusters.
3. Discussion
In the present study, we investigated an in vitro tick feeding system designed to overcome the limitations encountered with previous models. During the challenge period, the temperature of the blood meal was kept stable at 37 °C, which is ideal for adult
R. appendiculatus feeding. It has been reported that variation in the feeding medium’s (blood) temperature could affect tick attachment and the feeding process of
R. appendiculatus, resulting in stopping their blood feeding or even detachment from the membrane [
15]. The position of the blood above the ticks and the maintenance of a stable blood temperature, as well as the delivery of blood meal without interrupting the blood-feeding process, are improvements that were achieved by this system. In addition, the system disclosed in this study has a blood vessel feeding device with an inlet and outlet that can be connected to a peristaltic pump, either for circulating the blood or to collect blood samples in order to screen for pathogens injected by infected ticks.
During this study, ticks were reactivated by incubating them for at least one week at room temperature before using them in the experiment. It was reported that when
R. appendiculatus ticks were exposed to short daylight periods, they could enter a diapause state [
21], resulting in non-attachment of ticks on both artificial membranes and rabbits. Exposing ticks to daylight periods is one of the primary factors that ensures the activation of ticks to come out of the diapause state.
At the same time, keeping ticks in a 14-h daylight cycle for a short period allows for their activation, and then improves their attachment to the host [
21]. In our study, the willingness of ticks to attach and feed was adapted by adjusting the conditions for feeding to be optimal. Cattle hair was added onto the membrane during membrane preparation to simulate a cattle smell, as semiochemical conditions attract ticks. The results obtained by [
15] showed excellent attachment to a goldbeater’s skin membrane treated with cattle ear wash and tick feces compared to a membrane without any treatment. Additionally, the temperature and humidity were controlled as physical stimuli, using an incubator. Finally, both conditions (tick attractant stimuli and ideal temperature and humidity conditions) served to mimic natural conditions. Results reported here highlight the efficiency of using goldbeater’s skin reinforced with silicone in an artificial feeding system that is generally similar to the old goldbeater’s skin approach described by Waladde et al. [
15].
In this study, we investigated an in vitro tick feeding system designed to overcome the limitations encountered with previous models. During the experimental period, the temperature of the blood used in the feeding system was kept constant at 37 °C, which is ideal for the feeding of adult
R. appendiculatus ticks attached to the artificial feeding membrane. It has been reported that variation in the feeding medium’s (blood) temperature could affect tick attachment and the feeding process of
R. appendiculatus ticks, resulting in the ticks terminating the feeding process or even detaching from the membrane [
15]. The position of the blood above the ticks and the maintenance of a constant blood temperature, as well as the delivery of blood meal without interrupting the blood-feeding process, are improvements that were achieved by this in vitro feeding system. In addition, the system disclosed in this study has a blood vessel feeding device with an inlet and outlet that can be connected to a peristaltic pump, either for circulating the blood or to collect blood samples in order to screen for pathogens injected by infected ticks.
Noteworthy, is that a high attachment rate was recorded after four days post-tick seeding. By this time, tick attachment was greater than 70%, regardless of the blood anticoagulant used. In fact, this period corresponds to the duration of maturation of sporoblasts into sporozoites of
T. parva in the salivary gland acini of infected adult
R. appendiculatus ticks [
22]. Therefore, artificial feeding of
T. parva-infected-ticks could be interrupted at this stage to collect sporozoites for vaccine preparations. As recently reported, the isolation of viable and infectious
T. parva sporozoites by feeding infected adult ticks on an in vitro tick feeding system was successfully achieved [
23].
The tick engorgement process began 24 h after placing ticks in the tick receptacle. This observation is similar to the results previously reported [
15] in which,
R. appendiculatus ticks started feeding on blood 12 to 48 h after seeding using defibrinated blood and a goldbeater’s skin. Defibrinated or anticoagulated bovine blood was used to formulate the tick blood meal in order to prevent blood clotting during the attachment and feeding process. Feeding success of
R. appendiculatus until repletion was greater when the ticks were fed on heparinized and defibrinated cattle blood rather than when other anticoagulants were used.
4. Materials and Methods
4.1. In Vitro Tick Feeding System
The in vitro tick feeding system used in the present study was described previously in [
23,
24]. Briefly, each feeder consisted of five parts (
Figure 2a,b): (1) a blood heating element, (2) blood vessel, (3) membrane frame, (4) connector, and (5) tick receptacle. Each feeder was connected to the system’s adjustable digital heating device, allowing the blood to be maintained at constant set temperature (
Figure 2c).
4.2. Ticks
Ticks used in this experiment were originally collected in the field in Uganda (Serere) in 2016 and obtained and cycled at the Clinvet Morocco facility. The laboratory-reared ticks were pathogen-free, maintained by routine passage on rabbits from larvae to adult. All stages were maintained at 18 ± 2 °C and a relative humidity (RH) of ≥60%. The adult ticks used in this study were at least two weeks of age after molting. To increase their susceptibility to attaching and feeding, the ticks were transferred to conditions simulating ideal environmental conditions with 25 ± 2 °C, RH ≥ 60%, and 14 h of daylight for at least one week before the beginning of the experiment.
4.3. Membrane and Feeder Preparation
The in vitro tick feeding system was prepared based on a method described previously [
23]. Briefly, the feeding units were made of five pieces connected to each other (
Figure 2a,b). The tick receptacle (listed as number 5 in
Figure 2b and containing ticks) was separated from the blood receptacle (listed as number 2 in
Figure 2b and containing blood) by a previously prepared silicone-based membrane that was gently fixed to the membrane frame. Feeding membranes consisted of goldbeater’s skin originally made from bovine intestine (Preservation Equipment Ltd., Norfolk, UK) with a thickness of 30 μm, which were treated with a thin layer of silicone rubber mixture (Smooth-On, Inc., East Texas, PA, USA) to improve the softness, resulting in a final membrane thickness of 100–120 μm. The silicone mixture was prepared using 5 g of each Ecoflex Supersoft 00-50 silicone A and B (Smooth-On, Easton, PA, USA) mixed with 2 mL of hexane (Sigma-Aldrich, St. Louis, MO, USA). Just after adding the silicone mixture, bovine hair was added to the membrane to act as a tick attractant that aided in tick attachment. The membrane was allowed to polymerize overnight at room temperature and was then measured for thickness.
4.4. Blood Preparation
Blood was collected from healthy donor cattle using two different anticoagulants: lithium heparin tubes and blood collection bags supplemented with citrate phosphate dextrose (CPD) (MacoPharma, Mouvaux, France). In addition, blood that had been defibrinated mechanically using a spatula was also used. All the blood was supplemented with 0.2% glucose (Sigma-Aldrich, St. Louis, MO, USA) and stored at 4 °C for a maximum of 15 days during its use. Prior to adding the blood to the blood receptacle, it was heated to 37 °C and treated with 0.5 µL/mL of gentamycin (10 mg/mL, Sigma-Aldrich). A volume of 10 mL of blood/feeder was then added to each blood receptacle. The addition of antibiotics to the blood and washing of the blood receptacle systematically during every blood change (every 12 h) was carried out to avoid contamination of the blood.
4.5. Tick Incubation and Engorgement Assessment
In total, 8 feeders were used with 60 ticks per feeder (sex ratio 1:1) using defibrinated blood (4 feeders) and citrate phosphate dextrose (4 feeders) anticoagulant blood. In addition, 2 feeders were used with 30 ticks per feeder for heparinized blood. The availability of ticks did not allow for the usage of more feeders for heparinized blood. The whole system was placed inside an incubator at 27 ± 2 °C with a RH ≥ 60% and a 14 h light/10 h dark photoperiod. The system was assessed twice a day during blood changes (every 12 h). To remove the blood residues before adding new blood, each feeder was rinsed using sterile 0.9% NaCl heated to 37 °C. Tick assessment (attachment, mortality, engorgement, and presence of feces) was performed during every blood change.
4.6. Data Analysis
Data were analyzed using Excel to calculate the means, percentages, and standard deviations. Furthermore, Pearson’s chi-square test was performed to analyze the attachment rates between the three types of blood collection methods. p-values < 0.05 were considered statistically significant.


 ,
, 


{kind=link}
{kind=link}