

Co-Producing Phycocyanin and Bioplastic in Arthrospira platensis Using Carbon-Rich Wastewater
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Species, Inoculum Preparation and Culture Condition
2.2. Culture Productivity and Health
2.3. Chemical Analyses
2.3.1. Phycocyanin
2.3.2. Polyhydroxybutyrate (PHB)
2.4. Statistical Analysis
3. Results and Discussion
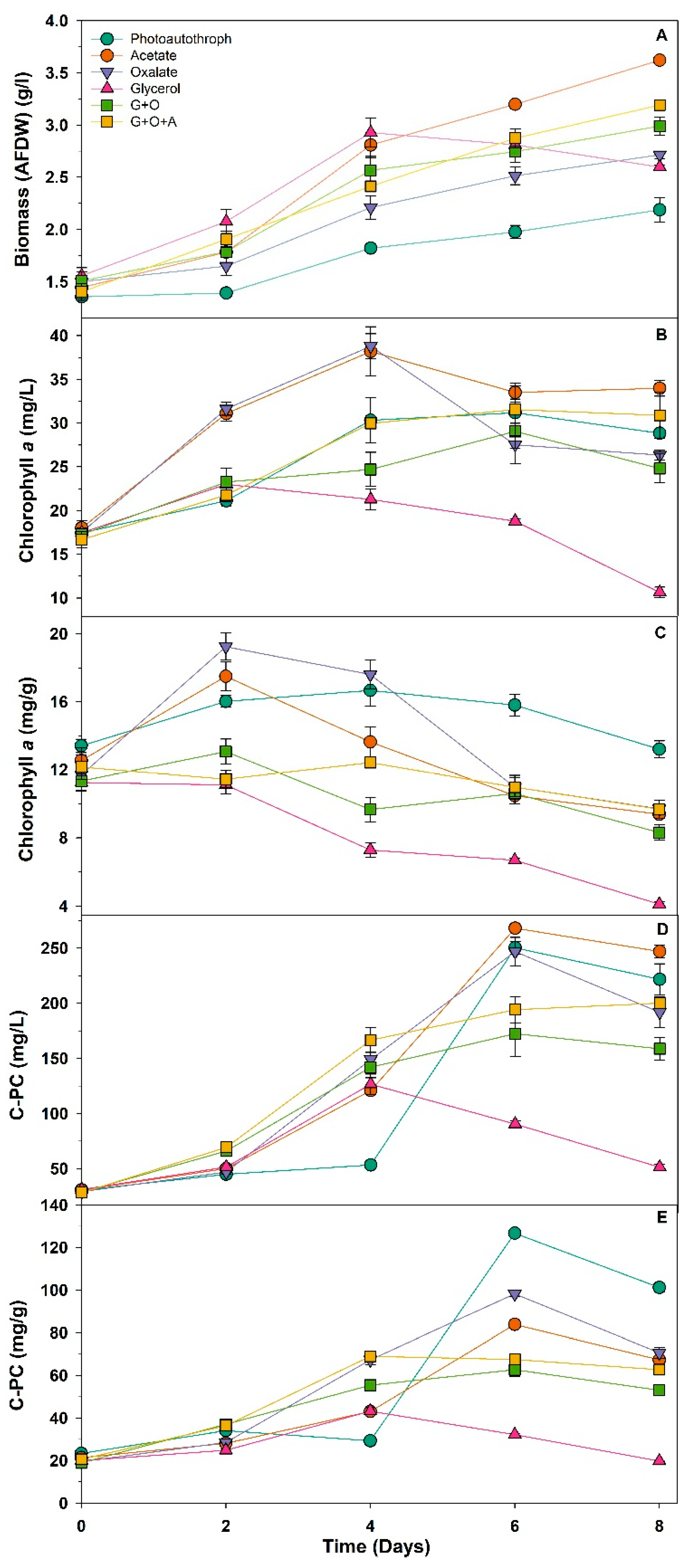
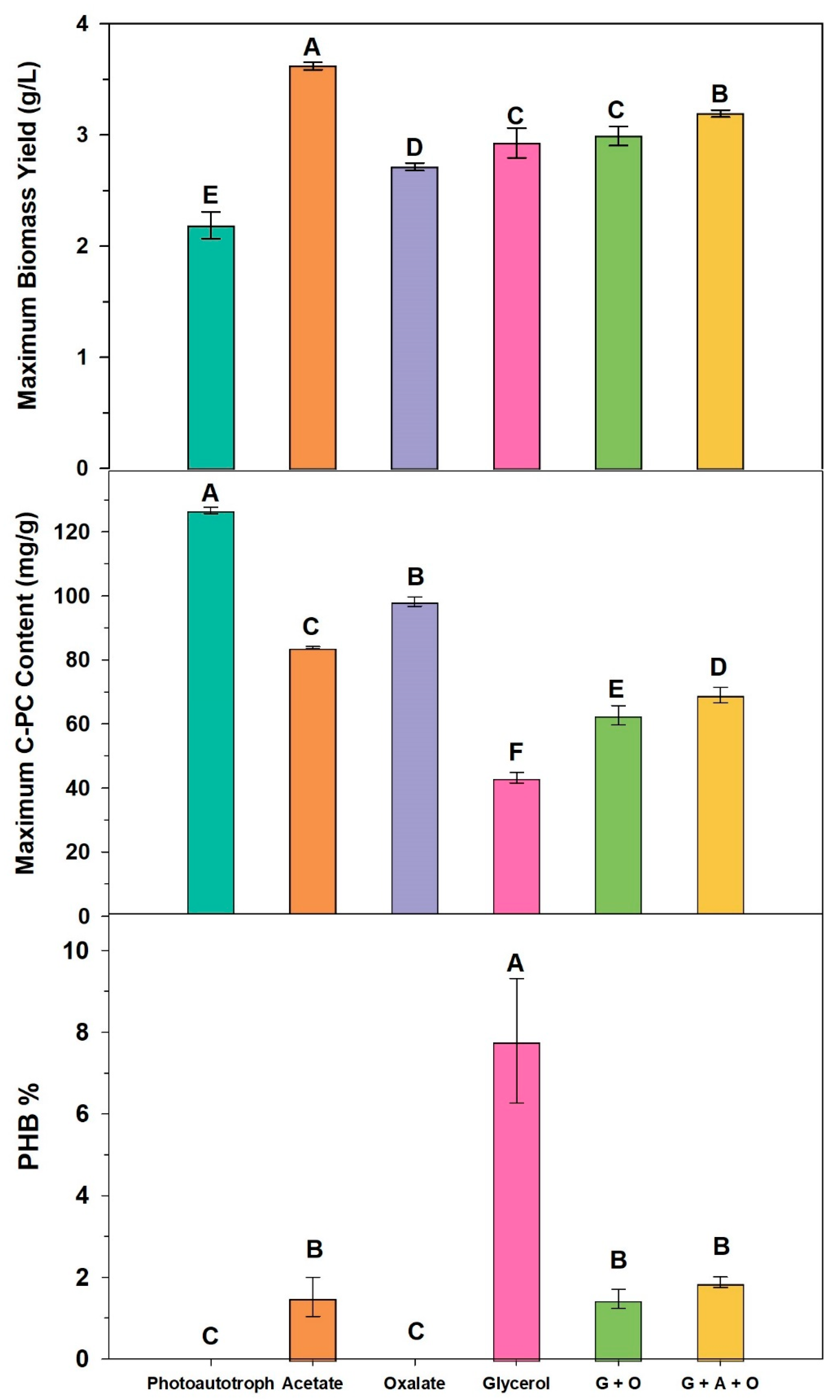
3.1. Biomass and Pigment Production
3.2. Effects of Organic Carbon Source on Phycocyanin and PHA Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PHA | polyhydroxyalkanoate |
| PHB | polyhydroxybutyrate |
| C-PC | phycocyanin |
| PUFA | polyunsaturated fatty acid |
References
- Russo, G.L.; Langellotti, A.L.; Oliviero, M.; Sacchi, R.; Masi, P. Sustainable production of food grade omega-3 oil using aquatic protists: Reliability and future horizons. New Biotechnol. 2021, 62, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A Precious Bio-resource in Agriculture, Ecosystem, and Environmental Sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.K.; Kaur, R.; Bansal, A.; Kapur, S.; Sundaram, S. Biotechnological exploitation of cyanobacteria and microalgae for bioactive compounds. In Biotechnological Production of Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2020; pp. 221–259. [Google Scholar]
- Borowitzka, M.A. Algal biotechnology. In The Algae World; Springer: Dordrecht, The Netherland, 2015; pp. 319–338. [Google Scholar]
- Vernès, L.; Granvillain, P.; Chemat, F.; Vian, M. Phycocyanin from Arthrospira platensis. Production, extraction and analysis. Curr. Biotechnol. 2015, 4, 481–491. [Google Scholar] [CrossRef]
- Nwoba, E.G.; Parlevliet, D.A.; Laird, D.W.; Alameh, K.; Moheimani, N.R. Sustainable phycocyanin production from Arthrospira platensis using solar-control thin film coated photobioreactor. Biochem. Eng. J. 2019, 141, 232–238. [Google Scholar] [CrossRef]
- Nwoba, E.G.; Parlevliet, D.A.; Laird, D.W.; Alameh, K.; Moheimani, N.R. Outdoor phycocyanin production in a standalone thermally-insulated photobioreactor. Bioresour. Technol. 2020, 315, 123865. [Google Scholar] [CrossRef]
- Subhashini, J.; Mahipal, S.V.; Reddy, M.C.; Reddy, M.M.; Rachamallu, A.; Reddanna, P. Molecular mechanisms in C-Phycocyanin induced apoptosis in human chronic myeloid leukemia cell line-K562. Biochem. Pharmacol. 2004, 68, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Ravi, M.; De, S.L.; Azharuddin, S.; Paul, S.F. The beneficial effects of Spirulina focusing on its immunomodulatory and antioxidant properties. Nutr. Diet. Suppl. 2010, 2, 73–83. [Google Scholar]
- Rodríguez-Sánchez, R.; Ortiz-Butrón, R.; Blas-Valdivia, V.; Hernández-García, A.; Cano-Europa, E. Phycobiliproteins or C-phycocyanin of Arthrospira (Spirulina) maxima protect against HgCl2-caused oxidative stress and renal damage. Food Chem. 2012, 135, 2359–2365. [Google Scholar] [CrossRef]
- Gellenbeck, K.W. Utilization of algal materials for nutraceutical and cosmeceutical applications—What do manufacturers need to know? J. Appl. Phycol. 2012, 24, 309–313. [Google Scholar] [CrossRef]
- Sun, L.; Wang, S.; Chen, L.; Gong, X. Promising fluorescent probes from phycobiliproteins. IEEE J. Sel. Top. Quantum Electron. 2003, 9, 177–188. [Google Scholar] [CrossRef]
- Singh, A.K.; Sharma, L.; Mallick, N.; Mala, J. Progress and challenges in producing polyhydroxyalkanoate biopolymers from cyanobacteria. J. Appl. Phycol. 2017, 29, 1213–1232. [Google Scholar] [CrossRef]
- McAdam, B.; Brennan Fournet, M.; McDonald, P.; Mojicevic, M. Production of polyhydroxybutyrate (PHB) and factors impacting its chemical and mechanical characteristics. Polymers 2020, 12, 2908. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Kiran, B.R.; Mohan, S.V. Polyhydroxybutyrate production by Chlorella sorokiniana SVMIICT8 under Nutrient-deprived mixotrophy. Bioresour. Technol. 2022, 354, 127135. [Google Scholar] [CrossRef]
- Costa, S.S.; Miranda, A.L.; de Morais, M.G.; Costa, J.A.V.; Druzian, J.I. Microalgae as source of polyhydroxyalkanoates (PHAs)—A review. Int. J. Biol. Macromol. 2019, 131, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.J.; Oliveira Dalla Valentina, L.V.; Hidalgo Schulz, A.A.; Tomaz Duarte, M.A. From obtaining to degradation of PHB: Material properties. Part I. Ing. Y Cienc. 2017, 13, 269–298. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y. Plastic bacteria? Progress and prospects for polyhydroxyalkanoate production in bacteria. Trends Biotechnol. 1996, 14, 431–438. [Google Scholar] [CrossRef]
- Troschl, C.; Meixner, K.; Drosg, B. Cyanobacterial PHA production—Review of recent advances and a summary of three years’ working experience running a pilot plant. Bioengineering 2017, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, M.; Nakai, K.; Miyake, M.; Asada, Y.; Taya, M. Production of poly-β-hydroxybutyrate by thermophilic cyanobacterium, Synechococcus sp. MA19, under phosphate-limited conditions. Biotechnol. Lett. 2001, 23, 1095–1099. [Google Scholar] [CrossRef]
- Sharma, L.; Mallick, N. Enhancement of poly-β-hydroxybutyrate accumulation in Nostoc muscorum under mixotrophy, chemoheterotrophy and limitations of gas-exchange. Biotechnol. Lett. 2005, 27, 59–62. [Google Scholar] [CrossRef]
- Jau, M.-H.; Yew, S.-P.; Toh, P.S.; Chong, A.S.; Chu, W.-L.; Phang, S.-M.; Najimudin, N.; Sudesh, K. Biosynthesis and mobilization of poly (3-hydroxybutyrate)[P (3HB)] by Spirulina platensis. Int. J. Biol. Macromol. 2005, 36, 144–151. [Google Scholar] [CrossRef]
- Eriksen, N.T. Production of phycocyanin—A pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-S.; Wu, J.-Y. Effect of carbon sources on growth and lipid accumulation of newly isolated microalgae cultured under mixotrophic condition. Bioresour. Technol. 2015, 184, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, J.F.; Cerón García, M.; Sánchez Mirón, A.; Belarbi, E.H.; Camacho, F.G.; Grima, E.M. Pilot-plant-scale outdoor mixotrophic cultures of Phaeodactylum tricornutum using glycerol in vertical bubble column and airlift photobioreactors: Studies in fed-batch mode. Biotechnol. Prog. 2004, 20, 728–736. [Google Scholar]
- Fan, X.; Burton, R.; Zhou, Y. Glycerol (byproduct of biodiesel production) as a source for fuels and chemicals mini review. Open Fuels Energy Sci. J. 2010, 3, 17–22. [Google Scholar] [CrossRef] [Green Version]
- White, C.; Laird, D.W.; Hughes, L.J. From carbon waste to carbon product: Converting oxalate to polyhydroxybutyrate using a mixed microbial culture. J. Environ. Chem. Eng. 2017, 5, 2362–2365. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, Y. High cell density mixotrophic culture of Spirulina platensis on glucose for phycocyanin production using a fed-batch system. Enzym. Microb. Technol. 1997, 20, 221–224. [Google Scholar] [CrossRef]
- Chojnacka, K.; Noworyta, A. Evaluation of Spirulina sp. growth in photoautotrophic, heterotrophic and mixotrophic cultures. Enzym. Microb. Technol. 2004, 34, 461–465. [Google Scholar] [CrossRef]
- Chojnacka, K.; Zielińska, A. Evaluation of growth yield of Spirulina (Arthrospira) sp. in photoautotrophic, heterotrophic and mixotrophic cultures. World J. Microbiol. Biotechnol. 2012, 28, 437–445. [Google Scholar] [CrossRef]
- Marquez, F.J.; Sasaki, K.; Kakizono, T.; Nishio, N.; Nagai, S. Growth characteristics of Spirulina platensis in mixotrophic and heterotrophic conditions. J. Ferment. Bioeng. 1993, 76, 408–410. [Google Scholar] [CrossRef]
- Vonshak, A.; Cheung, S.M.; Chen, F. Mixotrophic growth modifies the response of Spirulina (Arthrospira) platensis (Cyanobacteria) cells to light. J. Phycol. 2000, 36, 675–679. [Google Scholar] [CrossRef] [Green Version]
- Narayan, M.; Manoj, G.; Vatchravelu, K.; Bhagyalakshmi, N.; Mahadevaswamy, M. Utilization of glycerol as carbon source on the growth, pigment and lipid production in Spirulina platensis. Int. J. Food Sci. Nutr. 2005, 56, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zheng, W.; Yang, F.; Bai, Y.; Wong, Y.-S. Mixotrophic culture of high selenium-enriched Spirulina platensis on acetate and the enhanced production of photosynthetic pigments. Enzym. Microb. Technol. 2006, 39, 103–107. [Google Scholar] [CrossRef]
- Nematollahi, M.A.; Laird, D.W.; Hughes, L.J.; Raeissosadati, M.; Moheimani, N.R. Effect of organic carbon source and nutrient depletion on the simultaneous production of a high value bioplastic and a specialty pigment by Arthrospira platensis. Algal Res. 2020, 47, 101844. [Google Scholar] [CrossRef]
- Yadav, V.K.; Yadav, V.K. Impact of differnet carbon supplement on extraction of C-phycocyanin (C-PC) followed by poly-beta-hydroxybutyrate (PHB) from Nostoc Muscorum. Int. J. Pharm. Sci. Res. 2017, 8, 2469–2476. [Google Scholar] [CrossRef]
- Zarrouk, C. Contribution a l’etude d’une Cyanophycee. Influence de Divers Facteurs Physiques et Chimiques sur la Croissance et la Photosynthese de Spirulina mixima. Ph.D. Thesis, University of Paris, Paris, France, 1966. [Google Scholar]
- Ogbonna, J.C.; Soejima, T.; Ugwu, C.U.; Tanaka, H. An integrated system of solar light, artificial light and organic carbon supply for cyclic photoautotrophic-heterotrophic cultivation of photosynthetic cells under day–night cycles. Biotechnol. Lett. 2001, 23, 1401–1406. [Google Scholar] [CrossRef]
- Moheimani, N.R.; Borowitzka, M.A.; Isdepsky, A.; Sing, S.F. Standard methods for measuring growth of algae and their composition. In Algae for Biofuels and Energy; Springer: Dordrecht, The Netherlands, 2013; pp. 265–284. [Google Scholar]
- Jeffrey, S.t.; Humphrey, G. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]
- De Morais, E.G.; Druzian, J.I.; Nunes, I.L.; de Morais, M.G.; Costa, J.A.V. Glycerol increases growth, protein production and alters the fatty acids profile of Spirulina (Arthrospira) sp LEB 18. Process Biochem. 2019, 76, 40–45. [Google Scholar] [CrossRef]
- Martins, A.A.; Mata, T.M.; Oliveira, O.; Oliveira, S.; Mendes, A.M.; Caetano, N.S. Sustainability evaluation of biodiesel from Arthrospira platensis and Chlorella vulgaris under mixotrophic conditions and salinity stress. Chem. Eng. Trans. 2016, 49, 571–576. [Google Scholar]
- Matsudo, M.C.; Moraes, F.A.; Bezerra, R.P.; Arashiro, R.E.; Sato, S.; Carvalho, J.C.M. Use of acetate in fed-batch mixotrophic cultivation of Arthrospira platensis. Ann. Microbiol. 2015, 65, 1721–1728. [Google Scholar] [CrossRef]
- Pereira, M.I.; Chagas, B.M.; Sassi, R.; Medeiros, G.F.; Aguiar, E.M.; Borba, L.H.; Silva, E.P.; Neto, J.C.A.; Rangel, A.H. Mixotrophic cultivation of Spirulina platensis in dairy wastewater: Effects on the production of biomass, biochemical composition and antioxidant capacity. PLoS ONE 2019, 14, e0224294. [Google Scholar] [CrossRef] [Green Version]
- Setyoningrum, T.M.; Nur, M.A. Optimization of C-phycocyanin production from S. platensis cultivated on mixotrophic condition by using response surface methodology. Biocatal. Agric. Biotechnol. 2015, 4, 603–607. [Google Scholar] [CrossRef]
- Zhai, J.; Li, X.; Li, W.; Rahaman, M.H.; Zhao, Y.; Wei, B.; Wei, H. Optimization of biomass production and nutrients removal by Spirulina platensis from municipal wastewater. Ecol. Eng. 2017, 108, 83–92. [Google Scholar] [CrossRef]
- Zhang, X.-W.; Zhang, Y.-M.; Chen, F. Application of mathematical models to the determination optimal glucose concentration and light intensity for mixotrophic culture of Spirulina platensis. Process Biochem. 1999, 34, 477–481. [Google Scholar] [CrossRef]
- Yang, C.; Hua, Q.; Shimizu, K. Energetics and carbon metabolism during growth of microalgal cells under photoautotrophic, mixotrophic and cyclic light-autotrophic/dark-heterotrophic conditions. Biochem. Eng. J. 2000, 6, 87–102. [Google Scholar] [CrossRef]
- Cheirsilp, B.; Torpee, S. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: Effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour. Technol. 2012, 110, 510–516. [Google Scholar] [CrossRef]
- Nogueira, D.A.; da Silveira, J.M.; Vidal, E.M.; Ribeiro, N.T.; Veiga Burkert, C.A. Cell disruption of Chaetoceros calcitrans by microwave and ultrasound in lipid extraction. Int. J. Chem. Eng. 2018, 2018, 9508723. [Google Scholar] [CrossRef] [Green Version]
- Klotz, A.; Georg, J.; Bučinská, L.; Watanabe, S.; Reimann, V.; Januszewski, W.; Sobotka, R.; Jendrossek, D.; Hess, W.R.; Forchhammer, K. Awakening of a dormant cyanobacterium from nitrogen chlorosis reveals a genetically determined program. Curr. Biol. 2016, 26, 2862–2872. [Google Scholar] [CrossRef] [Green Version]
- Duangsri, C.; Mudtham, N.-A.; Incharoensakdi, A.; Raksajit, W. Enhanced polyhydroxybutyrate (PHB) accumulation in heterotrophically grown Arthrospira platensis under nitrogen deprivation. J. Appl. Phycol. 2020, 32, 3645–3654. [Google Scholar] [CrossRef]
- Corrêa, P.S.; Teixeira, C.M.L.L. Polyhydroxyalkanoates and pigments coproduction by Arthrospira (Spirulina) platensis cultivated in crude glycerol. J. Appl. Phycol. 2021, 33, 1487–1500. [Google Scholar] [CrossRef]
- Choi, G.-G.; Bae, M.-S.; Ahn, C.-Y.; Oh, H.-M. Induction of axenic culture of Arthrospira (Spirulina) platensis based on antibiotic sensitivity of contaminating bacteria. Biotechnol. Lett. 2008, 30, 87–92. [Google Scholar] [CrossRef]
- Mogale, M. Identification and Quantification of Bacteria Associated with Cultivated Spirulina and Impact of Physiological Factors; University of Cape Town: Cape Town, South Africa, 2016. [Google Scholar]
- Markou, G.; Kougia, E.; Kefalogianni, I.; Tsagou, V.; Arapoglou, D.; Chatzipavlidis, I. Effect of glycerol concentration and light intensity on growth and biochemical composition of Arthrospira (Spirulina) platensis: A study in semi-continuous mode with non-aseptic conditions. Appl. Sci. 2019, 9, 4703. [Google Scholar] [CrossRef] [Green Version]
- Grama, B.S.; Agathos, S.N.; Jeffryes, C.S. Balancing photosynthesis and respiration increases microalgal biomass productivity during photoheterotrophy on glycerol. ACS Sustain. Chem. Eng. 2016, 4, 1611–1618. [Google Scholar] [CrossRef]
- Da Silva, C.K.; Costa, J.A.V.; de Morais, M.G. Polyhydroxybutyrate (PHB) synthesis by Spirulina sp. LEB 18 using biopolymer extraction waste. Appl. Biochem. Biotechnol. 2018, 185, 822–833. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.M.; Huchzermeyer, B.; Rath, T. PHB accumulation in Nostoc muscorum under different carbon stress situations. J. Appl. Phycol. 2012, 24, 157–162. [Google Scholar] [CrossRef]
- Singh, M.K.; Rai, P.K.; Rai, A.; Singh, S.; Singh, J.S. Poly-β-hydroxybutyrate production by the cyanobacterium Scytonema geitleri Bharadwaja under varying environmental conditions. Biomolecules 2019, 9, 198. [Google Scholar] [CrossRef] [Green Version]
- Khetkorn, W.; Incharoensakdi, A.; Lindblad, P.; Jantaro, S. Enhancement of poly-3-hydroxybutyrate production in Synechocystis sp. PCC 6803 by overexpression of its native biosynthetic genes. Bioresour. Technol. 2016, 214, 761–768. [Google Scholar] [CrossRef]
- Krasaesueb, N.; Incharoensakdi, A.; Khetkorn, W. Utilization of shrimp wastewater for poly-β-hydroxybutyrate production by Synechocystis sp. PCC 6803 strain ΔSphU cultivated in photobioreactor. Biotechnol. Rep. 2019, 23, e00345. [Google Scholar] [CrossRef]
- Hauf, W.; Schlebusch, M.; Hüge, J.; Kopka, J.; Hagemann, M.; Forchhammer, K. Metabolic changes in Synechocystis PCC6803 upon nitrogen-starvation: Excess NADPH sustains polyhydroxybutyrate accumulation. Metabolites 2013, 3, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Andrade, M.R.; Costa, J.A. Mixotrophic cultivation of microalga Spirulina platensis using molasses as organic substrate. Aquaculture 2007, 264, 130–134. [Google Scholar] [CrossRef]
- Ye, H.; He, Y.; Xie, Y.; Sen, B.; Wang, G. Fed-batch fermentation of mixed carbon source significantly enhances the production of docosahexaenoic acid in Thraustochytriidae sp. PKU# Mn16 by differentially regulating fatty acids biosynthetic pathways. Bioresour. Technol. 2020, 297, 122402. [Google Scholar] [PubMed]
- Zou, S.P.; Wang, Z.J.; Zhao, K.; Zhang, B.; Niu, K.; Liu, Z.Q.; Zheng, Y.G. High-level production of d-pantothenic acid from glucose by fed-batch cultivation of Escherichia coli. Biotechnol. Appl. Biochem. 2021, 68, 1227–1235. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Specific Growth Rate (d−1) | Maximum Specific Growth Rate (d−1) * | Biomass Productivity (mg/l/d) | Maximum Biomass Productivity (mg/l/d) * | Maximum C-PC Productivity (mg/l/d) ** | PHB Productivity (mg/l/d) | |
|---|---|---|---|---|---|---|
| Photoautotroph | 0.09 ± 0.004 c | 0.12 ± 0.001 d | 111 ± 7 f | 130 ± 31 d | 36.7 ± 0.5 ab | 0 c |
| Acetate | 0.17 ± 0.008 a | 0.24 ± 0.009 a | 272 ± 8 a | 341 ± 9 a | 39.6 ± 0.1 a | 3.6 ± 1.1 b |
| Oxalate | 0.11 ± 0.002 bc | 0.14 ± 0.004 d | 152 ± 2 d | 177 ± 9 c | 36.3 ± 1.2 ab | 0 c |
| Glycerol | 0.09 ± 0.009 c | 0.23 ± 0.009 ab | 130 ± 7 e | 344 ± 16 a | 23.9 ± 1.2 d | 11.1 ± 1.7 a |
| G + O | 0.12 ± 0.003 b | 0.19 ± 0.002 c | 183 ± 1 c | 260 ± 11 b | 28.3 ± 0.1 c | 3.6 ± 0.7 b |
| G + O + A | 0.15 ± 0.006 a | 0.20 ± 0.009 bc | 227 ± 5 b | 259 ± 7 b | 34.6 ± 1.5 b | 3.9 ± 0.7 b |
| Algal Species | Culture Medium | Culture Mode | Phycocyanin (% w/w) | Max PHB (% w/w) | PHB Productivity (mg/L/d) | Ref. |
|---|---|---|---|---|---|---|
| Arthrospira platensis | Zarrouk | Batch, photoheterotrophic, 0.5% w/v acetate, nitrogen deprivation | n.r | 19.2 ± 0.5 | 14.5 ± 0.7 | [53] |
| Zarrouk | Batch, mixotrophic, 0.1% w/v butyrate, nitrogen deprivation | n.r | 17.8 ± 0.7 | 12.5 ± 0.6 | [53] | |
| Zarrouk | Batch, photoheterotrophic, 0.75% w/v glucose, nitrogen deprivation | n.r | 15.4 ± 0.7 | 10.8 ± 0.8 | [53] | |
| Zarrouk | Batch, photoheterotrophic, 0.1% w/v propionate, nitrogen deprivation | n.r | 12.6 ± 0.5 | 8.9 ± 0.6 | [53] | |
| Zarrouk | Batch, mixotrophic, 3.0–6.14 g/L pure glycerol | 7.8 ± 3.3 to 4.2 ± 3.1 | 1.3 ± 0.6 to 1.1 ± 1.5 | 2.4–2.6 | [54] | |
| Zarrouk | Batch, mixotrophic, 3.0–6.14 g/L crude glycerol | 10.8 ± 4.3 to 2.7 ± 2.6 | 0.06 ± 0.001 to 0.5 ± 0.11 | 0.09–1.0 | [54] | |
| Zarrouk | Batch, mixotrophic, acetate, formate, glycerol, oxalate, air ± CO2 | 11.0–14.5 | 0.1–3.0 | 0.2–7.8 | [35] | |
| Zarrouk | Fed batch, mixotrophic, acetate | 8.4 ± 0.1 | 1.5 ± 0.5 | 3.6 ± 1.1 | This study | |
| Zarrouk | Fed batch, mixotrophic, oxalate | 9.8 ± 0.1 | 0 | 0 | This study | |
| Zarrouk | Fed batch, mixotrophic, glycerol | 4.3 ± 0.2 | 7.8 ± 1.5 | 11.1 ± 1.7 | This study | |
| Zarrouk | Fed batch, mixotrophic, glycerol + oxalate | 6.3 + 0.3 | 1.5 ± 0.2 | 3.6 ± 0.7 | This study | |
| Zarrouk | Fed batch, mixotrophic, glycerol + oxalate + acetate | 6.9 + 0.2 | 1.9 ± 0.1 | 3.9 ± 0.7 | This study | |
| Spirulina sp. LEB18 | Biopolymer extraction waste | Batch, mixotrophic, 25% v/v waste | n.r | 10.6 | 4.7 | [59] |
| Nostoc muscorum | ES | Batch, mixotrophic, 1% w/v glucose + 1% w/v acetate, phosphate deprivation | n.r | 16.4 + 2.7 | 6.5 | [60] |
| Scytonema geitleri | Chu 10 | Batch, mixotrophic, 30 mM acetate | 7.1 | n.r | [61] | |
| Synechocystis sp. PCC6803-OEphaAB | BG 11 + 30 g/L chloramphenicol | Batch, mixotrophic, 4 mM acetate, nitrogen deprivation | n.r | 35 | n.r | [62] |
| Synechocystis sp. PCC6803-ΔSphU | shrimp wastewater | Batch, mixotrophic | n.r | 32.5 + 1.7 | 12.7 | [63] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shayesteh, H.; Laird, D.W.; Hughes, L.J.; Nematollahi, M.A.; Kakhki, A.M.; Moheimani, N.R. Co-Producing Phycocyanin and Bioplastic in Arthrospira platensis Using Carbon-Rich Wastewater. BioTech 2023, 12, 49. https://doi.org/10.3390/biotech12030049
Shayesteh H, Laird DW, Hughes LJ, Nematollahi MA, Kakhki AM, Moheimani NR. Co-Producing Phycocyanin and Bioplastic in Arthrospira platensis Using Carbon-Rich Wastewater. BioTech. 2023; 12(3):49. https://doi.org/10.3390/biotech12030049
Chicago/Turabian StyleShayesteh, Hajar, Damian W. Laird, Leonie J. Hughes, Mohammad A. Nematollahi, Amin Mirshamsi Kakhki, and Navid R. Moheimani. 2023. "Co-Producing Phycocyanin and Bioplastic in Arthrospira platensis Using Carbon-Rich Wastewater" BioTech 12, no. 3: 49. https://doi.org/10.3390/biotech12030049