1. Introduction
The linkage between the polyp and medusa stages and their identification as a single nominal species is the most important task of hydrozoan taxonomy. This kind of work has continued for over a hundred years with the aid of many hydrozoan zoologists, but it is still far from complete.
One of the main methods of such work is the rearing of medusae from a hydroid colony or the rearing of the hydroid from the medusa in controlled experimental conditions. This kind of aquarium investigation was achieved for many of the hydroids and medusae found in habitats near well-equipped scientific laboratories and biological stations. However, it is very much more complicated to carry out with hydrozoans inhabiting deep water in areas of the world ocean that are difficult to access (like Arctic areas, etc.).
The taxonomy of hydrozoans of the superfamily Campanulinoidea Bouillon, 1984 (= Family Campanulinidae Hincks, 1868, according to [
1,
2]) is still problematic. Hydroids of this (super-)family are mostly “
Cuspidella-like” (hydrotheca tubular, usually curved, with a conical (pyramidal) operculum consisting of several valves) and are quite similar to each other. Their medusae are rather different, however, being referred to as several genera and even as different families (for details see [
3]).
The medusa
Ptychogena lactea A. Agassiz, 1865 is considered by some authors as a rare species [
1,
2,
4], however, it is well-known to hydrozoan zoologists thanks to its rather large size and noticeable appearance. It is truly rare in boreal waters, but it can be abundant in some subarctic areas and in the Arctic.
The polyps of
P. lactea were unknown for over 100 years. The proposal of a possible hydroid of
P. lactea described by Naumov ([
1]: p. 297, fig. 189) was based on finding of hydroids belonging to the same family Campanulinidae, found in similarly deep water in the neighboring Kara Sea (the medusa was found in the Barents Sea several hundreds of kilometers from these polyps). Thus, this suggestion was not fully established and was later regarded as doubtful or erroneous. The hydroid of
P. lactea was said to be unknown [
4,
5,
6], which was true at the time. Although the first rearing of the hydroid from the medusa
P. lactea, with a description and illustrations, was published in my Russian monograph [
7], more recent authors have still considered the hydroid stage of
P. lactea to be unknown [
8,
9,
10]. More comprehensive evidence is presented here.
2. Material and Methods
The author of this paper participated in a polar expedition of the Zoological Institute of the Russian Academy of Sciences, based at a field station on the Island of Hayes in the middle of the Franz-Josef Land archipelago (80°34′ N; 57°41′ E). The ice on the site was 1.5–2 m thick. The salinity was 33‰ and the temperature of the water was (–) 1.7–1.8 °C, which were the stable the whole year round with exception of the short period of opened water in July–August, when some desalination and “warming up” to 0 °C were observed in upper 1 m water layer thanks to ice molding. Throughout the calendar year from October 1981 to September 1982 regular weekly SCUBA diving and plankton sampling were carried out from two constant ice-holes at depths of 4 m to 30 m for covering a large bottom area. Normally there were two dives through two artificial ice-holes every week with durations of about one hour every dive. The holes for diving were placed above the sublittoral slope. The diver was alone under water, but attached to the signal rope about 60 m long; this allowed the diver to visit the bottom in depth intervals from 2 to 30 m. In total, about 90 dives were executed by three divers.
All material was sorted and identified by specialists with microscopes immediately in the field laboratory in between the dives. Because of the 2 m-thick ice, heavy frost and frequent snow storms the dive together with preparations required the whole working day. In July 1982, when the ice started to break, the dives were stopped for two weeks because of the danger of moving ice and icebergs. In August 1982 the dives were re-established on open water from the boat. In September 1982 the dives were carried out again through the ice-holes in the new ice.
Plankton sampling was executed by plankton net with the same regularity as dives, using the same ice-holes by vertical catch from a depth of 8 m (ice-hole depth limit) to the surface. In the short period of opened water (middle of July–August) the plankton was sampled from the fast ice margin. In such cases the plankton collector was dressed in a diving suite and attached to a rope for security.
The biological situation and seasonal changes of the underwater site were constantly monitored.
There was a cold room (temperature from −1 to 0 °C, which is close to natural water temperature) in a field laboratory with small 30 L aquaria for temporary holding of sampled animals. The aquaria were isolated from each other and from additional inflow or circulation of seawater; just the air was pumped.
When hydromedusae of Ptychogena lactea A. Agassiz, 1865 appeared on the natural site, four alive mature specimens of P. lactea were taken to the field laboratory and placed in an aquarium for rearing hydroids. Aquarial observations were carried out for about three weeks until all medusa died; subsequently, numerous small colonies of hydroids Cuspidella procumbens were discovered everywhere in the aquarium.
3. Results and Discussion
During the winter period of work (October to March) no specimen of P. lactea was found in nature. The first, a single specimen of this medusa, was observed on the site in March. Suddenly in April a very abundant arrival of mature medusae P. lactea was observed in the water column within the inspected depth interval 0–30 m. All these medusae were mature, not young, with a bell diameter of 35 mm to 60 mm. None of these medusae had inhabited this shallow water site before (confirmed by regular observations), but they undoubtedly migrated there from the outer sea and probably came from deeper waters, which are close to the studied site.
The medusae were so numerous that it was easy to catch them from the ice-hole surface with a simple net. However, when performing next weekly dives after a few days of such dense concentration, it was found that all the medusae rapidly disappeared completely and were not recorded again during several months of regular observations at the same site, which were continued until the expedition ended in October.
Medusae in aquaria swam actively for 2 to 3 days; then they dropped down to the bottom, became immobile, died off and soon disintegrated. It is likely the same happened with the P. lactea medusae at the field site as they finished their life-cycle.
Ten to fifteen days after the death of the medusa, several small colonies (of total length 1–1.5 mm, with 1–3 hydranth each) of hydroids were discovered creeping on the walls and on stones and gravel at the bottom of an aquarium. They were typical “Cuspidella-like” hydroids; stolonial colonies with a transparent and very delicate filiform stolon strongly attached to the substrate. It was impossible to detach them safely from the substrate; thus, colonies were collected and fixed together with little stones or pieces of algae.
After the same amount of time passed since medusae had disappeared from the underwater field site, colonies of the same “
Cuspidella-like” hydroids appeared en masse creeping on stones and on the algae
Alaria esculenta and
Delesseria sanguinea. Certainly these are the same hydroids of the medusa
Ptychogena lactea, which were observed in the aquarium. These hydroids were identified as
Cuspidella procumbens Kramp, 1911—a very rarely-found Arctic species for which the life-cycle, the medusa stage and even the gonosome were unknown [
11,
12,
13].
Aquarium and field observations of hydroid colonies of C. procumbens until July did not detect any gonosomes development and aquarial work was stopped. Underwater sampling and inspection of the field sites, however, continued until the end of the expedition in October. The development of gonophores on colonies of C. procumbens (= P. lactea) was eventually recorded in samples from the field site dated 8 September.
In sum, two parallel sets of observations were carried out: one in aquaria and one in the field. It is important to note that during 12 months of frequent field sampling, the only hydroid and hydromedusa from the superfamily Campanulinida (sensu [
14]) found at the natural underwater site at the Island of Hayes in the Franz-Josef Land archipelago were the hydroid
C. procumbens and the hydromedusa
P. lactea. No other hydrozoan “species-candidates” (relative to Campanulinida, Laodiceidae or similar) occured there [
7].
Order Leptothecata Cornelius, 1992 [15] Suborder Conica Broch, 1910 [
16]
Superfamily Campanulinida Bouillon, 1984 [
14]
Family Laodiceidae L. Agassiz, 1862 [
17]
The composition, phylogenetic classification and even the names of Leptothecata higher taxa are under the discussion [
18], which is not a task of the given article.
Ptychogena A. Agassiz, 1865 [
19]
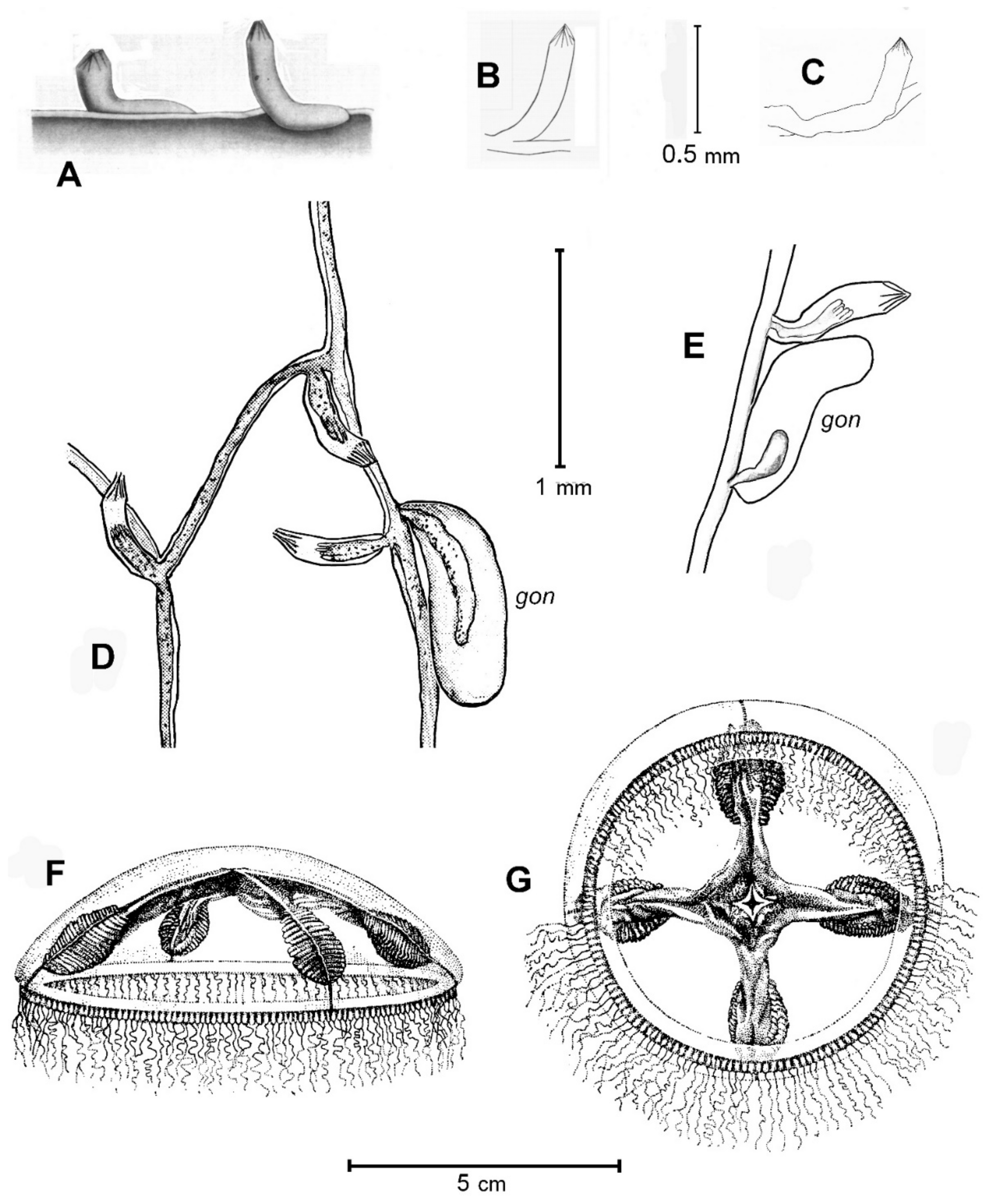
Ptychogena lactea A. Agassiz, 1865 (
Figure 1A–G)
Medusa Ptychogena lactea: A. Agassiz, 1865 [
19]: pp. 137–139, fig. 220–224; Mayer, 1910 [
20]: p. 215; Kramp, 1919 [
21]: pp. 31–39, fig. 5, pl. 3, fig. 1–6; 1933 [
22]: p. 557, fig. 19 and 20; 1959 [
23]: p. 137, fig. 157; 1968 [
24]: pp. 67–68, fig. 175; Yashnov, 1948 [
25]: p. 71, tab. 20, fig. 1; Naumov, 1960 [
1]: pp. 297–299, fig. 189–190 (medusa, non-polyp); Russell, 1953 [
26]: pp. 228–229; 1963 [
27]: p. 3, fig. 7; Arai, Brickmann-Voss, 1980 [
4]: pp. 83–84, fig. 46; Stepanjants, 1989 [
28]: p. 412; Antsulevich, 2015 [
7]: pp. 282–284, fig. 129c,d; Bouillon, Boero, 2000 [
6]: p. 184 (polyp unknown); Minemizu et al., 2015 [
29]: p. 121 (fig.); p. 292; Schuchert, 2017 [
8]: p. 357, fig. 5 (hydroid unknown); 2022 [
10], World Hydrozoa Database, accessed on 16 May 2022 (hydroid unknown); Schuchert et al., 2017 [
9]: p. 174 (hydroid is possibly
Stegopoma-like, but must be considered unknown).
Hydroid Ptychogena lactea: non Naumov, 1960 [
1]: pp. 297–299, fig. 189; non Calder, 1970 [
12]: p. 1512, pl.3, fig. 1; Antsulevich, 2015 [
7]: pp. 282–284, fig. 129a,b (
P. lactea =
C. procumbens).
Hydroid Cuspidella procumbens: Kramp, 1911: p. 384, pl.24, fig. 2–6; Calder, 1970 [
12]: p. 1514, pl.3, fig. 3; Schuchert, 2001 [
13]: pp. 58–59, fig. 44; Antsulevich, 2015 [
7]: pp. 282–284, fig. 129a,b (
C. procumbens =
P. lactea).
3.1. Description (after Material from the Franz-Josef Land Archipelago)
Colonies of hydroids stolonial, creeping by filiform stolons on stones and on algae (Alaria esculenta and Delesseria sanguinea); no upright shoots. Perisarc colorless, transparent, very delicate (thin); for this reason, a colony is sometimes almost invisible on a substrate. Hydrothecae spaced out, the interval between two hydrothecae being notably longer than hydrothecal length. Hydrothecae mostly oriented parallel to the hydrorhiza and with the basal part pressed to it. Occasionally, the hydrotheca can be oriented almost at right angles to the stolon direction.
Hydrotheca is typical “Cuspidella-like”, tubular, non-pedicellate, no diaphragm, the basal part is about two-thirds (rarely half) of the hydrothecal length and may be adnate to the substrate; its distal part curves upward at an obtuse angle (i.e., greater than 90°). Operculum pyramidal, with 10–14 triangular valves. “Renovations” on the hydrotheca were never observed. Polyp with 10 to 12 tentacles.
Gonothecae (firstly described by Antsulevich [
7]) are placed on the stolon near hydrothecae, in pairs with them. Gonotheca single, of unusual form, not upright, but flat and laying in a reclining position, adnate to the substratum along its length. Gonotheca elongated, nearly twice the length of a hydrotheca, slightly curved. Its perisarc, molded when soft, is flat when in contact with the substrate or the adjacent stolon, otherwise broadly convex. A blastostyle without medusa buds was observed inside a gonotheca on 8th September (water temperature was −1.7 °C).
Dimensions: Hydrotheca total length 0.62–0.73 mm; height of the free curved part (with operculum) 0.31–0.41 mm; width 0.15–0.16 mm; stolon thickness 0.07 mm. Gonotheca length 0.96–1.0 mm; width 0.24–0.27 mm.
Mature medusa large, bell diameter 35–60 mm. The convex umbrella may be hemispherical or flat. The mesoglea is very thick. Stomach short, less than one half of umbrella height, quadrangular, with four short oral lips which may be slightly folded or crenulated. Manubrium with funnel-shaped lobes connected to four radial canals. Each radial canal with 20 to 30 lamelliform diverticulae on either side, in which are located the gonads. Each diverticulum bears one lamina, and gonads cover the lamellar folds. They form flat oval structures below the radial canals. The umbrella margin with numerous tentacles, up to a maximum of 500 (usually fewer). One to three small cordyli occur between neighboring tentacles. No cirri or ocelli.
In all specimens from Franz-Josef Land the bell is colorless, and gonads, radial canals and tentacles are milk-white (this color was a characteristic for the specific name “lactea”); sometimes some are yellowish.
To illustrate the medusa (
Figure 1F,G) is given, probably still the best drawing from Naumov [
1]. For the morphology and coloration of live specimens, see also good photographs taken in waters of NE Canada and Japan [
29,
30].
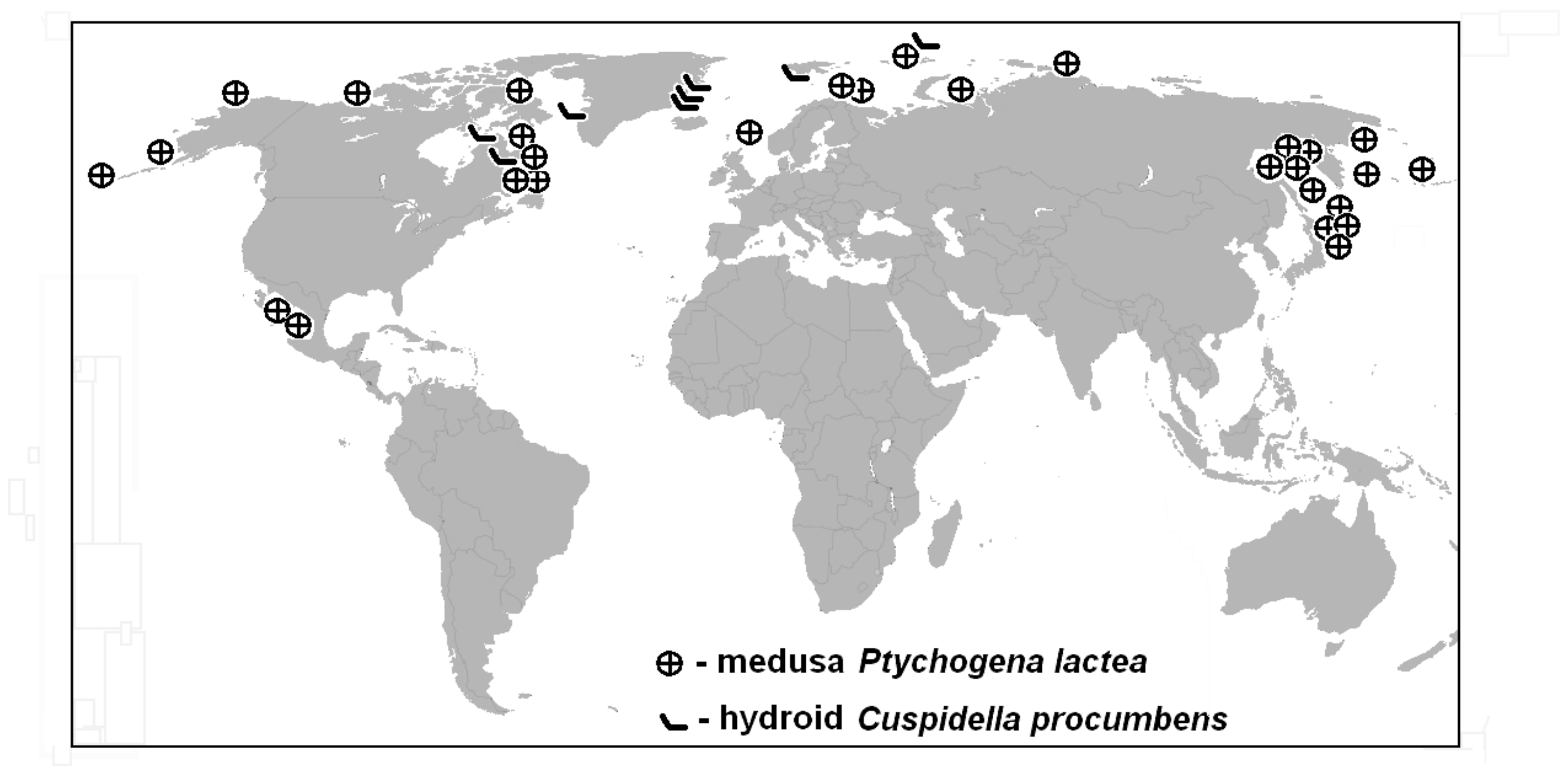
3.2. Distribution of the Hydroid (C. procumbens)
It is evident that large and noticeable medusae were found much more frequently than small transparent hydroids. That is why the data on medusa distribution predominates and is noted separately.
Type locality: north-east Greenland, area close to Danmarks Havn and off Cape Bismark [
11]. There were only few reports of this hydroid [
7,
11,
12]. Schuchert [
13] examined the same museum material from Greenland which was described by Kramp [
11].
Because of these limited findings, the ecology of
C. procumbens is poorly known. Vertical distribution is as follows: E Greenland (type locality)—Danmarks Havn (6–10 m), Stormbugt Bay (20–40 m; 20–30 m; 15–20 m), off Cape Bismark (40–60 m; 30–40 m) and one locality from 300 m [
11]. NE Canada—N Foxe Basin (106 m), SE Hudson Bay (100–107 m) [
12]. Franz-Josef Land, Island of Hayes (4–18 m) [
7]. Spitsbergen Archipelago (depth interval was not indicated, but judging from the substrate “algae” it should be a shallow water) [
31]. The substrate where hydroids were recorded is stones, gravel, algae, bryozoans and erect colonies of other hydroids. All localities belong to Arctic and even to high Arctic waters. No detailed data on ecology was provided in the literature.
The site in the Franz-Josef Land archipelago is characterized by thick ice coverage almost all year round, constant salinity 33‰ and negative temperature (−1.7–1.8 °C). Nevertheless, all 46 species of hydroids which have been found there demonstrate normal seasonality of development and life-circles, as those in temperate waters [
7].
3.3. Distribution of the Medusa P. lactea
Type locality: Atlantic Ocean, Massachusetts Bay, Nahant [
19]. Recently-known distribution of the medusa is much wider compared to the hydroid (
Figure 2). Circumpolar in the Arctic including high latitudes: coasts of Greenland, Franz-Josef Land, Barents Sea, Kara Sea, Laptev Sea, Beaufort Sea ([
3,
11,
20,
32] as
P. pinnatulata; [
1,
4,
7,
13,
25]).
Several records and some good photographs by Claude Nozeres from NE Canada: Baffin Island (Frobisher Bay), Labrador Sea (Smith Sound, Jones Sound), Nova Scotia, Halifax, Quebec—Rimouski, Fleuve Saint-Laurent [
5,
20,
30].
Found between Iceland and Ireland, Faroe-Shetland Channel [
8,
20]. In the Atlantic, in the opinion of Kramp [
23], the medusa may “occasionally” penetrate southwards as far as the Cape Cod region.
Most of the records and reports of abundance, however, were from the North Pacific: the Bering Sea, near Kamtchatka, the Commandor and Aleutean depression, the whole Okhotsk Sea, where the most abundant and sustainable population was recorded in the northern part of this sea; the oceanic side of the Kuril Islands and Kuril-Kamtchatka trench. It was not found in the warmer Sea of Japan [
1,
2,
33]; off Hokkaido, [
29,
34], British Columbia and Puget Sound, Strait of Georgia [
4].
For references see the distribution notes within the text.
P. lactea has been recorded from a depth of 1500 m [
1,
34] up to the surface [
7], but mostly at depths over 250 m. Vertical hauls with a plankton net would often be taken from deep water to the surface, so that the exact depth where a medusa was caught remained unknown. Specimens were recorded as “deep-water to 0 m” together with the maximum depth of sampling. This led to several discussions about whether it was a deep-water medusa or not. The comment “
P. lactea may be found near the surface where the water is very cold” [
4,
21] was fully confirmed in Franz-Josef Land, where the coldest water of the world ocean occurs [
7].
Seasonal changes and regular diel vertical migrations of
P. lactea were studied by Zavolokin [
33,
35] in the Russian Far Eastern seas. He had a large amount of this medusa material because the investigations were performed not with a plankton net (as usual), but with a special large trawl.
It has been demonstrated that
P. lactea is a dominant species among the large-sized hydromedusae by its abundance in the northern part of the Okhotsk Sea.
P. lactea is mainly a mesopelagic species. It has been found at every depth within 100–800 m, but mostly concentrated within the horizon 200–300 m. Diel migrations peculiar to this medusa occur in the open Okhotsk Sea, where it moves upwards every night to the 100–50 m depth horizon. Statistics show that individual medusae which undertake this migration into shallow water have a larger umbrella and are more mature, whereas the smaller ones prefer to stay deeper [
33,
35]. This may be important for sexual reproduction, because in the shallow zone hydroids may find better conditions and more available substrates to settle on. Hydroids were not found in deep water.
During the year, the greatest abundance of
P. lactea was observed in winter and in spring; in summer these medusae disappear from the water column [
33,
35], which corresponds in general to the author’s observations in Franz-Josef Land.
In the central and the northern parts of the Okhotsk Sea, the water temperature and salinity are quite similar to the high Arctic, both during the long winter period at all depths and in the mesopelagial zone during summer; in the Okhotsk Sea, water of temperature −1.7° to 0° is available for cold water animals like
P. lactea all year round [
36].
A wide variety of environmental factors linked to the occurrence of
P. lactea was reported for Northern Canada. In Frobisher Bay five specimens of the medusa were found at two stations at depths of 225 m and 92 m, and from 20 m or less in the Beaufort Sea [
5]. The salinity in the estuary of the Fleuve Saint-Laurent was evidently rather lower than a normal oceanic one.
It was reported that the main food of
P. lactea was eggs of various fish, mostly eggs of
Gadus chalcogrammus spawning in cold water at spring [
33].
From several known world-wide medusae species of the genus
Ptychogena, the only species for which the polyp stage is known is
Ptychogena crocea, Kramp & Dumas, 1925. The DNA sequence comparisons allow the conclusion that
P. crocea is the medusa stage of the hydroid
Stegopoma plicatile (M. Sars, 1863) [
9]. All “
Cuspidella-like” hydroids have a similar structure, whereas their medusae are more diverse and have been split into several genera and the above families [
3]. It was reasonably suggested that the other species of the genus
Ptychogena could have a similar hydroid [
8], what means that the unknown hydroid of
P. lactea could have “
Stegopoma-type” structure as well. However, according to the observations presented here, the hydroid reared from the medusa
P. lactea has the “
Cuspidella-like” appearance. This lack of coincidence may be finally resolved later by experimental rearing of identifiable (close to maturation) medusae from colonies of
Cuspidella procumbens and by DNA sequence comparison.
4. Conclusions
The links between the medusa P. lactea with its unknown hydroid and the hydroid C. procumbens with its unknown medusa were proposed by aquarium experiments and field observations in the Franz-Josef Land Archipelago. The two nominal species are proposed to be united here under the name Ptychogena lactea A. Agassiz, 1865, which has a priority. This suggestion should be confirmed later by additional experimental rearing of the well-identifiable medusae from the C. procumbens hydroids and by DNA sequence comparison.
The gonotheca of the hydroid is of unusual shape, flat and completely adnate to the substrate.
The geographical distribution both of the hydroid and the medusa coincide completely despite very few records of the hydroid.
P. lactea is typically an Arctic species; some records from the high-boreal zone do not constitute exceptions because this species selects “close-to-Arctic” environmental conditions everywhere by vertical migrations and distribution in the temperature gradient.
P. lactea is not a rare species and in its typical habitats it may be quite abundant. These habitats can be difficult to access for investigators, however, which makes it a “rare” species in practice.
Newly-liberated medusae were not observed; evidently they are released in the late autumn. Mature medusae are abundant in late winter–spring, but mostly disappear by summer. New hydroids appear in May under ice coverage; in the high Arctic, gonosomes develop in September.
P. lactea is also not really a deep-water species. Vertical distribution of medusa and hydroid stages are rather different. The medusa is mostly mesopelagic; the hydroid inhabits shallow water. The medusae of P. lactea probably spawn only in the shallow water zone, perhaps once a year, which gives their hydroids more chance to survive, and later to develop and liberate the next generation of young medusae. As for many other jellyfish, vertical and spatial migrations are typical for P. lactea medusae.


{kind=link}
{kind=link}

