
Mass Cultivation of Microalgae: II. A Large Species Pulsing Blue Light Concept

Abstract
:1. Introduction
2. Materials and Methods
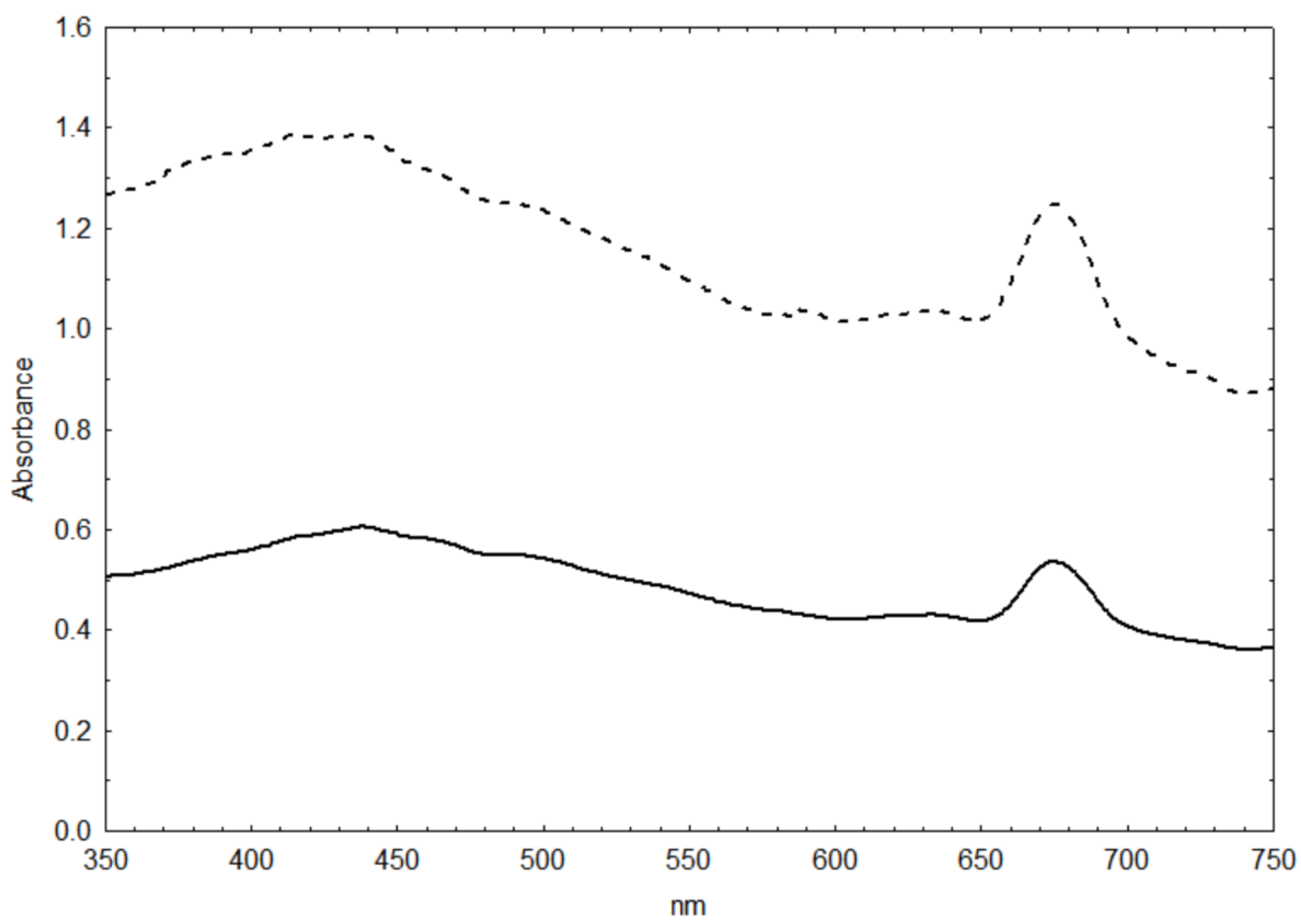
2.1. Dry Weight vs. Cell Size vs. Biovolume-Specific Absorbance
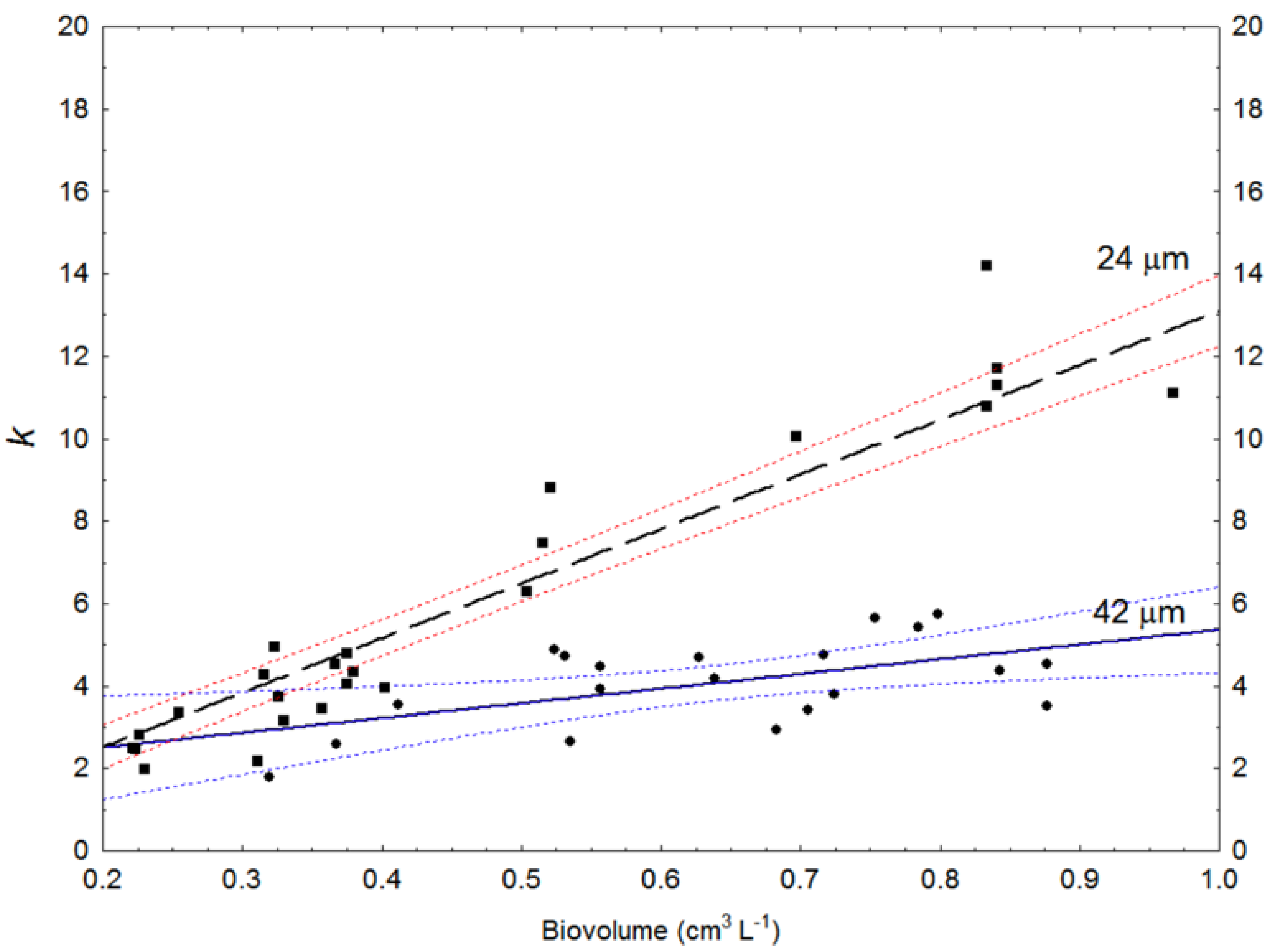
2.2. Diffuse Light Extinction vs. Cell Size
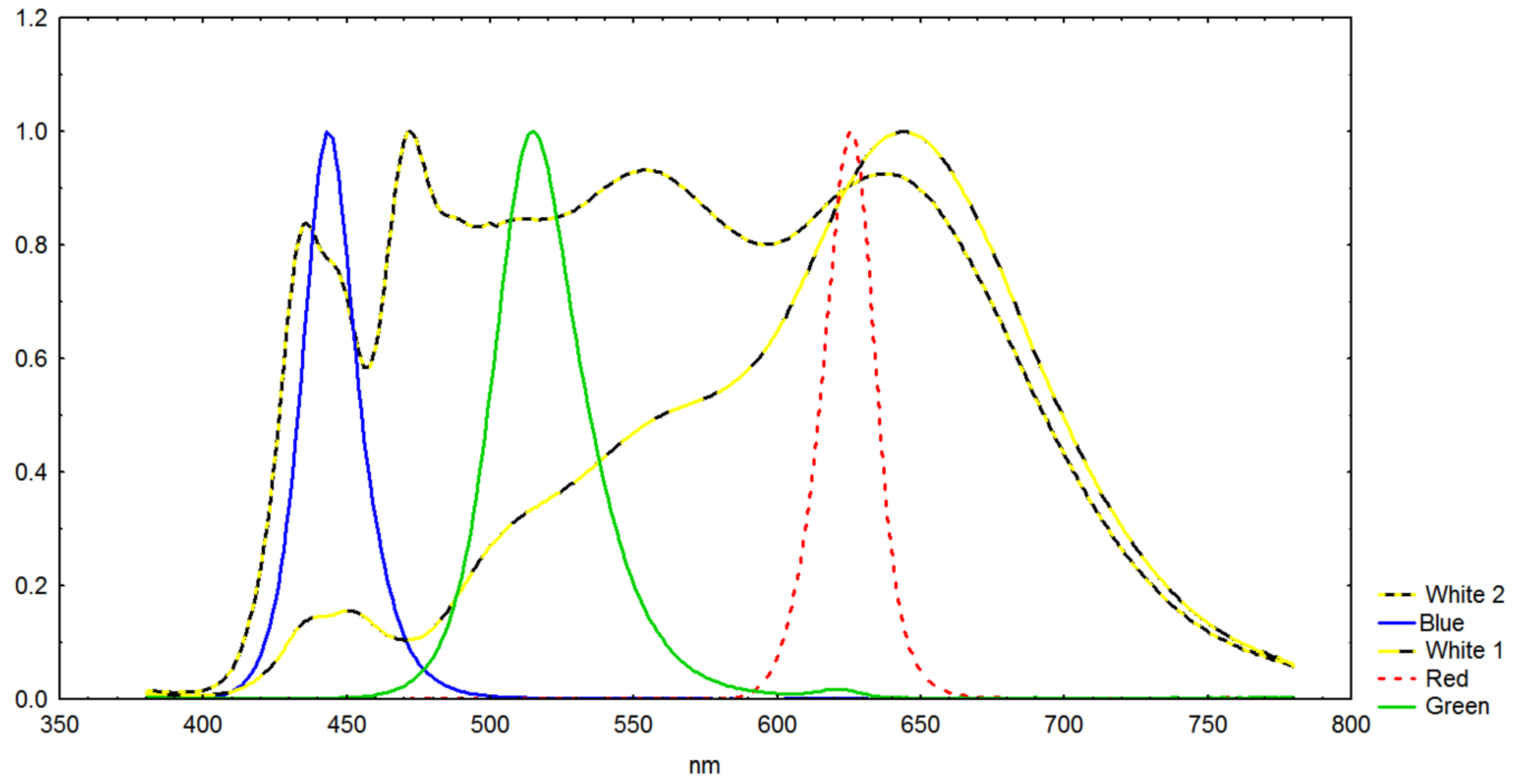
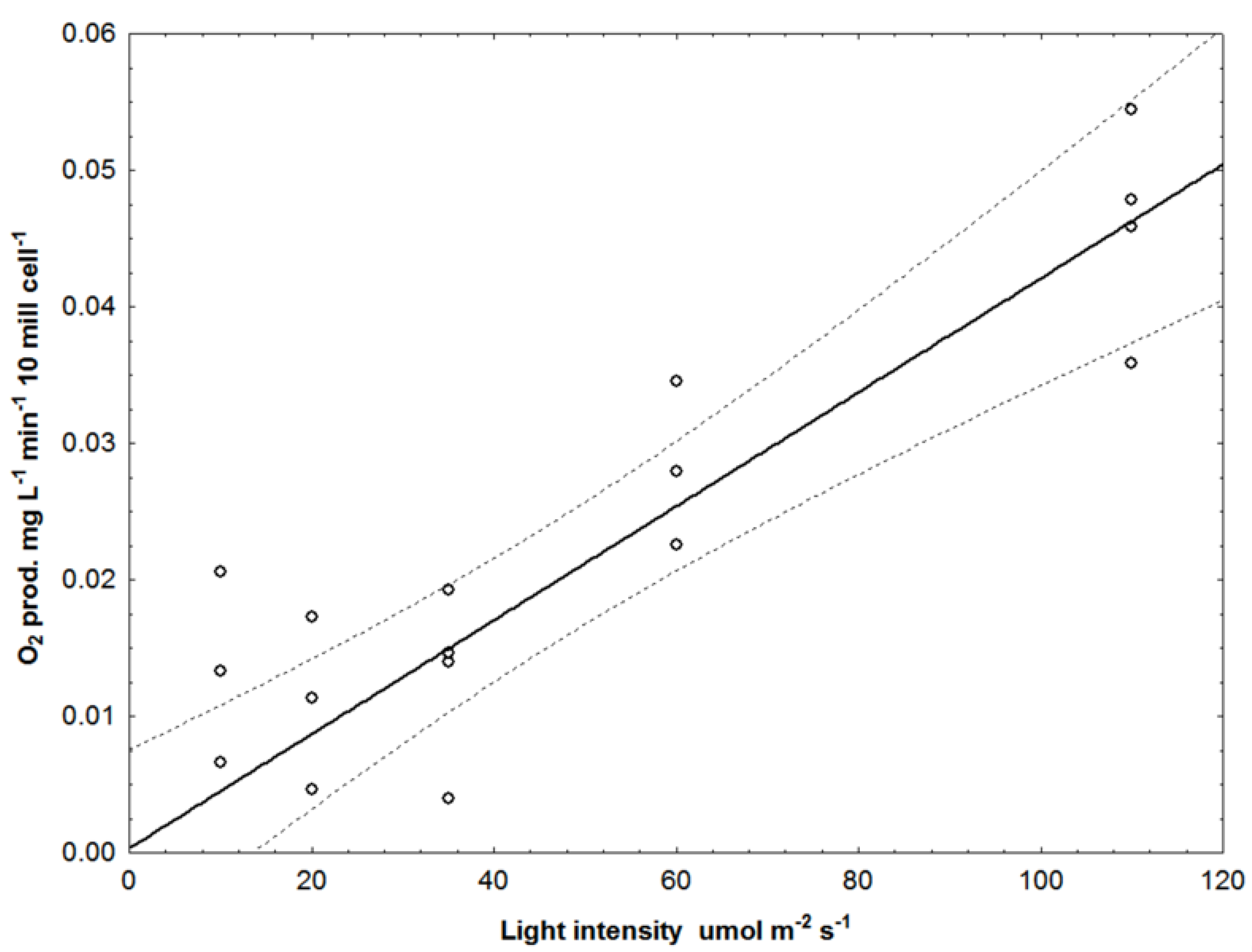
2.3. O2 Production vs. Spectrum vs. Linear or Pulsed Light
2.4. Lipid Class Analysis
3. Results
3.1. Dry Weight vs. Cell Size vs. Biomass-Specific Absorbance
3.2. Diffuse Light Extinction vs. Cell Size
3.3. O2 Production vs. Wavelength vs. Linear or Pulsed Light
- Mean O2 production for blue light was the highest and almost the same for linear and 100 Hz light.
- White 1 spectrum had high O2 production and alpha values with PWM light.
- Blue 100 Hz and linear light both had high O2 production and alpha values.
- Repeated exposure to the same linear and 100 Hz blue light intensities (Table 4) yielded higher O2 production values than the 3 min light gradient exposures.
3.4. Microalgae Growth with Blue Linear and Pulsed Light
3.5. Lipid Class Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beer, L.L.; Boyd, E.S.; Peters, J.W.; Posewitz, M.C. Engineering algae for biohydrogen and biofuel production. Curr. Opin. Biotechnol. 2009, 20, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Yadugiri, V. Milking diatoms–a new route to sustainable energy. Curr. Sci. 2009, 97, 748–750. Available online: https://www.jstor.org/stable/24112109 (accessed on 2 February 2023).
- Varfolomeev, S.; Wasserman, L. Microalgae as source of biofuel, food, fodder, and medicines. Appl. Biochem. Microbiol. 2011, 47, 789–807. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Lim, D.K.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Factories 2012, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.; Goswami, S. Microalgae—A green multi-product biorefinery for future industrial prospects. Biocatal. Agric. Biotechnol. 2020, 25, 101580. [Google Scholar] [CrossRef]
- Beigbeder, J.-B.; Sanglier, M.; de Medeiros Dantas, J.M.; Lavoie, J.-M. CO2 capture and inorganic carbon assimilation of gaseous fermentation effluents using Parachlorella kessleri microalgae. J. CO2 Util. 2021, 50, 101581. [Google Scholar] [CrossRef]
- Schiano di Visconte, G.; Spicer, A.; Chuck, C.J.; Allen, M.J. The microalgae biorefinery: A perspective on the current status and future opportunities using genetic modification. Appl. Sci. 2019, 9, 4793. [Google Scholar] [CrossRef]
- Bibi, F.; Jamal, A.; Huang, Z.; Urynowicz, M.; Ali, M.I. Advancement and role of abiotic stresses in microalgae biorefinery with a focus on lipid production. Fuel 2022, 316, 123192. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Vonshak, A. Scaling up microalgal cultures to commercial scale. Eur. J. Phycol. 2017, 52, 407–418. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Borowitzka, M.A. High-value products from microalgae—Their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.-J.; Chang, J.-S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Sirakov, M.; Palmieri, M.; Iovinella, M.; Davis, S.J.; Petriccione, M.; di Cicco, M.R.; De Stefano, M.; Ciniglia, C. Cyanidiophyceae (Rhodophyta) Tolerance to Precious Metals: Metabolic Response to Palladium and Gold. Plants 2021, 10, 2367. [Google Scholar] [CrossRef]
- Molazadeh, M.; Ahmadzadeh, H.; Pourianfar, H.R.; Lyon, S.; Rampelotto, P.H. The use of microalgae for coupling wastewater treatment with CO2 biofixation. Front. Bioeng. Biotechnol. 2019, 7, 42. [Google Scholar] [CrossRef]
- Guduru, R.K.; Gupta, A.A.; Dixit, U. Biological processes for CO2 capture. In Emerging Carbon Capture Technologies; Khalid, M., Dharaskar, S.A., Sillanpää, M., Siddiqui, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 371–400. [Google Scholar]
- Eilertsen, H.C.; Eriksen, G.K.; Bergum, J.-S.; Strømholt, J.; Elvevoll, E.; Eilertsen, K.-E.; Heimstad, E.S.; Giæver, I.H.; Israelsen, L.; Svenning, J.B. Mass cultivation of microalgae: I. Experiences with vertical column airlift photobioreactors, diatoms and CO2 sequestration. Appl. Sci. 2022, 12, 3082. [Google Scholar] [CrossRef]
- Vonshak, A. Recent advances in microalgal biotechnology. Biotechnol. Adv. 1990, 8, 709–727. [Google Scholar] [CrossRef]
- Han, P.; Lu, Q.; Fan, L.; Zhou, W. A review on the use of microalgae for sustainable aquaculture. Appl. Sci. 2019, 9, 2377. [Google Scholar] [CrossRef]
- Beijierinck, M. Kulturversuche mit Zoochloren, Lichenenggonidien und anderen niederen Algen. Physis 1890, 48, 725–785. [Google Scholar]
- Kok, B. Experiments on photosynthesis by Chlorella in flashing light. In Algal Culture from Laboratory to Pilot Plant; Oxford University Press: Oxford, UK, 1953; pp. 63–75. [Google Scholar]
- Nethravathy, M.; Mehar, J.G.; Mudliar, S.N.; Shekh, A.Y. Recent advances in microalgal bioactives for food, feed, and healthcare products: Commercial potential, market space, and sustainability. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1882–1897. [Google Scholar] [CrossRef]
- García, J.L.; De Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef]
- Gjøsund, S.H.; Skjermo, J.; Forbord, S.; Jafarzadeh, S.; Sletta, H.; Aasen, I.M.; Hagemann, A.; Chauton, M.S.; Aursand, I.G.; Evjemo, J.O. Bærekraftig Fôr Til Norsk Laks; Sintef Ocean: Trondheim, Norway, 2020; p. 117. [Google Scholar]
- Mohsenpour, S.F.; Hennige, S.; Willoughby, N.; Adeloye, A.; Gutierrez, T. Integrating micro-algae into wastewater treatment: A review. Sci. Total Environ. 2021, 752, 142168. [Google Scholar] [CrossRef] [PubMed]
- Fozer, D.; Kiss, B.; Lorincz, L.; Szekely, E.; Mizsey, P.; Nemeth, A. Improvement of microalgae biomass productivity and subsequent biogas yield of hydrothermal gasification via optimization of illumination. Renew. Energy 2019, 138, 1262–1272. [Google Scholar] [CrossRef]
- Lim, Y.A.; Chong, M.N.; Foo, S.C.; Ilankoon, I. Analysis of direct and indirect quantification methods of CO2 fixation via microalgae cultivation in photobioreactors: A critical review. Renew. Sustain. Energy Rev. 2021, 137, 110579. [Google Scholar] [CrossRef]
- Ugwu, C.; Aoyagi, H.; Uchiyama, H. Photobioreactors for mass cultivation of algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef] [PubMed]
- Blanken, W.; Cuaresma, M.; Wijffels, R.H.; Janssen, M. Cultivation of microalgae on artificial light comes at a cost. Algal Res. 2013, 2, 333–340. [Google Scholar] [CrossRef]
- Ranglová, K.; Bureš, M.; Manoel, J.C.; Lakatos, G.E.; Masojídek, J. Efficient microalgae feed production for fish hatcheries using an annular column photobioreactor characterized by a short light path and central LED illumination. J. Appl. Phycol. 2022, 34, 31–41. [Google Scholar] [CrossRef]
- Richmond, A. Biological principles of mass cultivation of photoautotrophic microalgae. In Handbook of Microalgal Culture: Applied Phycology and Biotechnology; Richmond, A., Hu, Q., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2013; pp. 169–204. [Google Scholar]
- Geider, R.; Osborne, B. Light absorption by a marine diatom: Experimental observations and theoretical calculations of the package effect in a small Thalassiosira species. Mar. Biol. 1987, 96, 299–308. [Google Scholar] [CrossRef]
- Marañón, E. Phytoplankton size structure. In Marine Biology: A Derivative of the Encyclopedia of Ocean Sciences, 2nd ed.; Steele, J., Thorpe, A., Turekian, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; Volume 85. [Google Scholar]
- Sinclair, D. Light scattering by spherical particles. JOSA 1947, 37, 475–480. [Google Scholar] [CrossRef]
- Baker, E.T.; Lavelle, J.W. The effect of particle size on the light attenuation coefficient of natural suspensions. J. Geophys. Res. Ocean. 1984, 89, 8197–8203. [Google Scholar] [CrossRef]
- Agustí, S. Light environment within dense algal populations: Cell size influences on self-shading. J. Plankton Res. 1991, 13, 863–871. [Google Scholar] [CrossRef]
- Nelson, N.B.; Prézelin, B.B.; Bidigare, R.R. Phytoplankton light absorption and the package effect in California coastal waters. Mar. Ecol. Prog. Ser. 1993, 94, 217–227. Available online: https://www.jstor.org/stable/24832708 (accessed on 2 February 2023). [CrossRef]
- Tennessen, D.J.; Bula, R.J.; Sharkey, T.D. Efficiency of photosynthesis in continuous and pulsed light emitting diode irradiation. Photosynth. Res. 1995, 44, 261–269. [Google Scholar] [CrossRef]
- Lunka, A.A.; Bayless, D.J. Effects of flashing light-emitting diodes on algal biomass productivity. J. Appl. Phycol. 2013, 25, 1679–1685. [Google Scholar] [CrossRef]
- Schulze, P.S.; Guerra, R.; Pereira, H.; Schüler, L.M.; Varela, J.C. Flashing LEDs for microalgal production. Trends Biotechnol. 2017, 35, 1088–1101. [Google Scholar] [CrossRef]
- Eilertsen, H.C.; Holm-Hansen, O. Effects of high latitude UV radiation on phytoplankton and nekton modelled from field measurements by simple algorithms. PolarRes 2000, 19, 173–182. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplanktonmethodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar]
- Hasle, G.R.; Syvertsen, E.E. Marine Diatoms. In Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: Cambridge, MA, USA, 1997; pp. 5–361. [Google Scholar]
- Menden-Deuer, S.; Lessard, E.J. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton. Limnol. Oceanogr. 2000, 45, 569–579. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine plankton diatoms. I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Jensen, I.; Mæhre, H.; Tømmerås, S.; Eilertsen, K.; Olsen, R.; Elvevoll, E. Farmed Atlantic salmon (Salmo salar L.) is a good source of long chain omega-3 fatty acids. Nutr. Bull. 2012, 37, 25–29. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Cequier-Sánchez, E.; Rodriguez, C.; Ravelo, A.G.; Zarate, R. Dichloromethane as a solvent for lipid extraction and assessment of lipid classes and fatty acids from samples of different natures. J. Agric. Food Chem. 2008, 56, 4297–4303. [Google Scholar] [CrossRef]
- Abreu, S.; Solgadi, A.; Chaminade, P. Optimization of normal phase chromatographic conditions for lipid analysis and comparison of associated detection techniques. J. Chromatogr. 2017, 1514, 54–71. [Google Scholar] [CrossRef]
- Platt, T.; Jassby, A.D. The relationship between photosynthesis and light for natural assemblages of coastal marine phytoplankton 1. J. Phycol. 1976, 12, 421–430. [Google Scholar] [CrossRef]
- Platt, T.; Gallegos, C.; Harrison, W.G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. J. Mar. Res. 1982, 38, 687–701. [Google Scholar]
- Yale, C.; Forsythe, A.B. Winsorized regression. Technometrics 1976, 18, 291–300. [Google Scholar] [CrossRef]
- Malerba, M.E.; Palacios, M.M.; Palacios Delgado, Y.M.; Beardall, J.; Marshall, D.J. Cell size, photosynthesis and the package effect: An artificial selection approach. New Phytol. 2018, 219, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Marañón, E. Cell size as a key determinant of phytoplankton metabolism and community structure. Annu. Rev. Mar. Sci. 2015, 7, 241–264. [Google Scholar] [CrossRef] [PubMed]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis, 2nd ed.; Stevenson, J.C., Ed.; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972. [Google Scholar]
- Webb, W.L.; Newton, M.; Starr, D. Carbon dioxide exchange of Alnus rubra: A mathematical model. Oecologia 1974, 17, 281–291. [Google Scholar] [CrossRef]
- Dubinsky, Z.; Falkowski, P.G.; Post, A.F.; Van Hes, U.M. A system for measuring phytoplankton photosynthesis in a defined light field with an oxygen electrode. J. Plankton Res. 1987, 9, 607–612. [Google Scholar] [CrossRef]
- Ryther, J.; Vaccaro, R. A comparison of the oxygen and 14C methods of measuring marine photosynthesis. ICES J. Mar. Sci. 1954, 20, 25–34. [Google Scholar] [CrossRef]
- Williams, P.; Raine, R. Agreement between the c-14 and oxygen methods of measuring phytoplankton production-reassessment of the photosynthetic quotient. Oceanol. Acta 1979, 2, 411–416. [Google Scholar]
- Trampe, E.; Hansen, P.J.; Kuhl, M. A comparison of photosynthesis measurements by O2 evolution, 14 C assimilation, and variable chlorophyll fluorescence during light acclimatization of the diatom Coscinodiscus granii. Algae 2015, 30, 103–119. [Google Scholar]
- Kim, N.-J.; Lee, C.-G. A theoretical consideration on oxygen production rate in microalgal cultures. Biotechnol. Bioprocess Eng. 2001, 6, 352–358. [Google Scholar] [CrossRef]
- Palanisamy, K.M.; Rahim, M.H.A.; Govindan, N.; Ramaraj, R.; Kuppusamy, P.; Maniam, G.P. Effect of blue light intensity and photoperiods on the growth of diatom Thalassiosira pseudonana. Bioresour. Technol. Rep. 2022, 19, 101152. [Google Scholar] [CrossRef]
- Wagner, H.; Jakob, T.; Wilhelm, C. Balancing the energy flow from captured light to biomass under fluctuating light conditions. New Phytol. 2006, 169, 95–108. [Google Scholar] [CrossRef]
- del Pilar Sánchez-Saavedra, M.; Voltolina, D. Effect of blue-green light on growth rate and chemical composition of three diatoms. J. Appl. Phycol. 1996, 8, 131–137. [Google Scholar] [CrossRef]
- Schellenberger Costa, B.; Jungandreas, A.; Jakob, T.; Weisheit, W.; Mittag, M.; Wilhelm, C. Blue light is essential for high light acclimation and photoprotection in the diatom Phaeodactylum tricornutum. J. Exp. Bot. 2013, 64, 483–493. [Google Scholar] [CrossRef]
- Stauber, J.L.; Jeffrey, S. Photosynthetic pigments in fifty—One species of marine diatoms. J. Phycol. 1988, 24, 158–172. [Google Scholar] [CrossRef]
- Ogbonna, J.C.; Yada, H.; Tanaka, H. Light supply coefficient: A new engineering parameter for photobioreactor design. J. Ferment. Bioeng. 1995, 80, 369–376. [Google Scholar] [CrossRef]
- Perner-Nochta, I.; Posten, C. Simulations of light intensity variation in photobioreactors. J. Biotechnol. 2007, 131, 276–285. [Google Scholar] [CrossRef]
- Nymark, M.; Valle, K.C.; Brembu, T.; Hancke, K.; Winge, P.; Andresen, K.; Johnsen, G.; Bones, A.M. An integrated analysis of molecular acclimation to high light in the marine diatom Phaeodactylum tricornutum. PLoS ONE 2009, 4, e7743. [Google Scholar] [CrossRef]
- Gilstad, M.; Sakshaug, E. Growth rates of ten diatom species from the Barents Sea at different irradiances and day lengths. Mar. Ecol. Prog. Ser. 1990, 64, 169–173. [Google Scholar] [CrossRef]
- Croteau, D.; Lacour, T.; Schiffrine, N.; Morin, P.I.; Forget, M.H.; Bruyant, F.; Ferland, J.; Lafond, A.; Campbell, D.A.; Tremblay, J.É. Shifts in growth light optima among diatom species support their succession during the spring bloom in the Arctic. J. Ecol. 2022, 110, 1356–1375. [Google Scholar] [CrossRef]
- Falkowski, P.G. Light-shade adaptation in marine phytoplankton. In Primary Productivity in the Sea; Springer: Berlin/Heidelberg, Germany, 1980; pp. 99–119. [Google Scholar]
- Lavaud, J. Fast regulation of photosynthesis in diatoms: Mechanisms, evolution and ecophysiology. Funct. Plant Sci. Biotechonology 2007, 1, 267–287. [Google Scholar]
- Rascher, U.; Nedbal, L. Dynamics of photosynthesis in fluctuating light. Curr. Opin. Plant Biol. 2006, 9, 671–678. [Google Scholar] [CrossRef]
- Smith, R.C.; Wilson, W.H. Photon scalar irradiance. Appl. Opt. 1972, 11, 934–938. [Google Scholar] [CrossRef]
- Eppley, R.W. Temperature and phytoplankton growth in the sea. Fish. Bull. 1972, 70, 1063–1085. [Google Scholar]
- Huseby, S.; Degerlund, M.; Eriksen, G.K.; Ingebrigtsen, R.A.; Eilertsen, H.C.; Hansen, E. Chemical diversity as a function of temperature in six northern diatom species. Mar. Drugs 2013, 11, 4232–4245. [Google Scholar] [CrossRef]
- Iluz, D.; Alexandrovich, I.; Dubinsky, Z. The enhancement of photosynthesis by fluctuating light. Artif. Photosynth. 2012, 288, 116–133. [Google Scholar]
- Terry, K.L. Photosynthesis in modulated light: Quantitative dependence of photosynthetic enhancement on flashing rate. Biotechnol. Bioeng. 1986, 28, 988–995. [Google Scholar] [CrossRef]
- Vejrazka, C.; Janssen, M.; Streefland, M.; Wijffels, R.H. Photosynthetic efficiency of Chlamydomonas reinhardtii in flashing light. Biotechnol. Bioeng. 2011, 108, 2905–2913. [Google Scholar] [CrossRef]
- Roselló Sastre, R.M. Kopplung Physiologischer und Verfahrenstechnischer Parameter Beim Wachstum und Bei der Produktbildung der Rotalge Porphyridium Purpureum; KIT Scientific Publishing: Karlsruhe, Germany, 2010. [Google Scholar]
- Mouget, J.-L.; de la Noë, J.; Legendre, L.; Jean, Y.; Viarouge, P. Long-term acclimatization of Scenedesmus bicellularis to high-frequency intermittent lighting (100 Hz). I. Growth, photosynthesis and photosystem II activity. J. Plankton Res. 1995, 17, 859–874. [Google Scholar] [CrossRef]
- Goss, R.; Lepetit, B. Biodiversity of NPQ. J. Plant Physiol. 2015, 172, 13–32. [Google Scholar] [CrossRef]
- Hüner, N.P.; Bode, R.; Dahal, K.; Hollis, L.; Rosso, D.; Krol, M.; Ivanov, A.G. Chloroplast redox imbalance governs phenotypic plasticity: The “grand design of photosynthesis” revisited. Front. Plant Sci. 2012, 3, 255. [Google Scholar] [CrossRef]
- Allahverdiyeva, Y.; Suorsa, M.; Tikkanen, M.; Aro, E.-M. Photoprotection of photosystems in fluctuating light intensities. J. Exp. Bot. 2015, 66, 2427–2436. [Google Scholar] [CrossRef]
- Laufs, U.; Parhofer, K.G.; Ginsberg, H.N.; Hegele, R.A. Clinical review on triglycerides. Eur. Heart J. 2020, 41, 99c–109c. [Google Scholar] [CrossRef]
- Bovet, P.; Faeh, D.; Madeleine, G.; Viswanathan, B.; Paccaud, F. Decrease in blood triglycerides associated with the consumption of eggs of hens fed with food supplemented with fish oil. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 280–287. [Google Scholar] [CrossRef]
- Olvera-Gonzalez, E.; Escalante-Garcia, N.; Myers, D.; Ampim, P.; Obeng, E.; Alaniz-Lumbreras, D.; Castaño, V. Pulsed led-lighting as an alternative energy savings technique for vertical farms and plant factories. Energies 2021, 14, 1603. [Google Scholar] [CrossRef]
- Valegård, K.; Andralojc, P.J.; Haslam, R.P.; Pearce, F.G.; Eriksen, G.K.; Madgwick, P.J.; Kristoffersen, A.K.; van Lun, M.; Klein, U.; Eilertsen, H.C. Structural and functional analyses of Rubisco from arctic diatom species reveal unusual posttranslational modifications. J. Biol. Chem. 2018, 293, 13033–13043. [Google Scholar] [CrossRef]
- Barry, A.N.; Starkenburg, S.R.; Sayre, R.T. Strategies for optimizing algal biology for enhanced biomass production. Front. Energy Res. 2015, 3, 1. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Diameter/S.D. μm | Cell Height/S.D. μm | Cell Concentration (Cells L−1) | Single Cell Biovolume (μm3) | Culture Biovolume-Specific DW (g cm−3) |
|---|---|---|---|---|
| 17.045/1.15 | 30.758/1.27 | 8,922,760 | 7070 | 0.179 |
| 33.636/7.42 | 20.985/3.23 | 11,457,635 | 18,703 | 0.151 |
| Mean Cell Width (μm) | Biovolume (cm3 L−1) | Light Intensity μmol m−2 s−1 at Depth | ||
|---|---|---|---|---|
| 0 m | 0.5 m | 1.0 m | ||
| 24 | 0.4 | 100 | 8.16 | 0.66 |
| 42 | 0.4 | 100 | 21.23 | 4.59 |
| 24 | 0.8 | 100 | 0.67 | 0.005 |
| 42 | 0.8 | 100 | 10.53 | 1.11 |
| Spectra | Unit–O2 Production | Linear | 1 Hz | 10 Hz | 50 Hz | 100 Hz | 500 Hz |
|---|---|---|---|---|---|---|---|
| Blue | mg O2 L−1 min−1 10 mill cell−1 | 0.022 | 0.016 | 0.012 | 0.012 | 0.020 | 0.016 |
| Green | 0.015 | 0.008 | 0.008 | 0.017 | 0.012 | 0.008 | |
| White 1 | 0.014 | 0.024 | 0.015 | 0.019 | 0.017 | 0.010 | |
| White 2 | 0.018 | 0.014 | 0.008 | 0.011 | 0.016 | 0.015 | |
| Red | 0.012 | 0.009 | 0.015 | 0.006 | 0.013 | 0.005 | |
| Unit–alpha values | |||||||
| Blue | mg O2 L−1 min−1 10 mill cell−1 μmol quanta m−2 s−1 | 0.00074 | 0.00056 | 0.00052 | 0.00027 | 0.00082 | 0.00055 |
| Green | 0.00064 | 0.00032 | 0.00028 | 0.00083 | 0.00039 | 0.00040 | |
| White 1 | 0.00052 | 0.00096 | 0.00054 | 0.00073 | 0.00077 | 0.00037 | |
| White 2 | 0.00059 | 0.00062 | 0.00037 | 0.00040 | 0.00067 | 0.00060 | |
| Red | 0.00036 | 0.00038 | 0.00048 | 0.00035 | 0.00036 | 0.00015 |
| Irradiance (μmol m−2 s−1) Blue Linear | n | Production (mg O2 L−1 min−1 10 mill Cell−1) | S.D. |
|---|---|---|---|
| 10 | 21 | 0.0337 | 0.0073 |
| 20 | 21 | 0.0433 | 0.0095 |
| 35 | 20 | 0.0467 | 0.0011 |
| 60 | 17 | 0.0508 | 0.0110 |
| 210 | 12 | 0.0591 | 0.0120 |
| Mean | 0.0467 | ||
| Blue PWM | |||
| 10 | 15 | 0.0374 | 0.0014 |
| 20 | 18 | 0.0485 | 0.0010 |
| 35 | 26 | 0.0461 | 0.0101 |
| 60 | 21 | 0.0466 | 0.0110 |
| 210 | 14 | 0.0554 | 0.0150 |
| Mean | 0.0468 | ||
| Experiment | Doublings Day−1 | Temperature (°C) | Oxygen (% Saturation) | pH |
|---|---|---|---|---|
| L1 | 1.23 (0.75) | 9.64 (1.67) | 99.2 (1.13) | 8.42 |
| L2 | 1.08 (0.70) | 9.65 (1.09) | 101.8 (2.20) | 8.50 |
| L3 | 0.72 (0.50) | 8.86 (1.38) | 99.6 (1.77) | 8.48 |
| Mean Linear | 1.01 | |||
| P1 | 1.09 (0.34) | 10.56 (0.86) | 111.6 (1.16) | 8.21 |
| P2 | 0.83 (0.33) | 10.04 (1.03) | 100.78 (0.48) | 8.46 |
| P3 | 1.00 (0.35) | 8.77 (1.33) | 100.33 (4.59) | 8.45 |
| Mean PWM | 0.97 |
| L1 | L2 | L3 | P1 | P2 | P3 | |
|---|---|---|---|---|---|---|
| TAG | 62.7 | 79.6 | 67.5 | 28.6 | 13.3 | 14.3 |
| FAIc | 2.2 | 0.0 | 0.3 | 0.8 | 0.5 | 0.3 |
| DAG | 3.8 | 2.9 | 4.4 | 18.7 | 14.5 | 13.8 |
| FFA | 0.9 | 0.4 | 1.1 | 3.4 | 1.6 | 1.9 |
| MAG | 0.4 | 0.1 | 0.2 | 2.6 | 0.9 | 1.0 |
| MGDG | 26.8 | 15.2 | 23.7 | 36.9 | 62.5 | 64.1 |
| DGDG | 3.2 | 1.7 | 2.9 | 8.9 | 6.8 | 4.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eilertsen, H.C.; Strømholt, J.; Bergum, J.-S.; Eriksen, G.K.; Ingebrigtsen, R. Mass Cultivation of Microalgae: II. A Large Species Pulsing Blue Light Concept. BioTech 2023, 12, 40. https://doi.org/10.3390/biotech12020040
Eilertsen HC, Strømholt J, Bergum J-S, Eriksen GK, Ingebrigtsen R. Mass Cultivation of Microalgae: II. A Large Species Pulsing Blue Light Concept. BioTech. 2023; 12(2):40. https://doi.org/10.3390/biotech12020040
Chicago/Turabian StyleEilertsen, Hans Chr., Jo Strømholt, John-Steinar Bergum, Gunilla Kristina Eriksen, and Richard Ingebrigtsen. 2023. "Mass Cultivation of Microalgae: II. A Large Species Pulsing Blue Light Concept" BioTech 12, no. 2: 40. https://doi.org/10.3390/biotech12020040