
A Bacterial Myeloperoxidase with Antimicrobial Properties

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Procedures
2.2.1. Genetic Constructions
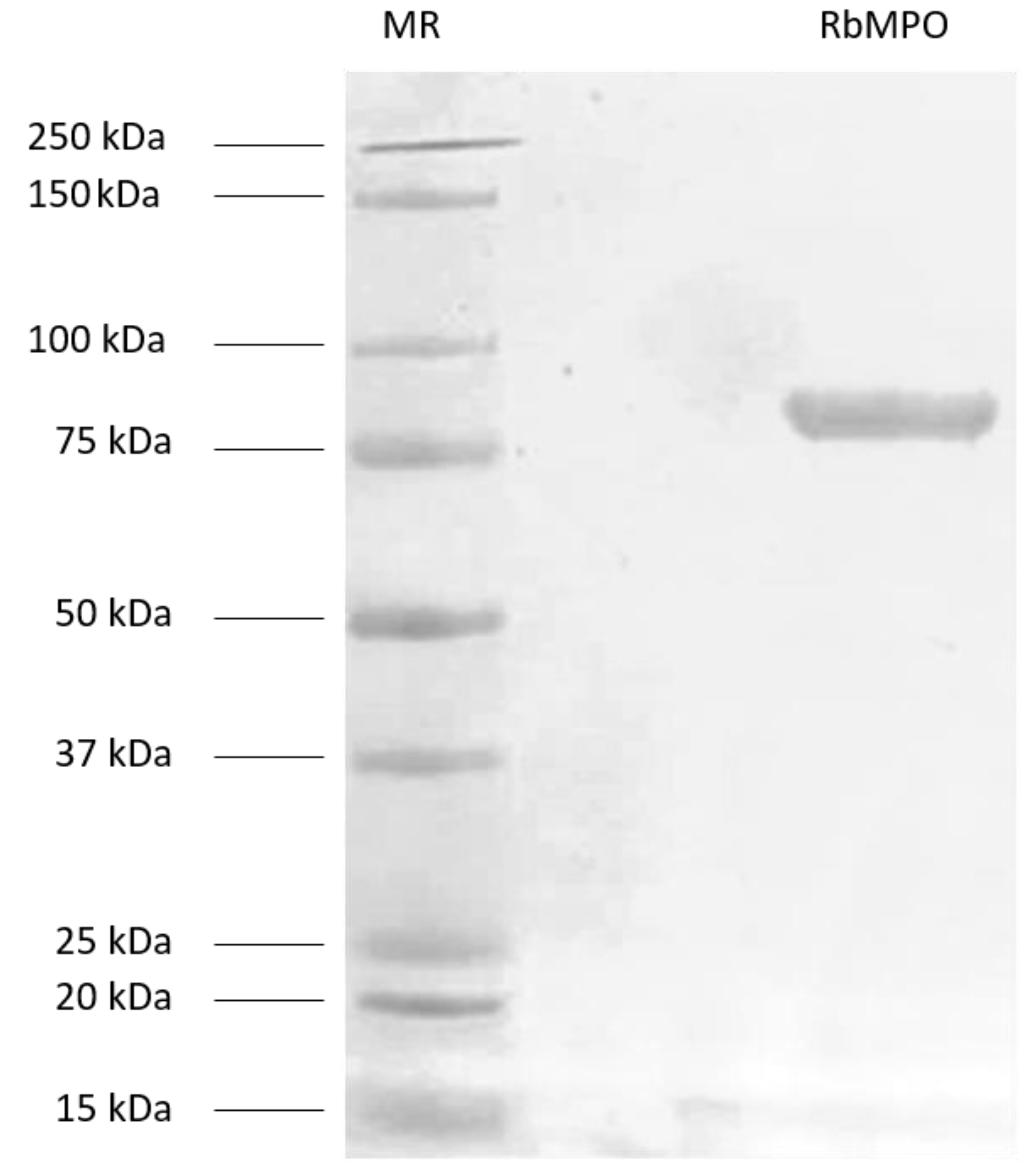
2.2.2. Production and Purification of RbMPO
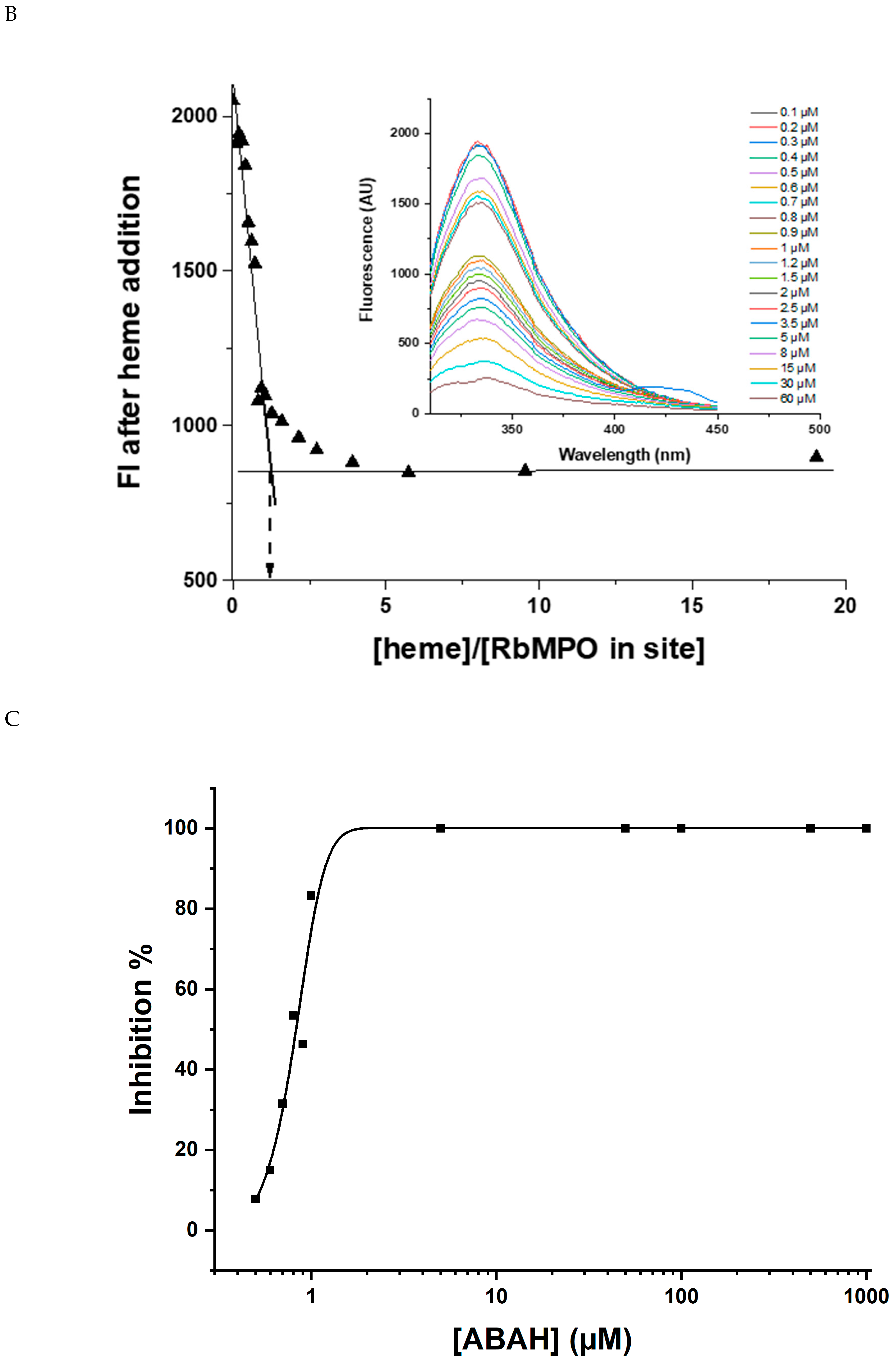
2.2.3. Site Titration of RbMPO by Heme
2.2.4. Inhibition of the Enzyme by ABAH
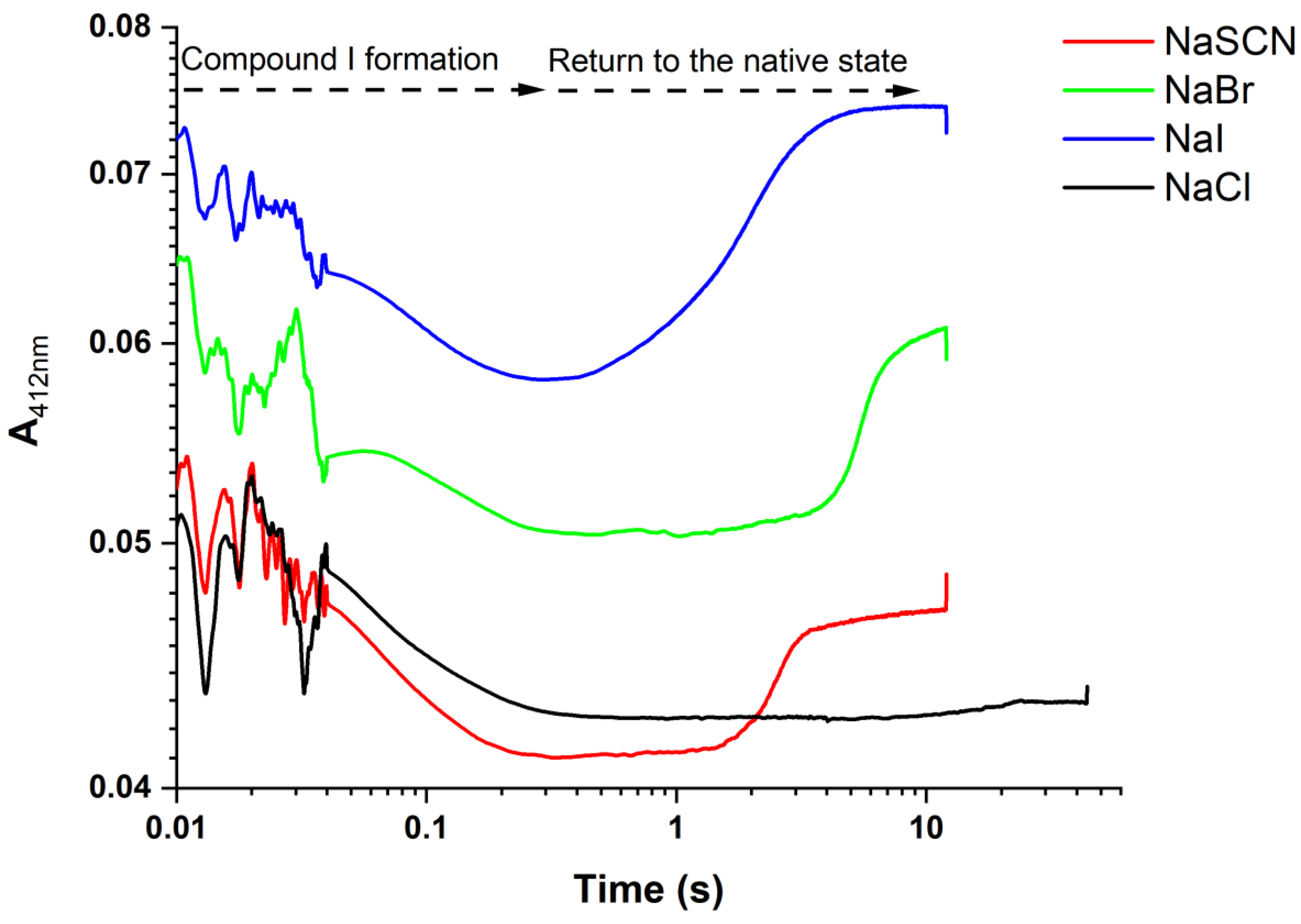
2.2.5. Regeneration of RbMPO under Native Form by Various Salts
2.2.6. Steady-State Kinetic Parameters of RbMPO
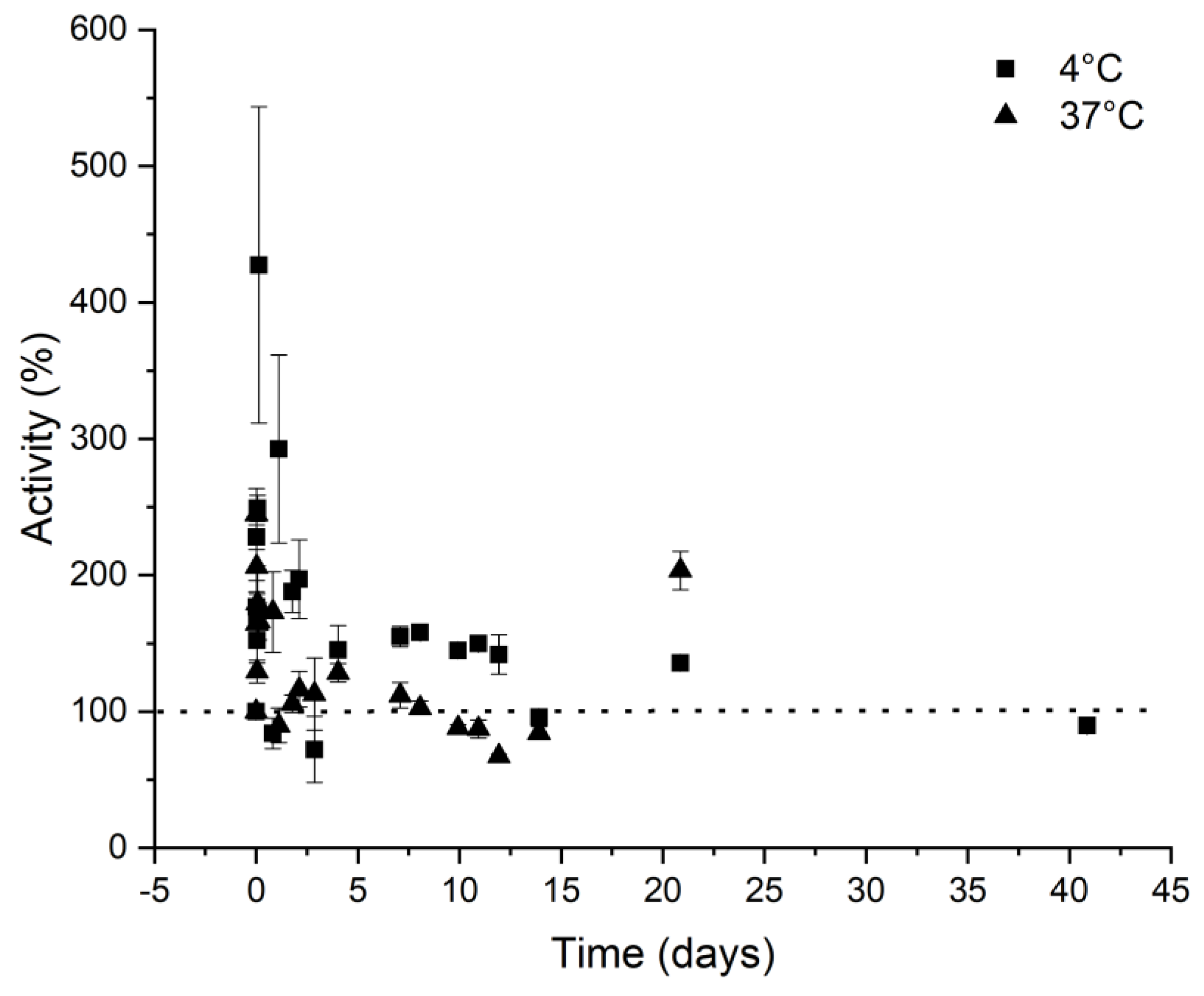
2.2.7. Stability of RbMPO at 4 and 37 °C
2.2.8. Catalase Activity
2.2.9. Peroxidase Activity
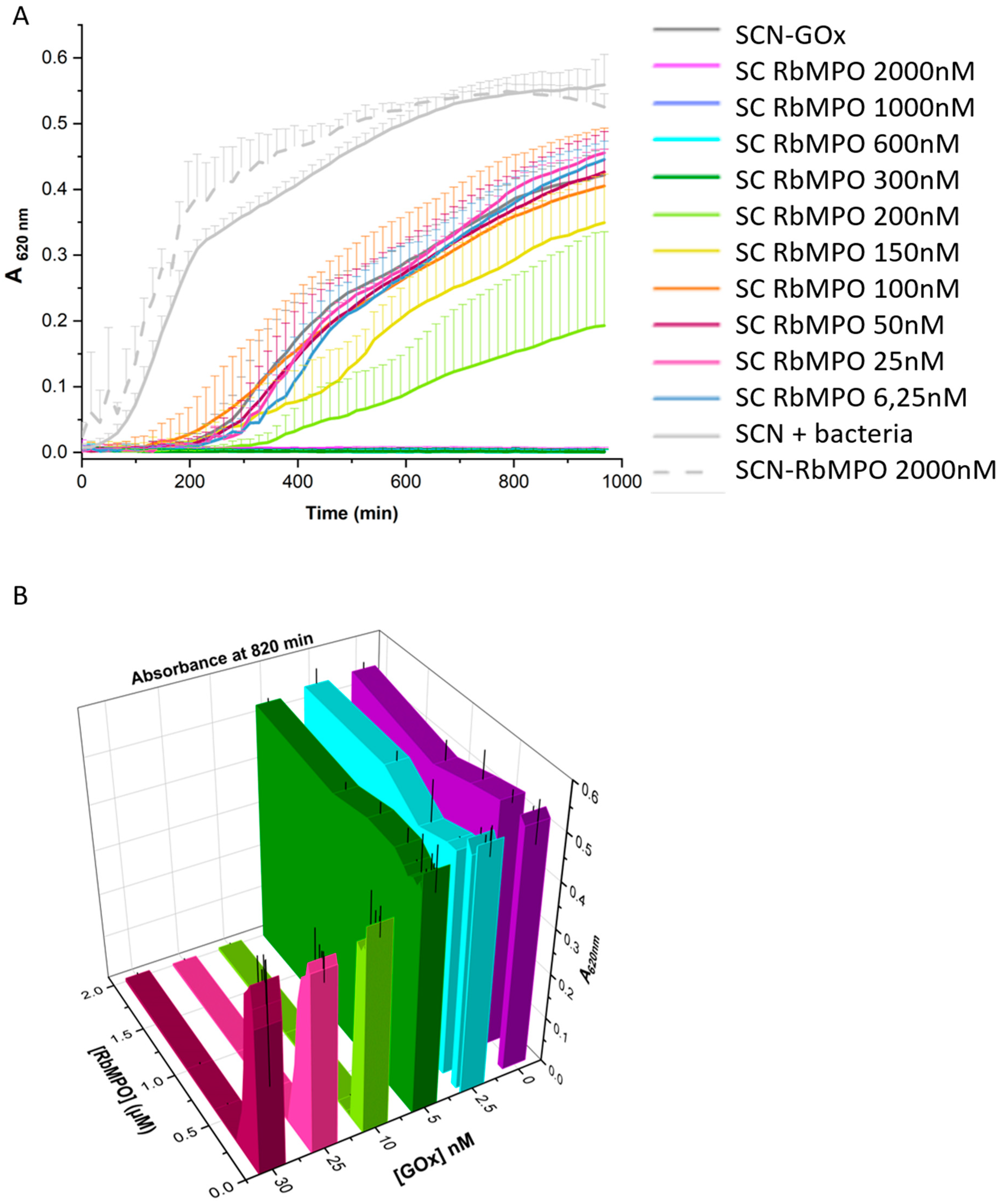
2.2.10. Microbicidal Effect on E. coli ATCC 25922
3. Results
3.1. Production and Purification of RbMPO
3.2. Reconstitution of the Active Site of RbMPO
3.3. Inhibition of the Enzyme by ABAH
3.4. Pre-Steady-State Kinetic Parameters of RbMPO
3.5. Steady-State Kinetic Parameters of RbMPO
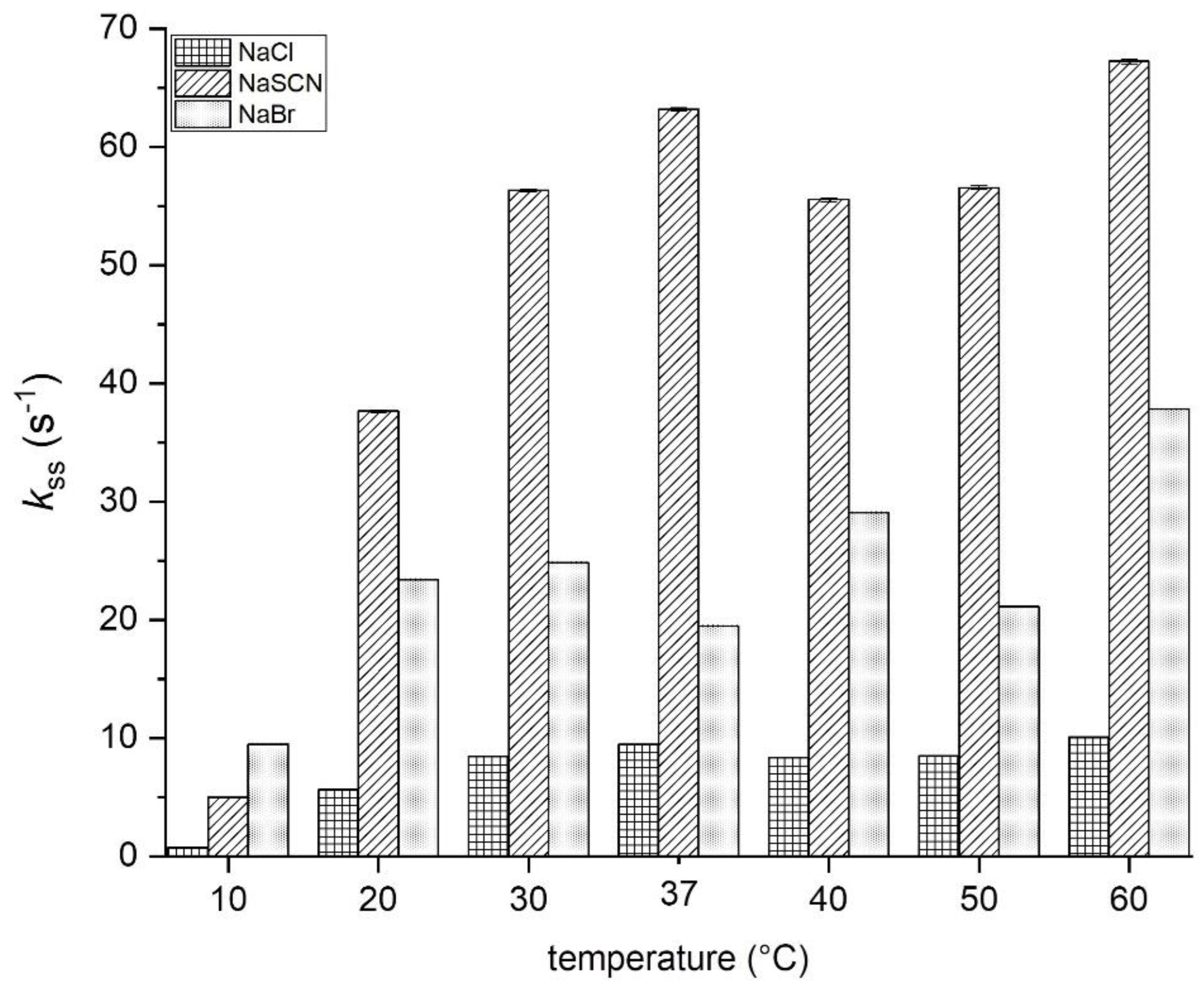
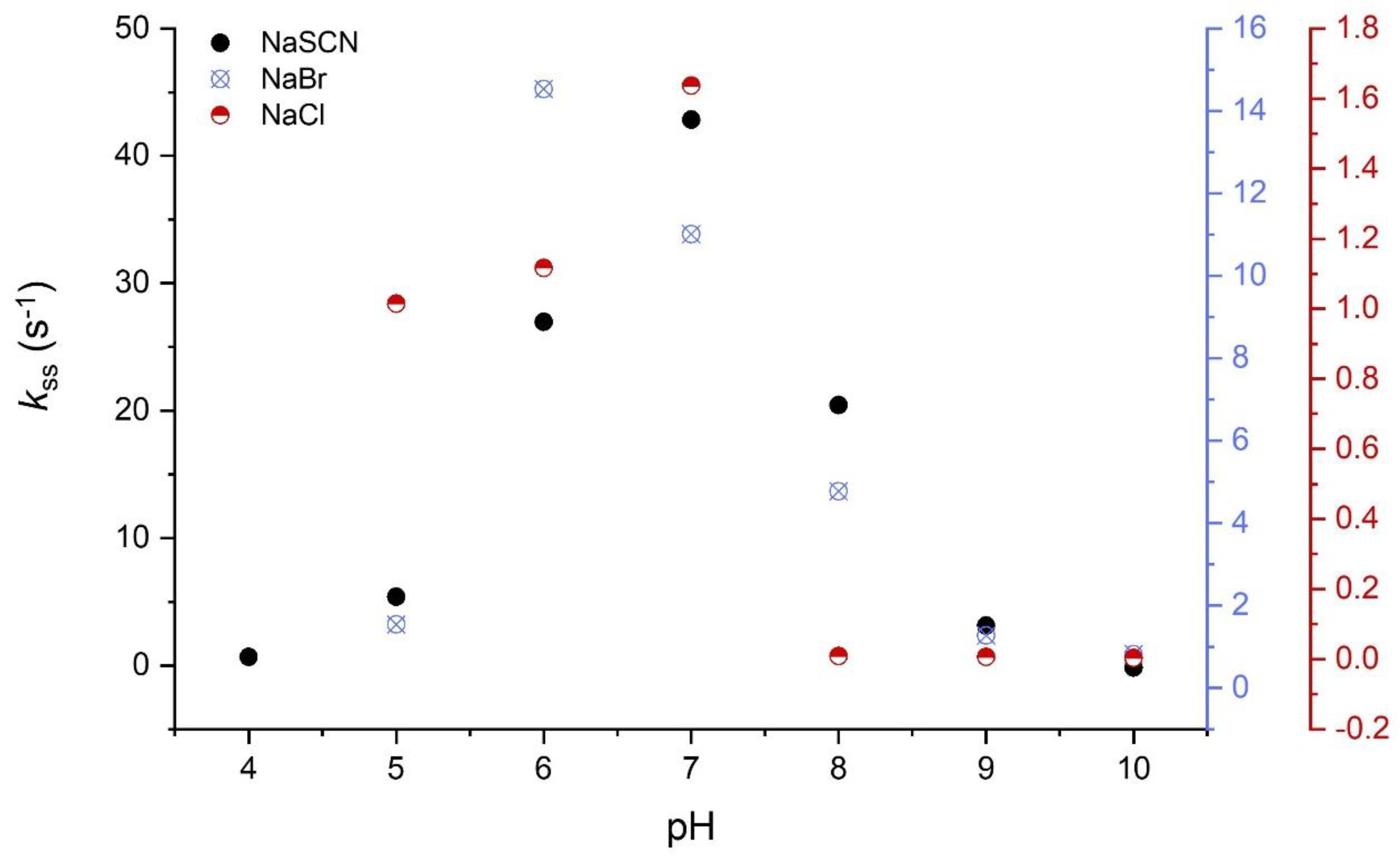
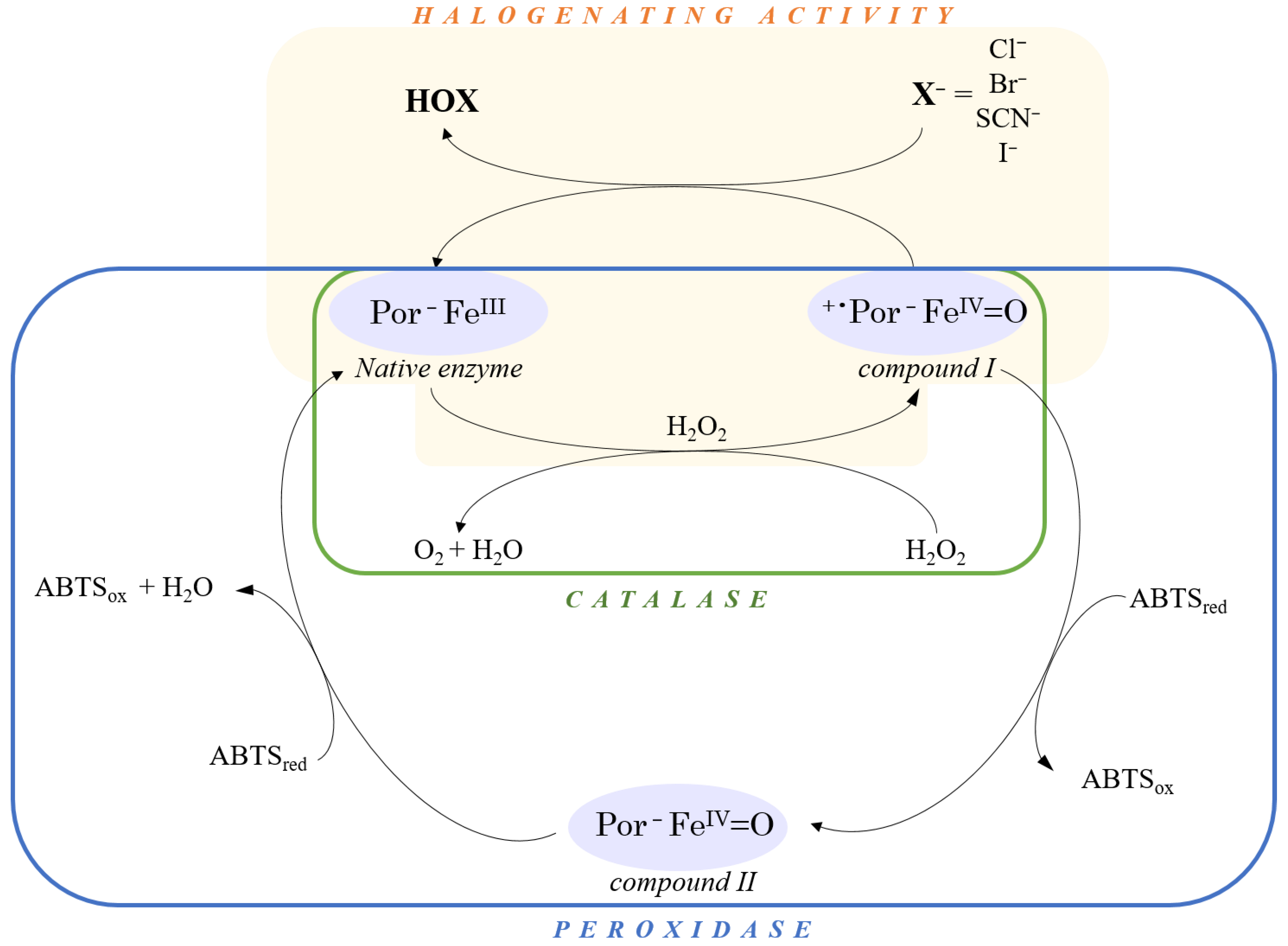
3.5.1. Halogenation Activity
3.5.2. Catalase Activity
3.5.3. Peroxidase Activity
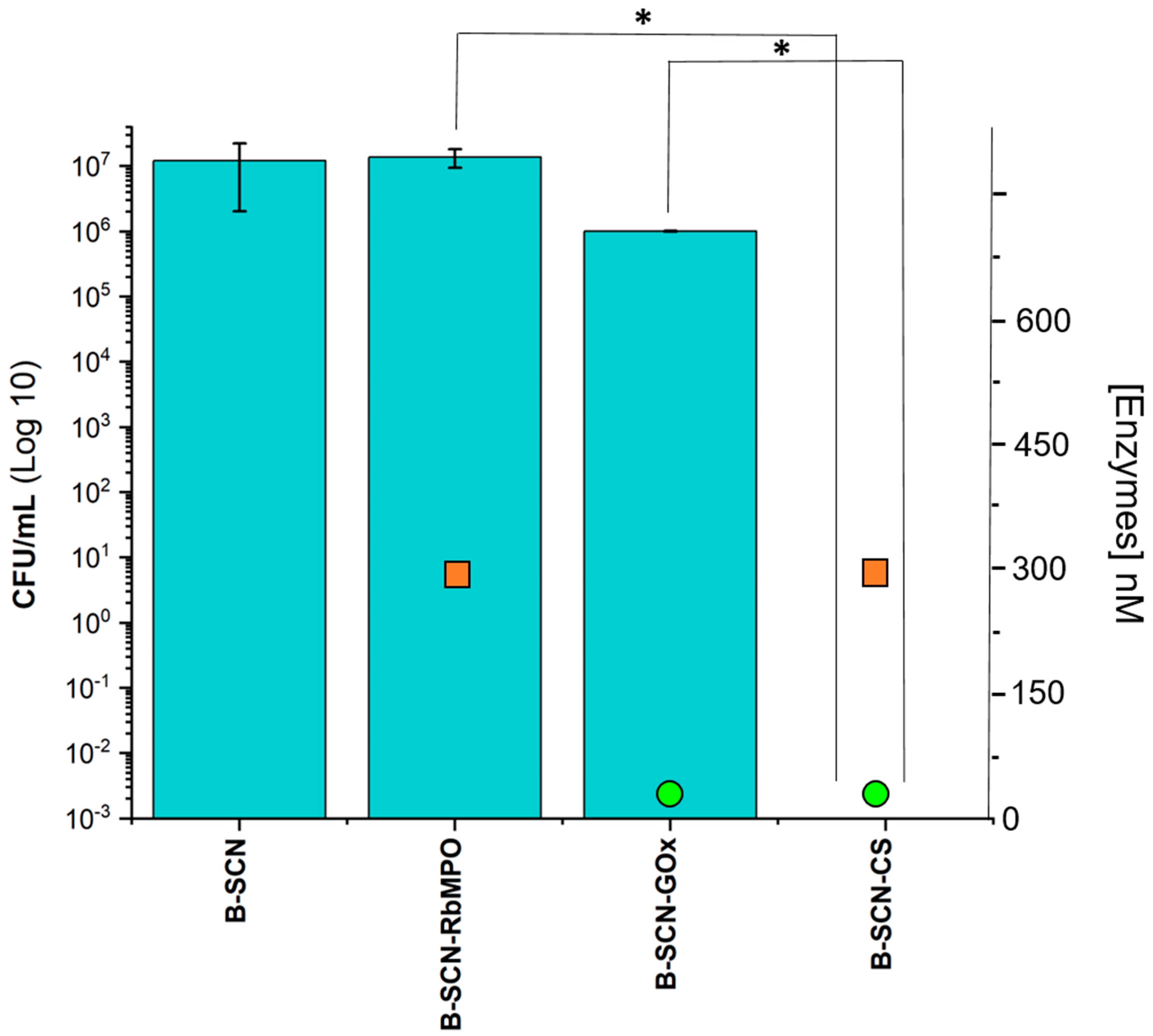
3.6. Microbicidal Activity
3.7. Activity in Function of Temperature, pH, and Storage Conditions
4. Discussion
5. Conclusions
6. Patent
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABAH | 4-aminobenzoic acid hydrazide |
| ABTS | 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid |
| APF | Aminophenylfluorescein |
| EPO | Eosinophil peroxidase |
| hMPO | human myeloperoxidase |
| IPTG | Isopropyl β-D-1-thiogalactopyranoside |
| LPO | Lactoperoxidase |
| MMPs | Mammalian peroxidases |
| PDB | Protein Data Bank |
| RbMPO | Heme peroxidase from Rhodopirellula baltica |
| TPO | Thyroid peroxidase |
References
- Savelli, B.; Li, Q.; Webber, M.; Jemmat, A.; Robitaille, A.; Zamocky, M.; Mathé, C.; Dunand, C. RedoxiBase: A database for ROS homeostasis regulated proteins. Redox Biol. 2019, 26, 101247. [Google Scholar] [CrossRef] [PubMed]
- Auer, M.; Nicolussi, A.; Schütz, G.; Furtmüller, P.G.; Obinger, C. How Covalent Heme to Protein Bonds Influence the Formation and Reactivity of Redox Intermediates of a Bacterial Peroxidase. J. Biol. Chem. 2014, 289, 31480–31491. [Google Scholar] [CrossRef] [PubMed]
- Oxvig, C.; Thomsen, A.R.; Overgaard, M.T.; Sørensen, E.S.; Højrup, P.; Bjerrum, M.J.; Gleich, G.J.; Sottrup-Jensen, L. Biochemical Evidence for Heme Linkage through Esters with Asp-93 and Glu-241 in Human Eosinophil Peroxidase: The ester with Asp-93 is only partially formed in vivo. J. Biol. Chem. 1999, 274, 16953–16958. [Google Scholar] [CrossRef] [PubMed]
- Taurog, A. Molecular Evolution of Thyroid Peroxidase. Biochimie 1999, 81, 557–562. [Google Scholar] [CrossRef]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- Gan, Q.; Chi, H.; Dalmo, R.A.; Meng, X.; Tang, X.; Xing, J.; Sheng, X.; Zhan, W. Characterization of myeloperoxidase and its contribution to antimicrobial effect on extracellular traps in flounder (Paralichthys olivaceus). Front. Immunol. 2023, 14, 1124813. [Google Scholar] [CrossRef]
- Weiss, G.; Schaible, U.E. Macrophage Defense Mechanisms against Intracellular Bacteria. Immunol. Rev. 2015, 264, 182–203. [Google Scholar] [CrossRef]
- Schürmann, N.; Forrer, P.; Casse, O.; Li, J.; Felmy, B.; Burgener, A.-V.; Ehrenfeuchter, N.; Hardt, W.-D.; Recher, M.; Hess, C.; et al. Myeloperoxidase Targets Oxidative Host Attacks to Salmonella and Prevents Collateral Tissue Damage. Nat. Microbiol. 2017, 2, 1–9. [Google Scholar] [CrossRef]
- Krivošíková, K.; Šupčíková, N.; Gaál Kovalčíková, A.; Janko, J.; Pastorek, M.; Celec, P.; Podracká, Ľ.; Tóthová, Ľ. Neutrophil extracellular traps in urinary tract infection. Front. Pediatr. 2023, 11, 1154139. [Google Scholar] [CrossRef]
- Davies, M.J.; Hawkins, C.L.; Pattison, D.I.; Rees, M.D. Mammalian Heme Peroxidases: From Molecular Mechanisms to Health Implications. Antioxid. Redox Signal. 2008, 10, 1199–1234. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, A.K.; Kaushik, S.; Sinha, M.; Singh, R.P.; Sharma, P.; Sirohi, H.; Kaur, P.; Singh, T.P. Lactoperoxidase: Structural Insights into the Function, Ligand Binding and Inhibition. Int. J. Biochem. Mol. Biol. 2013, 4, 108–128. [Google Scholar]
- Ruf, J.; Carayon, P. Structural and Functional Aspects of Thyroid Peroxidase. Arch. Biochem. Biophys. 2006, 445, 269–277. [Google Scholar] [CrossRef]
- Kettle, A.J.; Winterbourn, C.C. Chapter 12: Myeloperoxidase: Structure and Function of the Green Heme Peroxidase of Neutrophils; RCS Metallobiology. In Heme Peroxidases; 2015; pp. 272–308. [Google Scholar] [CrossRef]
- Fiedler, T.J.; Davey, C.A.; Fenna, R.E. X-ray Crystal Structure and Characterization of Halide-Binding Sites of Human Myeloperoxidase at 1.8 Å Resolution. J. Biol. Chem. 2000, 275, 11964–11971. [Google Scholar] [CrossRef]
- Nauseef, W.M. Biosynthesis of Human Myeloperoxidase. Arch. Biochem. Biophys. 2018, 642, 1–9. [Google Scholar] [CrossRef]
- Singh, P.K.; Iqbal, N.; Sirohi, H.V.; Bairagya, H.R.; Kaur, P.; Sharma, S.; Singh, T.P. Structural basis of activation of mammalian heme peroxidases. Prog. Biophys. Mol. Biol. 2018, 133, 49–55. [Google Scholar] [CrossRef]
- Furtmüller, P.G.; Burner, U.; Regelsberger, G.; Obinger, C. Spectral and Kinetic Studies on the Formation of Eosinophil Peroxidase Compound I and Its Reaction with Halides and Thiocyanate. Biochemistry 2000, 39, 15578–15584. [Google Scholar] [CrossRef]
- Furtmüller, P.G.; Jantschko, W.; Regelsberger, G.; Jakopitsch, C.; Arnhold, J.; Obinger, C. Reaction of Lactoperoxidase Compound I with Halides and Thiocyanate. Biochemistry 2002, 41, 11895–11900. [Google Scholar] [CrossRef]
- Arnhold, J.; Malle, E. Halogenation Activity of Mammalian Heme Peroxidases. Antioxidants 2022, 11, 890. [Google Scholar] [CrossRef]
- Singh, P.K.; Ahmad, N.; Yamini, S.; Singh, R.P.; Singh, A.K.; Sharma, P.; Smith, M.L.; Sharma, S.; Singh, T.P. Structural evidence of the oxidation of iodide ion into hyper-reactive hypoiodite ion by mammalian heme lactoperoxidase. Protein Sci. 2022, 31, 384–395. [Google Scholar] [CrossRef]
- Iwamoto, H.; Kobayashi, T.; Hasegawa, E.; Morita, Y. Reaction of Human Myeloperoxidase with Hydrogen Peroxide and Its True Catalase Activity. J. Biochem. 1987, 101, 1407–1412. [Google Scholar] [CrossRef]
- Auer, M.; Gruber, C.; Bellei, M.; Pirker, K.F.; Zamocky, M.; Kroiss, D.; Teufer, S.A.; Hofbauer, S.; Soudi, M.; Battistuzzi, G.; et al. A Stable Bacterial Peroxidase with Novel Halogenating Activity and an Autocatalytically Linked Heme Prosthetic Group. J. Biol. Chem. 2013, 288, 27181–27199. [Google Scholar] [CrossRef] [PubMed]
- Ahariz, M.; Courtois, P. Candida Albicans Susceptibility to Lactoperoxidase-Generated Hypoiodite. Clin. Cosmet. Investig. Dent. 2010, 2, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Cegolon, L. Investigating hypothiocyanite against SARS-CoV-2. Int. J. Hyg. Environ. Health 2020, 227, 113520. [Google Scholar] [CrossRef] [PubMed]
- Cegolon, L.; Salata, C.; Piccoli, E.; Juarez, V.; Palu’, G.; Mastrangelo, G.; Calistri, A. In Vitro Antiviral Activity of Hypothiocyanite against A/H1N1/2009 Pandemic Influenza Virus. Int. J. Hyg. Environ. Health 2014, 217, 17–22. [Google Scholar] [CrossRef]
- Patel, U.; Gingerich, A.; Widman, L.; Sarr, D.; Tripp, R.A.; Rada, B. Susceptibility of Influenza Viruses to Hypothiocyanite and Hypoiodite Produced by Lactoperoxidase in a Cell-Free System. PLoS ONE 2018, 13, e0199167. [Google Scholar] [CrossRef]
- Chochola, J.; Yamaguchi, Y.; Moguilevsky, N.; Bollen, A.; Strosberg, A.D.; Stanislawski, M. Virucidal Effect of Myeloperoxidase on Human Immunodeficiency Virus Type 1-Infected T Cells. Antimicrob. Agents Chemother. 1994, 38, 969–972. [Google Scholar] [CrossRef]
- El Messaoudi, K.; Verheyden, A.-M.; Thiry, L.; Fourez, S.; Tasiaux, N.; Bollen, A.; Moguilevsky, N. Human Recombinant Myeloperoxidase Antiviral Activity on Cytomegalovirus. J. Med. Virol. 2002, 66, 218–223. [Google Scholar] [CrossRef]
- Denys, G.A.; Devoe, N.C.; Gudis, P.; May, M.; Allen, R.C.; Stephens, J.T. Mechanism of Microbicidal Action of E-101 Solution, a Myeloperoxidase-Mediated Antimicrobial, and Its Oxidative Products. Infect. Immun. 2019, 87, e00261-19. [Google Scholar] [CrossRef]
- Musser, R.C. Enhanced Lactoperoxidase System for Treatment of Milk Products. WO2011116052A3. 11 March 2011. Available online: https://patents.google.com/patent/US20110229598A1/en (accessed on 3 April 2023).
- Shin, K.; Horigome, A.; Yamauchi, K. Oral Disinfectant, Food Additive Comprising the Disinfectant. WO2008105113A1. 28 February 2007. Available online: https://patents.google.com/patent/EP2127667A1/en (accessed on 3 April 2023).
- Tenovuo, J.; Pruitt, K.M.; Mansson-Rahemtulla, B.; Harrington, P.; Baldone, D.C. Products of Thiocyanate Peroxidation: Properties and Reaction Mechanisms. Biochim. Et Biophys. Acta (BBA)—Protein Struct. Mol. Enzymol. 1986, 870, 377–384. [Google Scholar] [CrossRef]
- Su, Q.; Klinman, J.P. Nature of Oxygen Activation in Glucose Oxidase from Aspergillus Niger: The Importance of Electrostatic Stabilization in Superoxide Formation. Biochemistry 1999, 38, 8572–8581. [Google Scholar] [CrossRef]
- Kettle, A.J.; Gedye, C.A.; Winterbourn, C.C. Mechanism of Inactivation of Myeloperoxidase by 4-Aminobenzoic Acid Hydrazide. Biochem. J. 1997, 321 Pt 2, 503–508. [Google Scholar] [CrossRef]
- Furtmüller, P.G.; Obinger, C.; Hsuanyu, Y.; Dunford, H.B. Mechanism of reaction of myeloperoxidase with hydrogen peroxide and chloride ion. Eur. J. Biochem. 2000, 267, 5858–5864. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate 1 | Substrate 2 | Reagent | Detection | Wavelength nm | ε M−1·cm−1 | |
|---|---|---|---|---|---|---|
| Transition RbMPO-Fe3+ to Compound I or Compound I to RbMPO-Fe3+ | H2O2 | X | X | RbMPO-Fe3+ | 412 | 55,390 |
| Catalase activity | H2O2 | X | X | H2O2 | 240 | 43.6 |
| Peroxidase activity | H2O2 | ABTS | X | ABTS.+ | 320 | 36,000 |
| Halogenase activity | H2O2 | NaCl | Taurine | Taurinechlorinated | 253 | 429 |
| H2O2 | NaBr | Taurine | TaurineBrominated | 289 | 415 | |
| H2O2 | NaSCN | X | SCNO− formation | 240 | 951 |
| Catalase Activity | Halogenase Activity | Peroxidase Activity | |||
|---|---|---|---|---|---|
| Substrate | H2O2 | SCN− | Br− | Cl− | ABTS |
| KM (mM) | 0.17 ± 0.11 | 0.51 ± 0.08 | 133 ± 32 | / | |
| kcat (s−1) | 56.6 ± 20.6 | 56.9 ± 6.9 | 76.3 ± 5.12 | / | |
| kcat/KM (M−1·s−1) | 3.33 × 105 ±0.21 × 105 | 1.12 × 105 ±0.31 × 105 | 576 ± 172 | 5.46 ± 0.67 * | 1.06 × 104 ±0.15 × 104 * |
| Ki (mM) | 1.07 ± 0.7 | - | - | / | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Céré, C.; Delord, B.; Kenfack Ymbe, P.; Vimbert, L.; Chapel, J.-P.; Stines-Chaumeil, C. A Bacterial Myeloperoxidase with Antimicrobial Properties. BioTech 2023, 12, 33. https://doi.org/10.3390/biotech12020033
Céré C, Delord B, Kenfack Ymbe P, Vimbert L, Chapel J-P, Stines-Chaumeil C. A Bacterial Myeloperoxidase with Antimicrobial Properties. BioTech. 2023; 12(2):33. https://doi.org/10.3390/biotech12020033
Chicago/Turabian StyleCéré, Claire, Brigitte Delord, Parfait Kenfack Ymbe, Léa Vimbert, Jean-Paul Chapel, and Claire Stines-Chaumeil. 2023. "A Bacterial Myeloperoxidase with Antimicrobial Properties" BioTech 12, no. 2: 33. https://doi.org/10.3390/biotech12020033