
Population-Inclusive Assigned-Sex-at-Birth Estimation from Skull Computed Tomography Scans


Abstract
:1. Introduction
1.1. The Role of Ancestry in the Biological Profile and Study Rationale
1.2. The Study Collection: New Mexico Decedent Image Database
1.3. Assigned Skeletal Sex Estimation
1.4. Methods for Estimating Assigned Sex
2. Materials and Methods
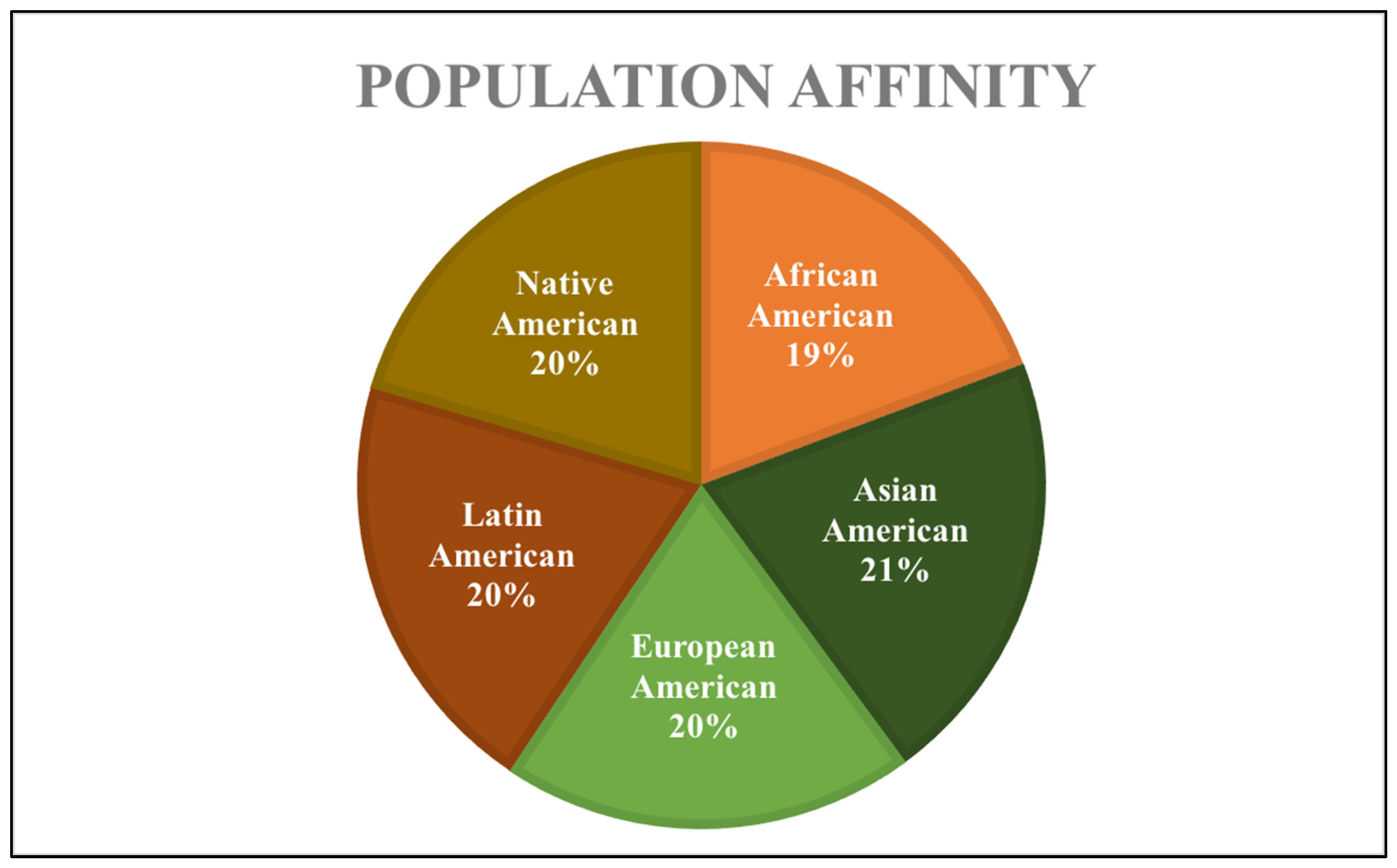
2.1. Study Sample
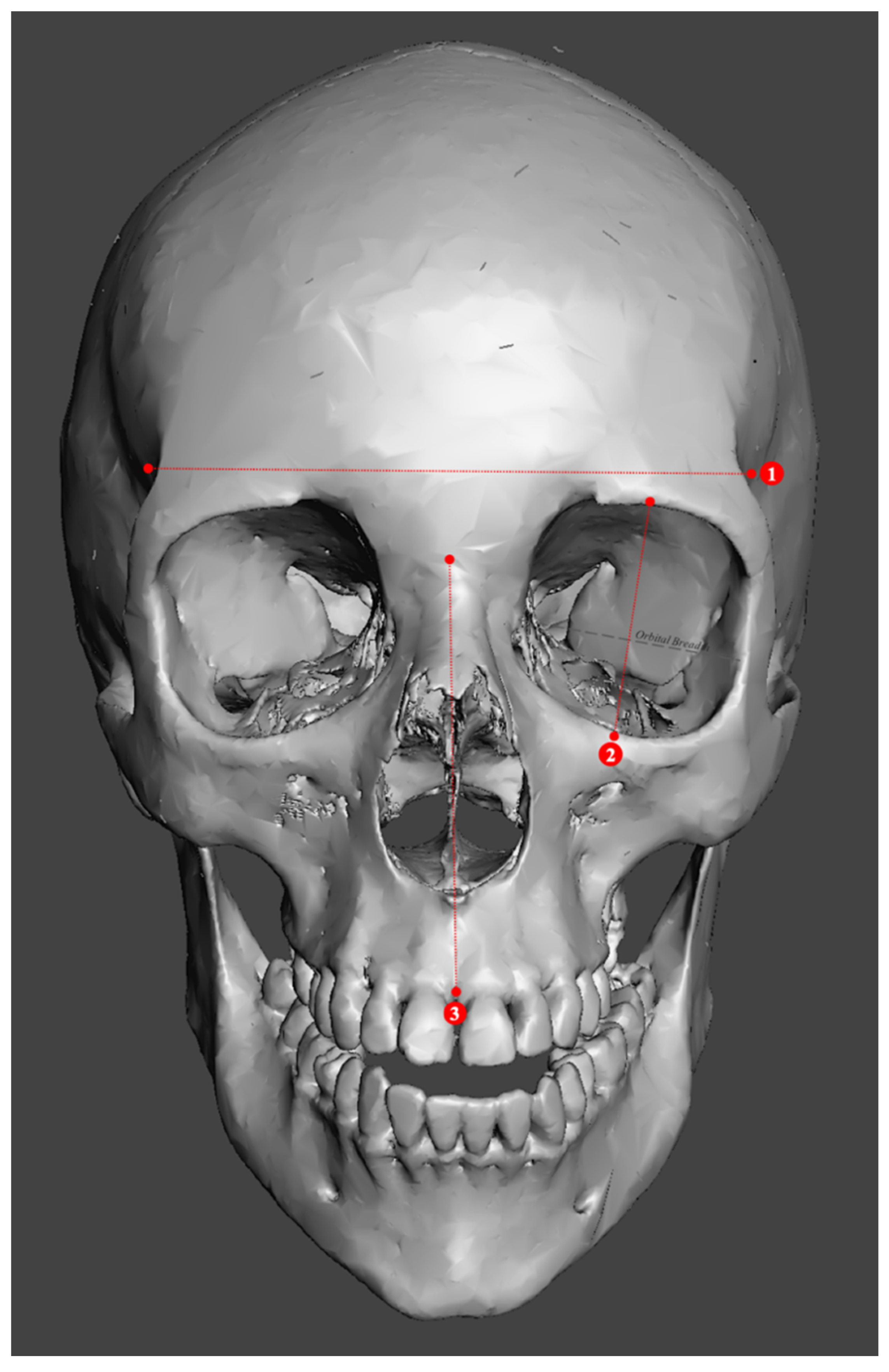
2.2. Study Sample Preparation and Data Collection
2.3. Statistical Analyses
3. Results
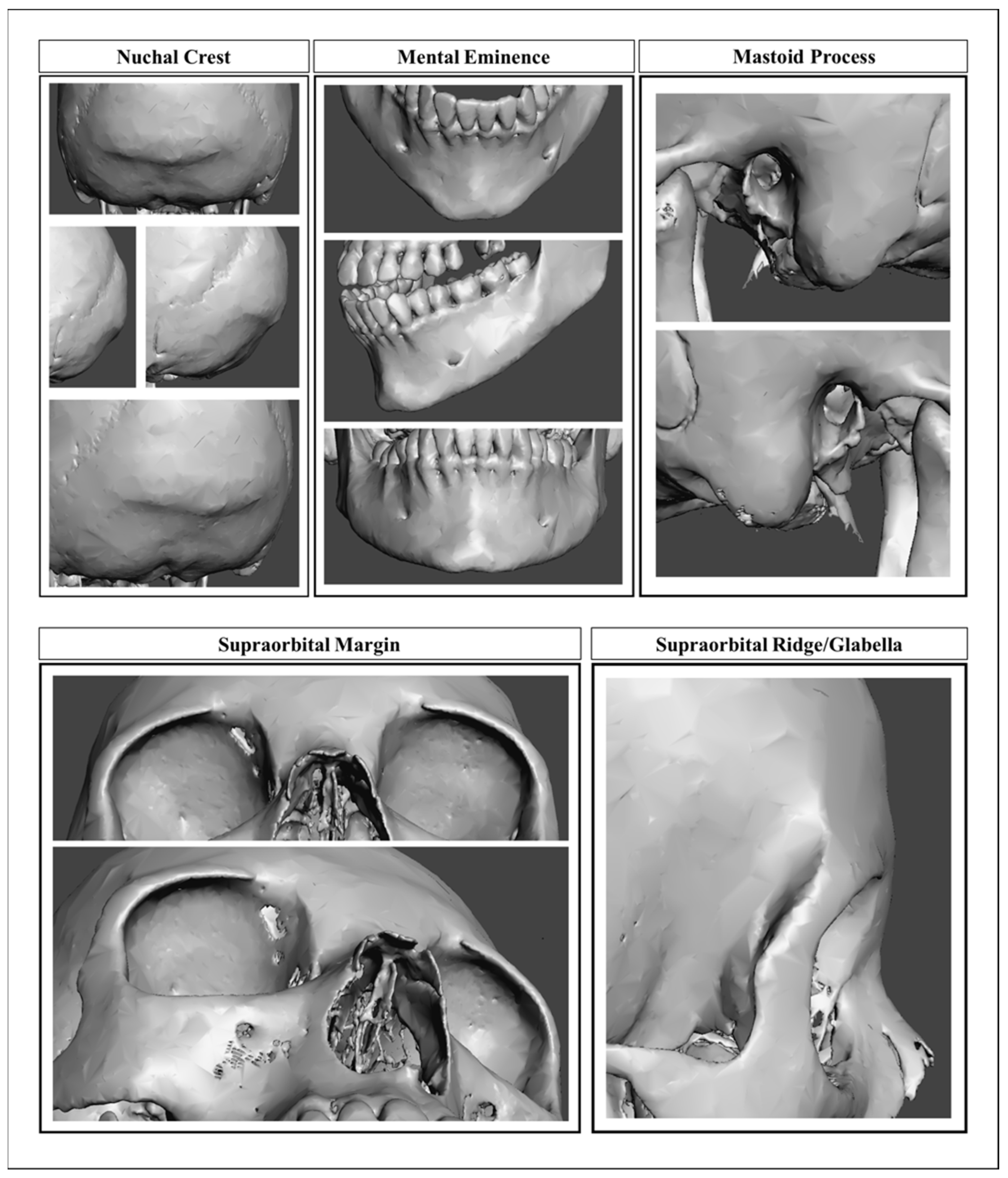
3.1. Nonmetric Models
3.2. Metric Models
3.3. Mixed Model
3.4. Intrarater Reliability
4. Discussion
4.1. Nonmetric Models
4.2. Metric Models
4.3. Mixed Model
4.4. Intrarater Reliability
4.5. Data Collection from 3D-VR CT Images
4.6. Sex and Gender
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klales, A.R. Sex Estimation of the Human Skeleton: History, Methods, and Emerging Techniques, 1st ed.; Elsevier Press: Waltham, MA, USA, 2020; pp. 1–364. [Google Scholar]
- Tallman, S.D. Cranial nonmetric sexual dimorphism and sex estimation in east and southeast Asian individuals. Forensic Anthropol. 2019, 2, 204–221. [Google Scholar] [CrossRef]
- Ubelaker, D.H.; DeGaglia, C.M. Population variation in skeletal sexual dimorphism. Forensic Sci. Int. 2017, 278, 407.e1–407.e7. [Google Scholar] [CrossRef]
- Acsádi, G.; Nemeskéri, J. History of Human Life Span and Mortality; Akadémiai Kiadó: Budapest, Hungary, 1970. [Google Scholar]
- Broca, P. Instructions Craniologiques et Craniométriques; Series 2; Society of Anthropology of Paris: Paris, France, 1875. [Google Scholar]
- Buikstra, J.E.; Ubelaker, D.H. Standards for Data Collection from Human Skeletal Remains; Research Series No. 44; Arkansas Archaeological Survey: Fayetteville, NC, USA, 1994. [Google Scholar]
- Dayal, M.R.; Spocter, M.A.; Bidmos, M.A. An assessment of sex using the skull of black South Africans by discriminant function analysis. Homo 2008, 59, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Garvin, H.M.; Sholts, S.B.; Mosca, L.A. Sexual dimorphism in human cranial trait scores: Effects of population, age, and body size. Am. J. Phys. Anthropol. 2014, 154, 259–269. [Google Scholar] [CrossRef]
- Graw, M.; Czarnetzki, A.; Haffner, H.T. The form of the supraorbital margin as a criterion in identification of sex from the skull: Investigations based on modern human skulls. Am. J. Phys. Anthropol. 1999, 108, 91–96. [Google Scholar] [CrossRef]
- Krogman, W.M. The skeleton in forensic medicine. Postgrad. Med. J. 1955, 17, A48–A62. [Google Scholar]
- Krogman, W.M.; Işcan, M.Y. The Human Skeleton in Forensic Medicine; Charles C Thomas: Springfield, IL, USA, 1986. [Google Scholar]
- Lewis, C.J.; Garvin, H.M. Reliability of the Walker cranial nonmetric method of implications for sex estimation. J. Forensic Sci. 2016, 61, 743–751. [Google Scholar] [CrossRef]
- Martin, R.; Knussman, R. Anthropologie: Handbuch der Vergleichenden Biologie des Menschen; Gustav Fisher Verlag: Stuttgart, Germany, 1988. [Google Scholar]
- Rogers, T.L. Determining the sex of human remains through cranial morphology. J. Forensic Sci. 2005, 50, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Spradley, M.K.; Jantz, R.L. Sex estimation in forensic anthropology: Skull versus postcranial elements. J. Forensic Sci. 2011, 56, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Tallman, S.D.; Go, M.C. Application of the optimized summed scored attributes method to sex estimation in Asian crania. J. Forensic Sci. 2018, 63, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.L. Sexing skulls using discriminant function analysis of visually assessed traits. Am. J. Phys. Anthropol. 2008, 136, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Walrath, D.E.; Turner, P.; Bruzek, J. Reliability test of the visual assessment of cranial traits for sex determination. Am. J. Phys. Anthropol. 2004, 125, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Karsten, J.K. A test of the preauricular sulcus as an indicator of sex. Am. J. Phys. Anthropol. 2017, 165, 604–608. [Google Scholar] [CrossRef]
- Letterman, G.S. The greater sciatic notch in American Whites and Negroes. Am. J. Phys. Anthropol. 1941, 98, 59–72. [Google Scholar] [CrossRef]
- Patterson, M.M.; Tallman, S.D. Cranial and postcranial metric sex estimation in modern Thai and Ancient Native American individuals. Forensic Anthropol. 2019, 2, 233–252. [Google Scholar] [CrossRef]
- Phenice, T.W. A newly developed visual method of sexing the os pubis. Am. J. Phys. Anthropol. 1969, 30, 297–301. [Google Scholar] [CrossRef]
- Rogers, T.L. A visual method of determining the sex of the skeleton using the distal humerus. J. Forensic Sci. 1999, 44, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.L.; Flournoy, L.E.; McCormick, W.F. The rhomboid fossa of the clavicle as a sex and age estimator. J. Forensic Sci. 2000, 45, 61–67. [Google Scholar] [CrossRef]
- Tallman, S.D.; Blanton, A.I. Distal humerus morphological variation and sex estimation in modern Thai individuals. J. Forensic Sci. 2020, 65, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Vance, V.L.; Steyn, M.; L’Abbé, E.N.L. Nonmetric sex determination from the distal and posterior humerus in Black and white South Africans. J. Forensic Sci. 2011, 56, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Walker, P.L. Greater sciatic notch morphology: Sex, age, and population differences. Am. J. Phys. Anthropol. 2005, 127, 385–391. [Google Scholar] [CrossRef]
- Washburn, S.L. Sex difference in the pubic bone. Am. J. Phys. Anthropol. 1948, 6, 199–207. [Google Scholar] [CrossRef]
- Bethard, J.D.; DiGangi, E.A. Moving beyond a lost cause: Forensic anthropology and ancestry estimates in the United States. J. Forensic Sci. 2020, 65, 1791–1792. [Google Scholar] [CrossRef] [PubMed]
- DiGangi, E.A.; Bethard, J.D. Uncloaking a lost cause: Decolonizing ancestry estimation in the United States. Am. J. Phys. Anthropol. 2021, 175, 422–436. [Google Scholar] [CrossRef]
- Cunha, E.; van Vark, G. The construction of sex discriminant functions from a large collection of skulls of known sex. Int. J. Anthropol. 1991, 6, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Eleveth, P.B.; Tanner, J.M. Worldwide Variation in Human Growth, 2nd ed.; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Franklin, D.; Freedman, L.; Milne, N. Sexual dimorphism and discriminant function sexing in indigenous South African crania. Homo 2005, 55, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Kemkes, A.; Gobel, T. Metric assessment of the “mastoid triangle” for sex determination: A validation study. J. Forensic Sci. 2006, 51, 985–989. [Google Scholar] [CrossRef]
- Adams, D.M.; Pilloud, M.A. The (mis)appropriation of biological anthropology in race science and the implications for forensic anthropology. Forensic Anthropol. 2021, 4, 1–22. [Google Scholar] [CrossRef]
- Albanese, J. A method for estimating sex using the clavicle, humerus, radius, and ulna. J. Forensic Sci. 2013, 58, 1413–1419. [Google Scholar] [CrossRef] [PubMed]
- Albanese, J.; Saunders, S.R. Is it possible to escape racial typology in forensic identification? In Forensic Anthropology and Medicine: Complementary Sciences from Recovery to Cause of Death; Schmitt, A., Cunha, E., Pinheiro, J., Eds.; Humana Press: Totowa, NJ, USA, 2006; pp. 281–316. [Google Scholar]
- Albanese, J.; Eklics, G.; Tuck, A. A metric method for sex determination using the proximal femur and fragmentary hipbone. J. Forensic Sci. 2008, 53, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Blakey, M.L. Understanding racism in physical (biological) anthropology. Am. J. Phys. Anthropol. 2021, 175, 316–325. [Google Scholar] [CrossRef]
- Carson, E.A. Maximum likelihood estimation of human craniometrics heritabilities. Am. J. Phys. Anthropol. 2006, 131, 169–180. [Google Scholar] [CrossRef]
- Edgar, H.J.H. Population structure, population, heterogeneity, and sources of error in the forensic estimation of “race”. In Proceedings of the 72nd Annual Scientific Meeting of the American Academy of Forensic Sciences, Anaheim, CA, USA, 17–22 February 2020. [Google Scholar]
- Moss, J.L. The forgotten victims of missing white woman syndrome: An examination of legal measures that contribute to the lack of search and recovery of missing black girls and women. Race Gender Soc. Just. 2018, 25, 737–762. [Google Scholar]
- Ross, A.H.; Pilloud, M. The need to incorporate human variation and evolutionary theory in forensic anthropology: A call for reform. Am. J. Phys. Anthropol. 2021, 176, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Sommers, Z. Missing white woman syndrome: An empirical analysis of race and gender disparities in online news coverage of missing persons. J. Crim. Law Criminol. 2016, 106, 275–314. [Google Scholar]
- Spradley, K.; Jantz, R.L. What are we really estimating in forensic anthropological practice, population affinity or ancestry? Forensic Anthropol. 2021, 4, 171–180. [Google Scholar] [CrossRef]
- Tallman, S.D.; Parr, N.M.; Winburn, A.P. Assumed differences; unquestioned typologies: The oversimplification of race and ancestry in forensic anthropology. Forensic Anthropol. 2021, 4, 73–96. [Google Scholar] [CrossRef]
- Albanese, J.; Tuck, A.; Gomes, J.; Cardoso, H.F.V. An alternative approach for estimating stature from long bones that is not population-or group-specific. Forensic Sci. Int. 2016, 259, 59–68. [Google Scholar] [CrossRef]
- Edgar, H.J.H.; Daneshvari Verry, S.; Moes, E.; Adolphi, N.L.; Bridges, P.; Nolte, K.B. New Mexico Decedent Image Database; Office of the Medical Investigator, University of New Mexico: Albuquerque, NM, USA, 2020. [Google Scholar] [CrossRef]
- Berry, S.R. Metadata Determination for Cadaveric Collection. Master’s Thesis, University of New Mexico, Albuquerque, NM, USA, 2014. [Google Scholar]
- Daneshvari Berry, S.; Edgar, H.J.H. Development of a large-scale, whole body CT image database. In Proceedings of the AMIA Annual Symposium, Washington, DC, USA, 6–8 November 2017. [Google Scholar]
- Daneshvari Berry, S.; Edgar, H.J.H. Announcement: The New Mexico decedent image database. Forensic Imaging 2021, 24, 1–3. [Google Scholar]
- Tise, M.L.; Spradley, M.K.; Anderson, B.E. Postcranial sex estimation of individuals considered Hispanic. J. Forensic Sci. 2013, 58 (Suppl. S1), S9–S14. [Google Scholar] [CrossRef] [PubMed]
- GLAAD. Available online: https://www.glaad.org/reference/transgender (accessed on 16 September 2021).
- Blackless, M.; Charuvastra, A.; Derryck, A.; Fausto-Sterling, A.; Lauzanne, K.; Lee, E. How sexually dimorphic are we? Review and synthesis. Am. J. Hum. Biol. 2000, 12, 151–156. [Google Scholar] [CrossRef]
- Stock, M.K. A preliminary analysis of the age of full expression of sexually dimorphic cranial traits. J. Forensic Sci. 2018, 63, 1802–1808. [Google Scholar] [CrossRef]
- Vanderschueren, D.; Vandenput, L.; Boonen, S.; Lindberg, M.L.; Bouillon, R.; Ohlsson, C. Androgens and bone. Endocr. Rev. 2004, 25, 389–425. [Google Scholar] [CrossRef] [Green Version]
- Bouillon, R.; Bex, M.; Vanderschueren, D.; Boonen, S. Estrogens are essential for male pubertal periosteal bone expansion. J. Clin. Endocrinol. Metab. 2004, 89, 6025–6029. [Google Scholar] [CrossRef] [Green Version]
- Carson, J.A.; Manolagas, S.C. Effects of sex steroids on bones and muscles: Similarities, parallels, and putative interactions in health and disease. Bone 2015, 80, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Vanderschueren, D.; Venken, K.; Ophoff, J.; Boullon, R.; Boonen, S. Sex steroids and the periosteum-reconsidering the roles of androgens and estrogens in periosteal expansion. J. Clin. Endocrinol. Metab. 2006, 91, 378–382. [Google Scholar] [CrossRef] [Green Version]
- Saggese, G.; Baroncelli, G.I.; Bertelloni, S. Puberty and bone development. Best Pract. Res. Clin. Endocrinol. Metab. 2002, 16, 53–64. [Google Scholar] [CrossRef]
- Arsuaga, J.L.; Carretero, J.M. Multivariate analysis of the sexual dimorphism of the hip bone in modern human population and in early hominids. Am. J. Phys. Anthropol. 1994, 93, 241–257. [Google Scholar] [CrossRef]
- Best, K.C.; Garvin, M.S.; Cabo, L.L. An investigation into the relationship between human cranial and pelvic sexual dimorphism. J. Forensic Sci. 2018, 63, 990–1000. [Google Scholar] [CrossRef]
- Frayer, D.; Wolpoff, M. Sexual dimorphism. Annu. Rev. Anthropol. 1985, 14, 429–473. [Google Scholar] [CrossRef]
- Scheuer, L.; Black, S. The Juvenile Skeleton, 1st ed.; Academic Press: London, UK, 2004; p. 140. [Google Scholar]
- Klales, A.R. Current practices in physical anthropology for sex estimation in unidentified, adult individuals. Am. J. Phys. Anthropol. 2013, 150, 168. [Google Scholar]
- Klales, A.R.; Ousley, S.D.; Vollner, J.M. A revised method of sexing the human innominate using Phenice’s nonmetric traits and statistical methods. Am. J. Phys. Anthropol. 2012, 149, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Stewart, T. Essentials for Forensic Anthropology; Charles C Thomas: Springfield, IL, USA, 1979. [Google Scholar]
- Curate, F.; Coelho, J.; Gonçalves, D.; Coelho, C.; Ferreira, M.T.; Navega, D.; Cunha, E. A method for sex estimation using the proximal femur. Forensic Sci. Int. 2016, 266, 579.e1–579.e7. [Google Scholar] [CrossRef] [Green Version]
- Bass, W.M. Human Osteology: A Laboratory and Field Manual, 5th ed.; Missouri Archaeological Society: Columbia, MO, USA, 2005. [Google Scholar]
- Garvin, H.M.; Ruff, C.B. Sexual dimorphism in skeletal browridge and chin morphologies determined using a new quantitative method. Am. J. Phys. Anthropol. 2012, 147, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Tallman, S.D.; Kincer, C.D.; Plemons, E.D. Centering transgender individuals in forensic anthropology and expanding binary sex estimation in casework and research. Forensic Anthropol. 2022, 5, 161–180. [Google Scholar] [CrossRef]
- Jantz, R.; Ousley, S. Fordisc 3: Third generation of computer-aided forensic anthropology. Rechtsmedizin 2013, 23, 97–99. [Google Scholar]
- Burns, K.R. Forensic Anthropology Training Manual, 2nd ed.; Pearson: New York, NY, USA, 2006. [Google Scholar]
- Gray, H. Gray’s Anatomy; LEA & FEBIGER: Philadelphia, PA, USA, 1966. [Google Scholar]
- Hrdlička, A. Practical Anthropometry, 4th ed.; Stewart, T.D., Ed.; Wistar Institute of Anatomy and Biology: Philadelphia, PA, USA, 1952. [Google Scholar]
- Krogman, W.M. The Human Skeleton in Forensic Medicine; Charles C Thomas: Springfield, IL, USA, 1962. [Google Scholar]
- Montagu, M.F.A. Introduction to Physical Anthropology; Charles C Thomas: Springfield, IL, USA, 1960. [Google Scholar]
- Rogers, T.L.; Saunders, S. Accuracy of sex determination using morphological traits of the human pelvis. J. Forensic Sci. 1994, 39, 1047–1056. [Google Scholar] [CrossRef]
- Stewart, T.D. Evaluation of evidence form the skeleton. In Legal Medicine; Gradwohl, R.B.H., Ed.; CV Mosley: St. Louis, MO, USA, 1954. [Google Scholar]
- Rennie, S.R. Summary Sex: A Multivariate Approach to Sex Estimation from the Human Pelvis. Ph.D. Thesis, Liverpool John Moores University, Liverpool, UK, 2018. [Google Scholar]
- Washburn, S.L. Sex difference in the pubic bone of Bantu and Bushman. Am. J. Phys. Anthropol. 1949, 7, 425–432. [Google Scholar] [CrossRef]
- Roth, M.; Ousley, S.D.; Tuamsuk, P. Sex estimation using non-metric traits in Thai crania with the Walker (2008) method. In Proceedings of the 65th Annual Meeting of the American Academy of Forensic Sciences, Washington, DC, USA, 18–23 February 2013. [Google Scholar]
- Winburn, A.P.; Jennings, A.L.; Steadman, D.W.; DiGangi, E.A. Ancestral diversity in skeletal collections: Perspectives on African American body donation. Forensic Anthropol. 2022, 5, 141–152. [Google Scholar] [CrossRef]
- Decker, S.J.; Davy-Jow, S.L.; Ford, J.M.; Hilbelink, D.R. Virtual determination of sex: Metric and nonmetric traits of the adult pelvis from 3D computed tomography models. J. Forensic Sci. 2011, 56, 1107–1114. [Google Scholar] [CrossRef]
- Herrera, M.D.; Tallman, S.D. Craniometric variation and ancestry estimation in two contemporary Caribbean populations. Forensic Sci. Int. 2019, 305, 110013. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.E.; Tise, M.L.; Trammell, L.H.; Anderson, B.E. Cranial morphological variation among contemporary Mexicans: Regional trends, ancestral affinities, and genetic comparisons. Am. J. Phys. Anthropol. 2013, 151, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Richard, A.H.; Parks, C.L.; Monson, K.L. Accuracy of standard craniometrics measurements using multiple data formats. Forensic Sci. Int. 2014, 242, 177–185. [Google Scholar] [CrossRef]
- Ross, A.H. Cranial evidence of pre-contact multiple population expansions in the Caribbean. Caribb. J. Sci. 2004, 40, 291–298. [Google Scholar]
- Stull, K.E.; Tise, M.L.; Ali, Z.; Fowler, D.R. Accuracy and reliability of measurements obtained from computed tomography 3D-volume rendered images. Forensic Sci. Int. 2014, 238, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Field, A. Discovering Statistics Using SPSS, 3rd ed.; SAGE: Thousand Oaks, CA, USA, 2009; pp. 267–678. [Google Scholar]
- Hefner, J.T. Cranial nonmetric variation and estimating ancestry. J. Forensic Sci. 2009, 54, 985–995. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Cohen, J. Weighted Kappa: Nominal scale agreement with provision for scaled disagreement or partial credit. Psychol. Bull. 1968, 70, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Barnes, M.L. Empirical methods and critical race theory: A discourse on possibilities for a hybrid methodology. Wis. L. Rev. 2016, 443, 443–476. [Google Scholar]
- Ford, C.L.; Airhihenbuwa, C.O. Commentary: Just what is critical race theory and what’s it doing in a progressive field like public health? Ethn. Dis. 2018, 28, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, V.D. Critical race empiricism: A new means to measure civil procedure. UC Irvine L. Rev. 2013, 3, 187–216. [Google Scholar]
- Go, M.C.; Yuki, N.; Chu, E.Y. On WEIRD anthropologists and their white skeletons. Forensic Anthropol. 2021, 4, 145–160. [Google Scholar] [CrossRef]
- Williams, B.A.; Rogers, T. Evaluating the accuracy and precision of cranial morphological traits for sex determination. J. Forensic Sci. 2006, 51, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Stinson, S. Sex differences in environmental sensitivity during growth and development. Yearb. Phys. Anthropol. 1985, 28, 123–147. [Google Scholar] [CrossRef]
- Stinson, S. Growth variation: Biological and cultural factors. In Human Biology: An Evolutionary and Biocultural Perspective, 2nd ed.; Stinson, S., Bogin, B., O’Rourke, D., Eds.; Wiley: Hoboken, NJ, USA, 2012; pp. 587–635. [Google Scholar]
- Klales, A.R.; Cole, S.J. Improving nonmetric sex classification for Hispanic individuals. J. Forensic Sci. 2017, 62, 975–980. [Google Scholar] [CrossRef]
- Spradley, M.K.; Jantz, R.L.; Robinson, A.; Peccerelli, F. Demographic change and forensic identification: Problems in metric identification of Hispanic skeletons. J. Forensic Sci. 2008, 53, 21–28. [Google Scholar] [CrossRef]
- Mendéndez, L.P. Comparing methods to assess intraobserver measurement of error of 3D craniofacial landmarks using geometric morphometrics through a digitizer arm. J. Forensic Sci. 2017, 62, 741–746. [Google Scholar] [CrossRef]
- Robinson, C.; Eisma, R.; Morgan, B.; Jeffery, A.; Graham, E.A.M.; Black, S.; Rutty, G.N. Anthropological measurement of lower limb and foot bones using multi-detector computed tomography. J. Forensic Sci. 2008, 53, 1289–1295. [Google Scholar] [CrossRef]
- Pinto, S.C.; Urbanová, P.; Ceaser, R.M., Jr. Two-dimensional wavelet analysis of supraorbital margins of the human skull for characterizing sexual dimorphism. IEEE Trans. Inf. Forensics Secur. 2016, 11, 1542–1548. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (years) | Population Affinity | AMAB | AFAB |
|---|---|---|---|
| 18–30 | African American | 10 | 9 |
| Asian American | 18 | 6 | |
| European American | 10 | 12 | |
| Latin American | 12 | 11 | |
| Native American | 11 | 10 | |
| 31–50 | African American | 11 | 12 |
| Asian American | 19 | 3 | |
| European American | 12 | 11 | |
| Latin American | 11 | 12 | |
| Native American | 11 | 12 | |
| 51–70 | African American | 15 | 18 |
| Asian American | 26 | 8 | |
| European American | 18 | 13 | |
| Latin American | 17 | 16 | |
| Native American | 20 | 17 | |
| 71–90 | African American | 4 | 4 |
| Asian American | 6 | 3 | |
| European American | 3 | 5 | |
| Latin American | 4 | 4 | |
| Native American | 4 | 3 | |
| Total Sample Size | 242 | 189 | |
| Metric Measurements | |
|---|---|
| 1. Minimum frontal breadth (ft-ft) | 10. Bicondylar breadth (cdl-cdl) |
| 2. Orbital height | 11. Biauricular breadth (au-au) |
| 3. Upper facial height (n-pr) | 12. Foramen magnum breadth |
| 4. Parietal chord (b-l) | 13. Occipital chord (l-o) |
| 5. Glabella occipital length (g-op) | 14. Bigonial breadth (go-go) |
| 6. Mastoid length | 15. Basion–bregma height (ba-b) |
| 7. Mandibular length | 16. Basion–nasion length (ba-n) |
| 8. Maximum ramus height | 17. Frontal chord (n-b) |
| 9. Bizygomatic breadth (zy-zy) | 18. Nasal height (n-ns) |
| Stepwise-Selected Classification Functions b | Classification Statistics | |||
|---|---|---|---|---|
| AFAB | AMAB | Overall c | ||
| Population-Inclusive | ||||
| Y = (glabella * 1.385) + (mastoid process * 0.902) + (mental eminence * 0.44) + (−5.888) | N | 149 | 189 | 338 |
| % | 86.6% | 87.1% | 87.0% | |
| Population-Inclusive d | ||||
| Y = (glabella * 1.363) + (mastoid process * 0.876) + (nuchal crest * 0.393) + (−5.664) | N | 103 | 122 | 225 |
| % | 88.0% | 82.4% | 85.0% | |
| African American | ||||
| Y = (glabella * 1.335) + (mastoid process * 1.046) + (−5.164) | N | 33 | 27 | 60 |
| % | 86.8% | 77.1% | 82.0% | |
| Asian American | ||||
| Y = (glabella * 3.033) + (mastoid process * 1.012) + (−6.438) | N | 16 | 61 | 77 |
| % | 84.2% | 93.2% | 91.0% | |
| European American | ||||
| Y = (glabella * 1.628) + (metal eminence * 1.002) + (−6.309) | N | 35 | 34 | 69 |
| % | 87.5% | 82.9% | 85.0% | |
| Latin American | ||||
| Y = (glabella * 1.324) + (nuchal crest * 0.995) + (−5.18) | N | 33 | 33 | 66 |
| % | 89.2% | 80.5% | 85.0% | |
| Native American | ||||
| Y = (glabella * 1.827) + (mastoid process * 1.276) + (−7.037) | N | 35 | 35 | 70 |
| % | 92.1% | 85.4 | 89.0% | |
| Applied Model | Classification Statistics | |||
|---|---|---|---|---|
| AFAB | AMAB | Overall | ||
| Population-Inclusive | N | 156 | 159 | 315 |
| % | 90.7% | 73.3% | 81.0% | |
| Population-Inclusive a | N | 53 | 56 | 109 |
| % | 91.4% | 76.7% | 83.2% | |
| African American | N | 37 | 26 | 63 |
| % | 88.1% | 70.3% | 79.7% | |
| Asian American | N | 16 | 61 | 77 |
| % | 84.2% | 93.8% | 91.7% b | |
| European American | N | 35 | 34 | 69 |
| % | 87.5% | 82.9% | 85.2% | |
| Latin American | N | 38 | 29 | 67 |
| % | 90.5% | 67.4% | 78.8% | |
| Native American | N | 38 | 37 | 75 |
| % | 92.7% | 84.1% | 88.2% | |
| Stepwise-Selected Classification Functions a and Sectioning Points (SP) b | Classification Statistics | |||
|---|---|---|---|---|
| AFAB | AMAB | Overall | ||
| Population-Inclusive | ||||
| Y = (glabella occipital length * 0.057) + (bizygomatic breadth * 0.126) + (biauricular breadth * −0.047) + (minimum frontal breadth * −0.069) + (nasal height * 0.059) + (orbital height * −0.115) + (mastoid height * 0.081) + (bigonial breadth * 0.037) + (maximum ramus height * 0.074) + (mandibular length * −0.046) + (−20.182); SP = −0.221 | N | 146 | 180 | 326 |
| % | 88.0% | 85.7% | 86.7% | |
| African American | ||||
| Y = (bizygomatic breadth * 0.335) + (biauricular breadth * −0.188) + (minimum frontal breadth * −0.185) + (mastoid height * 0.123) + (bicondylar breadth * −0.089) + (maximum ramus height * 0.185) + (−9.561); SP = −0.312 | N | 32 | 30 | 62 |
| % | 84.2% | 88.2% | 86.1% | |
| Asian American | ||||
| Y = (basion–nasion length * 0.142) + (frontal chord * 0.102) + (mastoid height * 0.101) + (−29.68); SP = −0.6335 | N | 15 | 52 | 67 |
| % | 93.8% | 86.7% | 88.2% | |
| European American | ||||
| Y = (bizygomatic breadth * 0.14) + (orbital height * −0.337) + (bigonial breadth * 0.079) + (maximum ramus height * 0.109) + (mandibular length * −0.085) + (−13.013); SP = −0.059 | N | 31 | 33 | 64 |
| % | 81.6% | 80.5% | 81.0% | |
| Latin American | ||||
| Y = (bizygomatic breadth * 0.136) + (maximum ramus height * 0.106) + (−24.507); SP = −0.087 | N | 34 | 30 | 64 |
| % | 82.9% | 71.4% | 77.1% c | |
| Native American | ||||
| Y = (glabella occipital length * 0.082) + (orbital height * −0.197) + (mastoid height * 0.082) + (bigonial breadth * 0.102) + (maximum ramus height * 0.075) + (−25.132); SP = −0.248 | N | 29 | 40 | 69 |
| % | 80.6% | 90.9% | 86.3% | |
| Applied Model | Classification Statistics | |||
|---|---|---|---|---|
| AFAB | AMAB | Overall | ||
| Population-Inclusive | N | 150 | 177 | 327 |
| % | 90.4% | 84.3% | 87.0% | |
| African American | N | 34 | 35 | 69 |
| % | 89.5% | 100% | 95.0% | |
| Asian American | N | 15 | 54 | 69 |
| % | 93.8% | 90.0% | 91.0% | |
| European American | N | 32 | 36 | 68 |
| % | 84.2% | 87.8% | 86.0% | |
| Latin American | N | 35 | 30 | 65 |
| % | 85.4% | 71.4% | 78.0% a | |
| Native American | N | 29 | 42 | 71 |
| % | 80.6% | 95.5% | 89.0% | |
| Stepwise-Selected Classification Functions a,b | Classification Statistics | |||
|---|---|---|---|---|
| AFAB | AMAB | Overall | ||
| Population-Inclusive | ||||
| Y = (glabella * 1.13) + (mastoid * 0.957) + (mental eminence * 0.594) + (glabella occipital length * 0.102) + (bizygomatic breadth * 0.1620) + (maximum ramus height * 0.147) + (mandibular length * −0.101) + (−44.921) | N | 111 | 182 | 293 |
| % | 88.8% | 93.3% | 91.6% | |
| Population-Inclusive c | ||||
| Y = (glabella score * 2.027) + (bizygomatic breadth * 0.263) + (−38.097) | N | 32 | 53 | 85 |
| % | 84.2% | 89.8% | 87.6% | |
| Applied Model | Classification Statistics | |||
|---|---|---|---|---|
| AFAB | AMAB | Overall | ||
| Population-Inclusive | N | 141 | 160 | 301 |
| % | 88.1% | 89.3% | 88.8% | |
| Population-Inclusive a | N | 52 | 57 | 109 |
| % | 91.2% | 86.8% | 88.8% | |
| Morphological Traits a | Kappa Value | Level of Agreement b | Asymptotic SE | Approximate T |
|---|---|---|---|---|
| Nuchal crest | 0.365 | Fair | 0.081 | 5.839 |
| Mastoid | 0.563 | Moderate | 0.088 | 7.267 |
| Supraorbital margin | 0.432 | Moderate | 0.088 | 6.097 |
| Glabella | 0.531 | Moderate | 0.083 | 7.393 |
| Mental eminence | 0.452 | Moderate | 0.088 | 6.210 |
| Measurement | Valid Cases (n) | Valid Cases (%) | Excluded Cases (n) | Total Cases (n) | ICC (for Average Measures) | 95% Confidence Interval |
|---|---|---|---|---|---|---|
| Glabella occipital length | 51 | 98.1% | 1 | 52 | 0.989 | 0.980–0.994 |
| Bizygomatic breadth | 52 | 100% | 0 | 52 | 0.931 | 0.880–0.960 |
| Basion–bregma height | 46 | 88.5% | 6 | 52 | 0.825 | 0.682–0.903 |
| Basion–nasion length | 51 | 98.1% | 1 | 52 | 0.923 | 0.865–0.956 |
| Biauricular breadth | 50 | 96.2% | 2 | 52 | 0.909 | 0.836–0.949 |
| Nasion–prosthion height | 38 | 73.1% | 14 | 52 | 0.981 | 0.961–0.990 |
| Minimum frontal breadth | 52 | 100% | 0 | 52 | 0.880 | 0.792–0.931 |
| Nasal height | 51 | 98.1% | 1 | 52 | 0.941 | 0.896–0.966 |
| Orbital height | 52 | 100% | 0 | 52 | 0.930 | 0.877–0.960 |
| Frontal chord | 47 | 90.4% | 5 | 52 | 0.908 | 0.836–0.949 |
| Parietal chord | 38 | 73.1% | 14 | 52 | 0.777 | 0.575–0.883 |
| Occipital chord | 39 | 75.0% | 13 | 52 | 0.926 | 0.859–0.961 |
| Foramen magnum breadth | 51 | 98.1% | 1 | 52 | 0.980 | 0.965–0.988 |
| Mastoid height | 51 | 98.1% | 1 | 52 | 0.837 | 0.509–0.929 |
| Bigonial breadth | 52 | 100% | 0 | 52 | 0.987 | 0.978–0.003 |
| Bicondylar breadth | 52 | 100% | 0 | 52 | 0.938 | 0.893–0.965 |
| Maximum ramus height | 52 | 100% | 0 | 52 | 0.945 | 0.904–0.968 |
| Mandibular length | 50 | 96.2% | 2 | 52 | 0.904 | 0.813–0.948 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelley, S.R.; Tallman, S.D. Population-Inclusive Assigned-Sex-at-Birth Estimation from Skull Computed Tomography Scans. Forensic Sci. 2022, 2, 321-348. https://doi.org/10.3390/forensicsci2020024
Kelley SR, Tallman SD. Population-Inclusive Assigned-Sex-at-Birth Estimation from Skull Computed Tomography Scans. Forensic Sciences. 2022; 2(2):321-348. https://doi.org/10.3390/forensicsci2020024
Chicago/Turabian StyleKelley, Samantha R., and Sean D. Tallman. 2022. "Population-Inclusive Assigned-Sex-at-Birth Estimation from Skull Computed Tomography Scans" Forensic Sciences 2, no. 2: 321-348. https://doi.org/10.3390/forensicsci2020024