
Post-Harvest Prevention of Fusariotoxin Contamination of Agricultural Products by Irreversible Microbial Biotransformation: Current Status and Prospects

Abstract
:1. Introduction
2. Biotransformation of Zearalenone
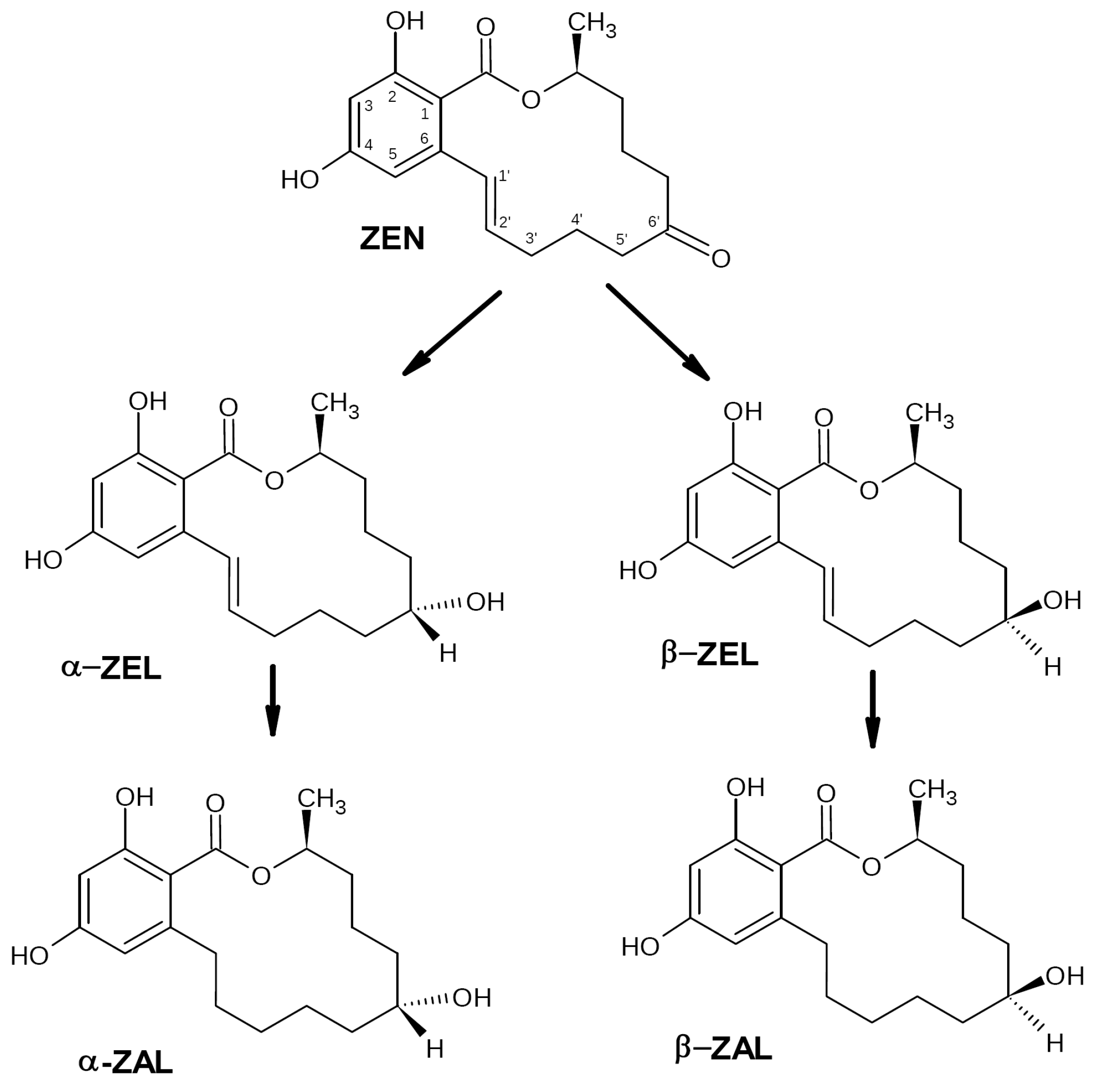
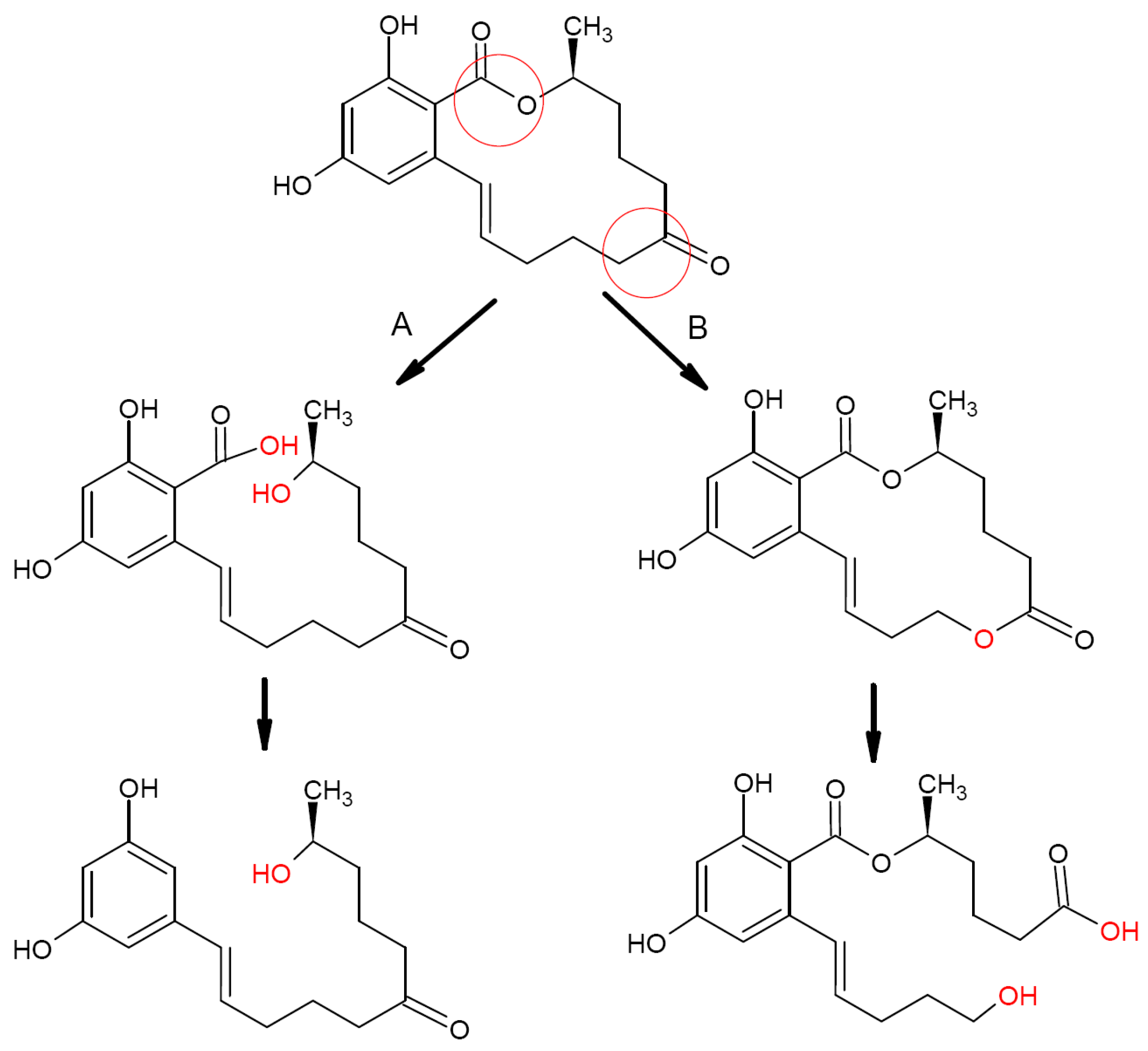
2.1. Zearalenone Biotransformation Pathways
2.2. Microbial Transformation of Zearalenone into Nontoxic Metabolites
2.2.1. ZEN-Transforming Bacteria
2.2.2. ZEN-Transforming Fungi and Yeasts
2.2.3. ZEN-Transforming Microbial Enzymes
3. Biotransformation of Deoxynivalenol
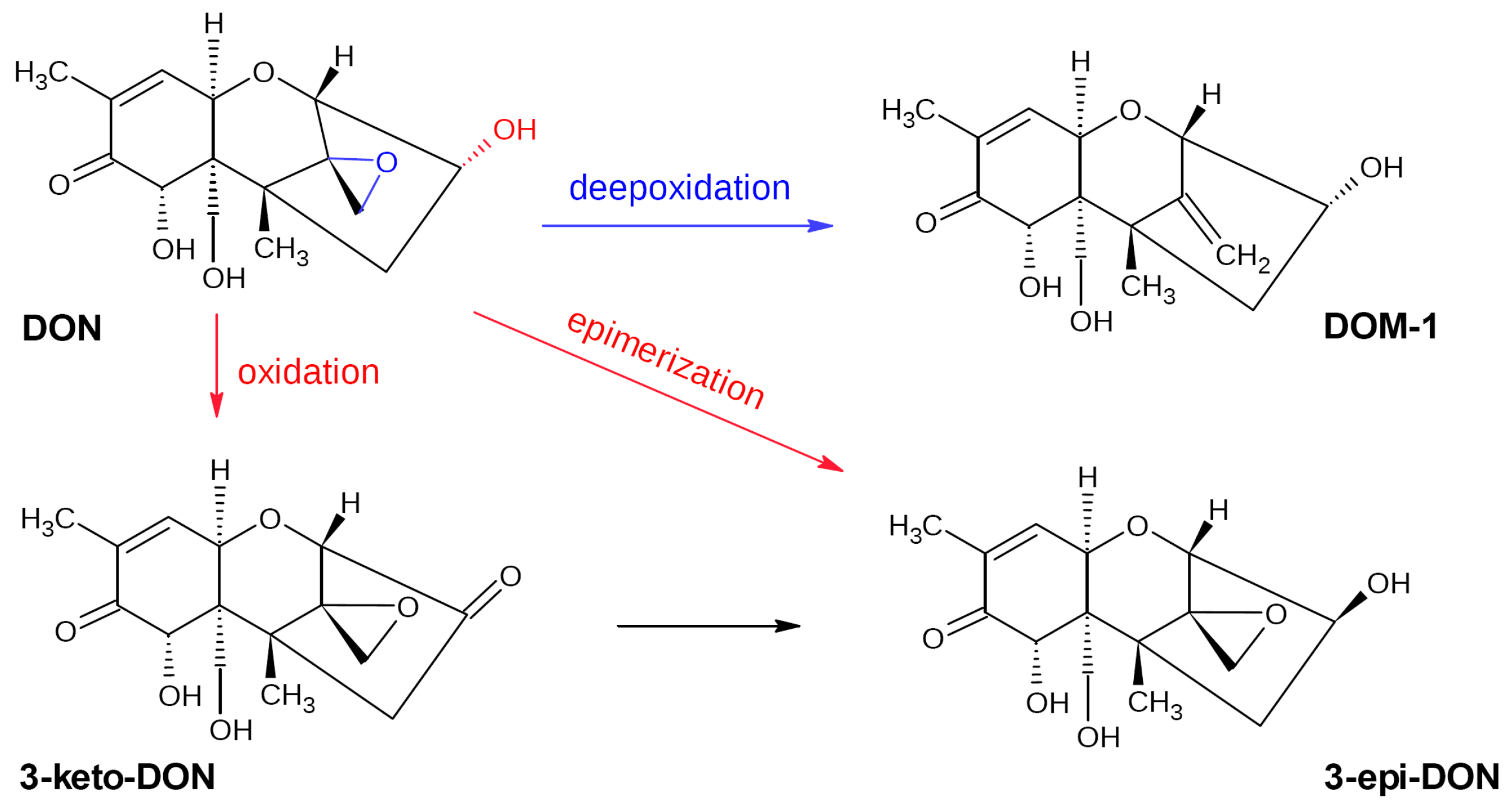
3.1. Deoxynivalenol Biotransformation Pathways
3.2. DON-Transforming Microorganisms
3.2.1. Anaerobic Bacteria
3.2.2. Aerobic Bacteria
3.2.3. DON-Transforming Enzymes
4. Biotransformation of Fumonisin B1
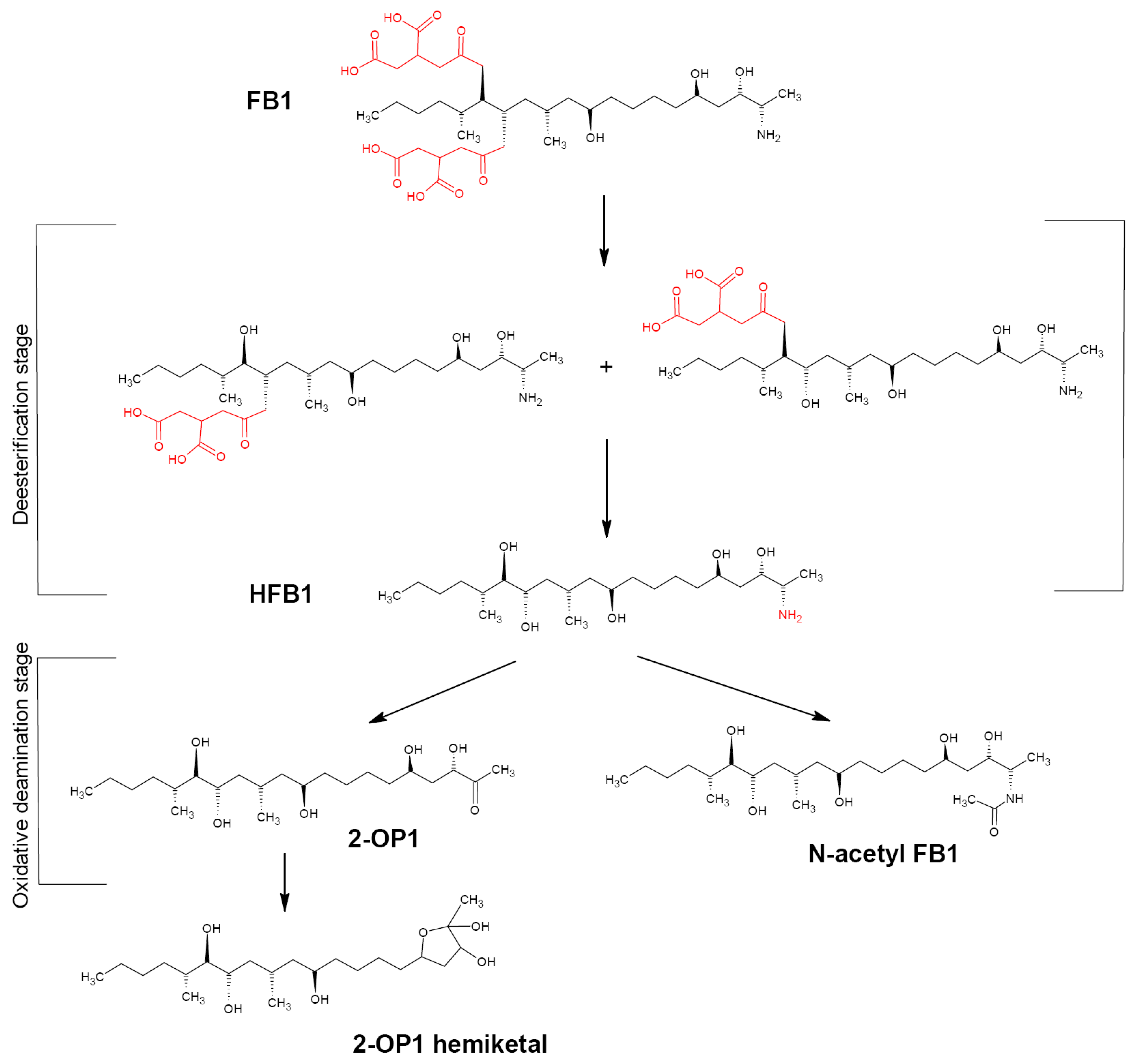
4.1. Fumonisin B1 Biotransformation Pathway
4.2. FB1-Transforming Microorganisms
4.2.1. FB1-Transforming Bacteria
4.2.2. FB1-Transforming Fungi and Yeasts
4.2.3. FB1-Transforming Enzymes
5. Prospects for Use of Microbial Destructors of Fusariotoxins and Promising Directions of Future Studies
5.1. Promising Directions of Studies Intended to Search for Potential Microbial and Enzymatic Agents Able to Transform Fusariotoxins
5.2. Future Directions of Studies to Solve Issues Related to the Practical Application of Fusariotoxin-Transforming Bioagents
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Wachowska, U.; Packa, D.; Wiwart, M. Microbial inhibition of Fusarium pathogens and biological modification of trichothecenes in cereal grains. Toxins 2017, 9, 408. [Google Scholar] [CrossRef]
- Mielniczuk, E.; Skwaryło-Bednarz, B. Fusarium head blight, mycotoxins and strategies for their reduction. Agronomy 2020, 10, 509. [Google Scholar] [CrossRef]
- BIOMIN Mycotoxin Survey Q3 2021 Results. Available online: https://www.biomin.net/science-hub/biomin-mycotoxin-survey-q3-2021-results/ (accessed on 2 April 2023).
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed—Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed]
- Ropejko, K.; Twarużek, M. Zearalenone and its metabolites—General overview, occurrence, and toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef]
- Minervini, F.; Dell’Aquila, M.E. Zearalenone and reproductive function in farm animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef]
- Ismaiel, A.A.; Papenbrock, J. Mycotoxins: Producing fungi and mechanisms of phytotoxicity. Agriculture 2015, 5, 492–537. [Google Scholar] [CrossRef]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in cereal grains—An update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.C.S.M.; De Pierri, C.R.; Evangelista, A.G.; Gomes, A.S.d.L.P.B.; Luciano, F.B. Deoxynivalenol: Toxicology, degradation by bacteria, and phylogenetic analysis. Toxins 2022, 14, 90. [Google Scholar] [CrossRef]
- Aydinoglu, F. Fumonisin mycotoxins: Metabolism, toxicity, detection and prevention. In Mathematics and Natural Sciences; Karapinar, R., Ed.; Livre de Lyon: Lyon, France, 2021; pp. 1–21. [Google Scholar]
- Rheeder, J.P.; Marasas, W.F.; Vismer, H.F. Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef]
- Qu, L.; Wang, L.; Ji, H.; Fang, Y.; Lei, P.; Zhang, X.; Jin, L.; Sun, D.; Dong, H. Toxic mechanism and biological detoxification of fumonisins. Toxins 2022, 14, 182. [Google Scholar] [CrossRef]
- Qin, X.; Zhang, R.X.; Ge, S.; Zhou, T.; Liang, Y.K. Sphingosine kinase AtSPHK1 functions in fumonisin B1-triggered cell death in Arabidopsis. Plant Physiol. Biochem. 2017, 119, 70–80. [Google Scholar] [CrossRef]
- Bojja, R.S.; Cerny, R.L.; Proctor, R.H.; Du, L. Determining the biosynthetic sequence in the early steps of the fumonisin pathway by use of three gene-disruption mutants of Fusarium verticillioides. J. Agric. Food Chem. 2004, 52, 2855–2860. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Song, G.; Lim, W. Effects of mycotoxin-contaminated feed on farm animals. J. Hazard. Mater. 2020, 389, 122087. [Google Scholar] [CrossRef]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public health impacts of foodborne mycotoxins. Ann. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Kamle, M.; Mahato, D.K.; Devi, S.; Lee, K.E.; Kang, S.G.; Kumar, P. Fumonisins: Impact on agriculture, food, and human health and their management strategies. Toxins 2019, 11, 328. [Google Scholar] [CrossRef]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.K.; Bellaloui, N.; Bruns, H.A. Investigating transgenic corn hybrids as a method for mycotoxin control. J. Food Nutr. Sci. 2016, 7, 44–54. [Google Scholar] [CrossRef]
- De la Campa, R.; Hooker, D.C.; Miller, J.D.; Schaafsma, A.W.; Hammond, B.G. Modeling effects of environmental, insect damage, and Bt genotypes on fumonisin accumulation in maize in Argentina and the Philippines. Mycopathologia 2005, 159, 539–552. [Google Scholar] [CrossRef]
- Hope, R.; Aldred, D.; Magan, N. Comparison of environmental profiles for growth and deoxynivalenol production by Fusarium culmorum and F. graminearum on wheat grain. Lett. Appl. Microbiol. 2005, 40, 295–300. [Google Scholar] [CrossRef]
- Alberts, J.F.; Lilly, M.; Rheeder, J.P.; Burger, H.-M.; Shephard, G.S.; Gelderblom, W.C.A. Technological and community-based methods to reduce mycotoxin exposure. Food Control 2017, 73 Pt A, 101–109. [Google Scholar] [CrossRef]
- Powell, A.J.; Vujanovic, V. Evolution of Fusarium head blight management in wheat: Scientific perspectives on biological control agents and crop genotypes protocooperation. Appl. Sci. 2021, 11, 8960. [Google Scholar] [CrossRef]
- Grenier, B.; Bracarense, A.P.; Leslie, J.F.; Oswald, I.P. Physical and chemical methods for mycotoxin decontamination in maize. In Mycotoxin Reduction in Grain Chains; Leslie, J.F., Logrieco, A.F., Eds.; Wiley Blackwell: New Delhi, India, 2014; pp. 116–129. [Google Scholar] [CrossRef]
- Hu, D.; Wu, A. Chemical and physical treatments for reducing mycotoxin contaminations. In Food Safety & Mycotoxins; Wu, A., Ed.; Springer: Singapore, 2019; pp. 145–165. [Google Scholar] [CrossRef]
- Ji, C.; Fan, Y.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, I.; Audenaert, K.; De Gelder, L. Biodegradation of mycotoxins: Tales from known and unexplored worlds. Front. Microbiol. 2016, 7, 561. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.; Zhou, T. Strategies and methodologies for developing microbial detoxification systems to mitigate mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wu, W.; Pan, J.; Long, M. Detoxification strategies for zearalenone using microorganisms: A review. Microorganisms 2019, 7, 208. [Google Scholar] [CrossRef] [PubMed]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Bohm, J.; Zentek, J. Decontamination and detoxification strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the effectiveness of microbial biodegradation. Food Addit. Contam. Part A 2010, 27, 510–520. [Google Scholar] [CrossRef]
- Azam, M.S.; Yu, D.; Wu, A. Enzymes for degradation of Fusarium mycotoxins. In Food Safety & Mycotoxins; Wu, A., Ed.; Springer: Singapore, 2019; pp. 113–135. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Songsermsakul, P.; Sontag, G.; Cichnamarkl, M.; Zentek, J.; Razzazifazeli, E. Determination of zearalenone and its metabolites in urine, plasma and faeces of horses by HPLC–APCI–MS. J. Chromatogr. B 2006, 843, 252–261. [Google Scholar] [CrossRef]
- Richardson, K.E.; Hagler, W.M., Jr.; Hamilton, P.B. Bioconversion of alpha-[14C] zearalenol and beta-[14C]zearalenol into [14C]zearalenone by Fusarium roseum ‘Gibbosum’. Appl. Environ. Microbiol. 1984, 47, 1206–1209. [Google Scholar] [CrossRef] [PubMed]
- Böswald, C.; Engelhardt, G.; Vogel, H.; Wallnöfer, P.R. Metabolism of the Fusarium mycotoxins zearalenone and deoxynivalenol by yeast strains of technological relevance. Nat. Toxins 1995, 3, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, K.H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of aflatoxin, ochratoxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Drouin, P.; Lepp, D.; Li, X.-Z.; Zhu, H.; Castex, M.; Zhou, T. A novel microbial zearalenone transformation through phosphorylation. Toxins 2021, 13, 294. [Google Scholar] [CrossRef] [PubMed]
- Binder, S.B.; Schwartz-Zimmermann, H.E.; Varga, E.; Bichl, G.; Michlmayr, H.; Adam, G.; Berthiller, F. Metabolism of zearalenone and its major modified forms in pigs. Toxins 2017, 9, 56. [Google Scholar] [CrossRef]
- Catteuw, A.; Broekaert, N.; De Baere, S.; Lauwers, M.; Gasthuys, E.; Huybrechts, B.; Callebaut, A.; Ivanova, L.; Uhlig, S.; De Boevre, M.; et al. Insights into in vivo absolute oral bioavailability, biotransformation, and toxicokinetics of zearalenone, α-zearalenol, β-zearalenol, zearalenone-14-glucoside, and zearalenone-14-sulfate in pigs. J. Agric. Food Chem. 2019, 67, 3448–3458. [Google Scholar] [CrossRef]
- Dellafiora, L.; Galaverna, G.; Righi, F.; Cozzini, P.; Dall’Asta, C. Assessing the hydrolytic fate of the masked mycotoxin zearalenone-14-glucoside—A warning light for the need to look at the “maskedome”. Food Chem. Toxicol. 2017, 99, 9–16. [Google Scholar] [CrossRef]
- Shikhaliyeva, I.; Teker, T.; Albayrak, G. Masked mycotoxins of deoxynivalenol and zearalenone—Unpredicted toxicity. Biomed. J. Sci. Tech. Res. 2020, 29, 22288–22293. [Google Scholar] [CrossRef]
- Tinyiro, S.E.; Wokadala, C.; Xu, D.; Yao, W. Adsorption and degradation of zearalenone by bacillus strains. Folia Microbiol. 2011, 56, 321–327. [Google Scholar] [CrossRef]
- Wang, G.; Yu, M.; Dong, F.; Shi, J.; Xu, J. Esterase activity inspired selection and characterization of zearalenone degrading bacteria Bacillus pumilus ES-21. Food Control 2017, 77, 57–64. [Google Scholar] [CrossRef]
- Guo, Y.; Zhou, J.; Tang, Y.; Ma, Q.; Zhang, J.; Ji, C.; Zhao, L. Characterization and genome analysis of a zearalenone-degrading Bacillus velezensis strain ANSB01E. Curr. Microbiol. 2020, 77, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wu, H.; Tang, Y.; Qiu, L. Cloning, expression of a peroxiredoxin gene from Acinetobacter sp. SM04 and characterization of its recombinant protein for zearalenone detoxification. Microbiol. Res. 2012, 167, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Ando, N.; Kimura, M.; Kakeya, H.; Osada, H.; Yamaguchi, I. A novel lactonohydrolase responsible for the detoxification of zearalenone: Enzyme purification and gene cloning. Biochem. J. 2002, 365, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cserháti, M.; Kriszt, B.; Krifaton, C.; Szoboszlay, S.; Hahn, J.; Toth, S.; Nagy, I.; Kukolya, J. Mycotoxin-degradation profile of Rhodococcus strains. Int. J. Food Microbiol. 2013, 166, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Krifaton, C.; Kriszt, B.; Risa, A.; Szoboszlay, S.; Cserháti, M.; Harkai, P.; Eldrige, M.; Wang, J.; Kukolya, J. Application of a yeast estrogen reporter system for screening zearalenone degrading microbes. J. Hazard. Mater. 2012, 244–245, 429–435. [Google Scholar] [CrossRef]
- Kriszt, R.; Krifaton, C.; Szoboszlay, S.; Cserháti, M.; Kriszt, B.; Kukolya, J.; Czeh, A.; Fehér-Tóth, S.; Török, L.; Szőke, Z.; et al. A new zearalenone biodegradation strategy using non-pathogenic Rhodococcus pyridinivorans K408 strain. PLoS ONE 2012, 7, e43608. [Google Scholar] [CrossRef]
- Risa, A.; Krifaton, C.; Kukolya, J.; Kriszt, B.; Cserháti, M.; Táncsics, A. Aflatoxin B1 and zearalenone-detoxifying profile of Rhodococcus type strains. Curr. Microbiol. 2018, 75, 907–917. [Google Scholar] [CrossRef]
- Fruhauf, S.; Thamhesl, M.; Fajtl, P.; Klingenbrunner, V.; Kunz-Vekiru, E.; Adam, G.; Schatzmayr, G.; Moll, W. Enzymatic hydrolysis of zearalenone by ZenA of Rhodococcus erythropolis PFA D8-1. In Proceedings of the Abstracts of the 37th Mycotoxin Workshop, Bratislava, Slovakia, 1–3 June 2015; Slovak University of Technology: Bratislava, Slovakia, 2015; p. 151. [Google Scholar]
- Yu, Y.; Qiu, L.; Wu, H.; Tang, Y.; Yu, Y.; Li, X.; Liu, D. Degradation of zearalenone by the extracellular extracts of Acinetobacter sp. SM04 liquid cultures. Biodegradation 2011, 22, 613–622. [Google Scholar] [CrossRef]
- Altalhi, A.D. Plasmid-mediated detoxification of mycotoxin zearalenone in Pseudomonas ZEA-1. Am. J. Biotechnol. Biochem. 2007, 3, 150–158. [Google Scholar] [CrossRef]
- El-Sharkawy, S.; Abul-Hajj, Y.J. Microbial cleavage of zearalenone. Xenobiotica 1988, 18, 365–371. [Google Scholar] [CrossRef]
- Kakeya, H.; Takahashi-Ando, N.; Kimura, M.; Onose, R.; Yamaguchi, I.; Osada, H. Biotransformation of the mycotoxin, zearalenone, to a non-estrogenic compound by a fungal strain of Clonostachys sp. Biosci. Biotechnol. Biochem. 2002, 66, 2723–2726. [Google Scholar] [CrossRef] [PubMed]
- Shcherbakova, L.A.; Mikityuk, O.D.; Nazarova, T.A.; Dzhavakhiya, V.G. Study of aflatoxin B1-destroying activity of Gliocladium roseum and Trichoderma viride and their antagonism toward toxigenic Aspergillus flavus. Agric. Biol. 2016, 51, 946–950. [Google Scholar] [CrossRef]
- Mikityuk, O.D.; Statsyuk, N.V.; Nazarova, T.A.; Shcherbakova, L.A.; Dzhavakhiya, V.G. Mycotoxin degradation by microbial metabolites. In Proceedings of the International Scientific Conference PLAMIC 2020 “Plants and Microorganisms: Biotechnology of the Future”, Saratov, Russia, 5–9 October 2020; Tikhonovich, I.A., Ed.; “Agrarnoe Obrazovanie i Nauka” Association: Saratov, Russia, 2020; p. 234. [Google Scholar]
- Sun, X.; He, X.; Xue, K.; Li, Y.; Xu, D.; Qian, H. Biological detoxification of zearalenone by Aspergillus niger strain FS10. Food Chem. Toxicol. 2014, 72, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Yu, J.; Yang, Y.; Yuan, X.; Yang, J.; Zhang, Y.; Sun, J.; Sun, X. Exploration on the enhancement of detoxification ability of zearalenone and its degradation products of Aspergillus niger FS10 under directional stress of zearalenone. Toxins 2021, 13, 720. [Google Scholar] [CrossRef] [PubMed]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarc’h, A.; André, F.; Delaforge, M.; Lebrihi, A. Transformation of zearalenone to zearalenone-sulfate by Aspergillus spp. World Mycotoxin J. 2010, 3, 183–191. [Google Scholar] [CrossRef]
- Gamage, N.; Barnett, A.; Hempel, N.; Duggleby, R.G.; Windmill, K.F.; Martin, J.L.; McManus, M.E. Human sulfotransferases and their role in chemical metabolism. Toxicol. Sci. 2006, 90, 5–22. [Google Scholar] [CrossRef]
- Plasencia, J.; Mirocha, C.J. Isolation and characterization of zearalenone sulfate produced by Fusarium spp. Appl. Environ. Microbiol. 1991, 57, 146–150. [Google Scholar] [CrossRef]
- Vekiru, E.; Hametner, C.; Mitterbauer, R.; Rechthaler, J.; Adam, G.; Schatzmayr, G.; Krska, R.; Schuhmacher, R. Cleavage of zearalenone by Trichosporon mycotoxinivorans to a novel nonestrogenic metabolite. Appl. Environ. Microbiol. 2010, 76, 2353–2359. [Google Scholar] [CrossRef]
- Sun, J.; Xia, Y.; Ming, D. Whole-genome sequencing and bioinformatics analysis of Apiotrichum mycotoxinivorans: Predicting putative zearalenone-degradation enzymes. Front. Microbiol. 2020, 11, 1866. [Google Scholar] [CrossRef]
- Takahashi-Ando, N.; Ohsato, S.; Shibata, T.; Hamamoto, H.; Yamaguchi, I.; Kimura, M. Metabolism of zearalenone by genetically modified organisms expressing the detoxification gene from Clonostachys rosea. Appl. Environ. Microbiol. 2004, 70, 3239–3245. [Google Scholar] [CrossRef]
- Xiang, L.; Wang, Q.; Zhou, Y.; Yin, L.; Zhang, G.; Ma, Y. High-level expression of a ZEN-detoxifying gene by codon optimization and biobrick in Pichia pastoris. Microbiol. Res. 2016, 193, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; Hsu, T.C.; Cheng, K.C.; Liu, J.R. Expression of the Clonostachys rosea lactonohydrolase gene by Lactobacillus reuteri to increase its zearalenone-removing ability. Microb. Cell Factories 2017, 16, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, W.; Wu, H.; Zhang, W.; Mu, W. Identification of a potent enzyme for the detoxification of zearalenone. J. Agric. Food Chem. 2020, 68, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Chang, X.; Wang, N. Cloning of ZEN-degrading enzyme zhd795 and study on degradation activity. J. Food Sci. Technol. 2018, 36, 28–32. [Google Scholar] [CrossRef]
- Cheng, B.; Shi, W.; Luo, J.; Peng, F.; Wan, C.; Wei, H. Cloning of zearalenone-degraded enzyme gene (zen-jjm) and its expression and activity analysis. J. Agric. Biotechnol. 2010, 18, 225–230. [Google Scholar]
- Yu, X.; Tu, T.; Luo, H.; Huang, H.; Su, X.; Wang, Y.; Wang, Y.; Zhang, J.; Bai, Y.; Yao, B. Biochemical characterization and mutational analysis of a lactone hydrolase from Phialophora americana. J. Agric. Food Chem. 2020, 68, 2570–2577. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Zhang, Y.; Zhang, X.; Huang, H. Cloning and characterization of three novel enzymes responsible for the detoxification of zearalenone. Toxins 2022, 14, 82. [Google Scholar] [CrossRef]
- Chang, X.; Liu, H.; Sun, J.; Wang, J.; Zhao, C.; Zhang, W.; Zhang, J.; Sun, C. Zearalenone removal from corn oil by an enzymatic strategy. Toxins 2020, 12, 117. [Google Scholar] [CrossRef]
- Shcherbakova, L.; Rozhkova, A.; Osipov, D.; Zorov, I.; Mikityuk, O.; Statsyuk, N.; Sinitsyna, O.; Dzhavakhiya, V.; Sinitsyn, A. Effective zearalenone degradation in model solutions and infected wheat grain using a novel heterologous lactonohydrolase secreted by recombinant Penicillium canescens. Toxins 2020, 12, 475. [Google Scholar] [CrossRef]
- Wang, M.; Yin, L.; Hu, H.; Selvaraj, J.N.; Zhou, Y.; Zhang, G. Expression, functional analysis and mutation of a novel neutral zearalenone-degrading enzyme. Int. J. Biol. Macromol. 2018, 118, 1284–1292. [Google Scholar] [CrossRef]
- Bi, K.; Zhang, W.; Xiao, Z.; Zhang, D. Characterization, expression and application of a zearalenone degrading enzyme from Neurospora crassa. AMB Express 2018, 8, 194. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Dorninger, C.; Faas, J.; Doupovec, B.; Aleschko, M.; Stoiber, C.; Höbartner-Gußl, A.; Schöndorfer, K.; Killinger, M.; Zebeli, Q.; Schatzmayr, D. Metabolism of zearalenone in the rumen of dairy cows with and without application of a zearalenone-degrading enzyme. Toxins 2021, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Dorninger, C.; Killinger, M.; Höbartner-Gußl, A.; Rosen, R.; Doupovec, B.; Aleschko, M.; Schwartz-Zimmermann, H.; Greitbauer, O.; Markovíc, Z.; Stankovíc, M.; et al. Enzymatic degradation of zearalenone in the gastrointestinal tract of pigs, chickens, and rainbow trout. Toxins 2023, 15, 48. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, C.; Yang, J.; Peng, X. A novel enzyme synthesized by Acinetobacter sp. SM04 is responsible for zearalenone biodegradation. Biosci. Biotechnol. Biochem. 2022, 86, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Xin, Y.; Su, X.; Wang, X.; Wang, Y.; Zhang, J.; Tu, T.; Yao, B.; Luo, H.; Huang, H. Efficient degradation of zearalenone by dye-decolorizing peroxidase from Streptomyces thermocarboxydus combining catalytic properties of manganese peroxidase and laccase. Toxins 2021, 13, 602. [Google Scholar] [CrossRef]
- Drzymala, S.S.; Binder, J.; Brodehl, A.; Penkert, M.; Rosowski, M.; Garbe, L.-A.; Koch, M. Estrogenicity of novel phase I and phase II metabolites of zearalenone and cis-zearalenone. Toxicon 2015, 105, 10–12. [Google Scholar] [CrossRef]
- Wang, X.; Qin, X.; Hao, Z.; Luo, H.; Yao, B.; Su, X. Degradation of four major mycotoxins by eight manganese peroxidases in presence of a dicarboxylic acid. Toxins 2019, 11, 566. [Google Scholar] [CrossRef]
- Garcia, S.O.; Feltrin, A.C.P.; Garda-Buffon, J. Zearalenone reduction by commercial peroxidase enzyme and peroxidases from soybean bran and rice bran. Food Addit. Contam. Part A 2018, 35, 1819–1831. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Y.; Liu, Y.; Ma, Q.; Ji, C.; Zhao, L. Detoxification of the mycoestrogen zearalenone by Bacillus licheniformis spore CotA laccase and application of immobilized laccase in contaminated corn meal. LWT 2022, 163, 113548. [Google Scholar] [CrossRef]
- Novozymes, A.S. Detoxification of Feed Products. World Patent 2009109607, 17 December 2009. [Google Scholar]
- Loi, M.; Fanelli, F.; Cimmarusti, M.T.; Mirabelli, V.; Haidukowski, M.; Logrieco, A.F.; Caliandro, R.; Mule, G. In Vitro single and combined mycotoxins degradation by Ery4 laccase from Pleurotus eryngii and redox mediators. Food Control 2018, 90, 401–406. [Google Scholar] [CrossRef]
- Song, Y.; Wang, Y.; Guo, Y.; Qiao, Y.; Ma, Q.; Ji, C.; Zhao, L. Degradation of zearalenone and aflatoxin B1 by Lac2 from Pleurotus pulmonarius in the presence of mediators. Toxicon 2021, 201, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Banu, I.; Lupu, A.; Aprodu, I. Degradation of zearalenone by laccase enzyme. Sci. Study Res. Chemistry Chem. Eng. Biotechnol. Food Ind. 2013, 14, 79. [Google Scholar]
- Xu, L.; Sun, X.; Wan, X.; Li, H.; Yan, F.; Han, R.; Li, H.; Li, Z.; Tian, Y.; Liu, X.; et al. Identification of a Bacillus amyloliquefaciens H6 thioesterase involved in zearalenone detoxification by transcriptomic analysis. J. Agric. Food. Chem. 2020, 68, 10071–10080. [Google Scholar] [CrossRef] [PubMed]
- Altalhi, A.D.; El-Deeb, B. Localization of zearalenone detoxification gene(s) in pZEA-1 plasmid of Pseudomonas putida ZEA-1 and expressed in Escherichia coli. J. Hazard. Mater. 2009, 161, 1166–1172. [Google Scholar] [CrossRef]
- Schwartz-Zimmermann, H.E.; Fruhmann, P.; Dänicke, S.; Wiesenberger, G.; Caha, S.; Weber, J.; Berthiller, F. Metabolism of deoxynivalenol and deepoxy-deoxynivalenol in broiler chickens, pullets, roosters and turkeys. Toxins 2015, 7, 4706–4729. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, G.S.; Pettersson, H.; Lundh, T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and deepoxy metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef]
- Kong, C.; Shin, S.Y.; Kim, B.G. Evaluation of mycotoxin sequestering agents for aflatoxin and deoxynivalenol: An in vitro approach. Springerplus 2014, 3, 1–4. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, D.; Cai, P.; Lin, H.; Ying, H.; Hu, Q.-N.; Wu, A. Elimination of Fusarium mycotoxin deoxynivalenol (DON) via microbial and enzymatic strategies: Current status and future perspectives. Trend. Food Sci. Technol. 2022, 124, 96–107. [Google Scholar] [CrossRef]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef]
- Bracarense, A.P.F.L.; Pierron, A.; Pinton, P.; Gerez, J.R.; Schatzmayr, G.; Moll, W.-D.; Zhou, T.; Oswald, I.P. Reduced toxicity of 3-epi-deoxynivalenol and de-epoxy-deoxynivalenol through deoxynivalenol bacterial biotransformation: In Vivo analysis in piglets. Food Chem. Toxicol. 2020, 140, 111241. [Google Scholar] [CrossRef]
- McCormick, S.P. Microbial detoxification of mycotoxins. J. Chem. Ecol. 2013, 39, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Khatibi, P.A.; Newmister, S.A.; Rayment, I.; McCormick, S.P.; Alexander, N.J.; Schmale, D.G. Bioprospecting for trichothecene-3-O-acetyltransferases in the fungal genus Fusarium yields functional enzymes that vary in their ability to modify the mycotoxin deoxynivalenol. Appl. Environ. Microbiol. 2011, 77, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.T.; Smith, T.K. Conjugation of deoxynivalenol by Alternaria alternata (54028 NRRL), Rhizopus microsporus var. rhizopodiformis (54029 NRRL) and Aspergillus oryzae (5509 NRRL). Mycotoxin Res. 2014, 28, 7–13. [Google Scholar] [CrossRef]
- He, C.; Fan, Y.; Liu, G.; Zhang, H. Isolation and identification of a strain of Aspergillus tubingensis with deoxynivalenol biotransformation capability. Int. J. Mol. Sci. 2008, 9, 2366–2375. [Google Scholar] [CrossRef]
- Tian, Y.; Tan, Y.; Liu, N.; Yan, Z.; Liao, Y.; Chen, J.; De Saeger, S.; Yang, H.; Zhang, Q.; Wu, A. Detoxification of deoxynivalenol via glycosylation represents novel insights on antagonistic activities of Trichoderma when confronted with Fusarium graminearum. Toxins 2016, 8, 335. [Google Scholar] [CrossRef]
- Islam, R.; Zhou, T.; Young, J.C.; Goodwin, P.H.; Pauls, K.P. Aerobic and anaerobic de-epoxydation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil. World J. Microbiol. Biotechnol. 2011, 28, 7–13. [Google Scholar] [CrossRef]
- Yu, H.; Zhou, T.; Gong, J.; Young, C.; Su, X.; Li, X.-Z.; Zhu, H.; Tsao, R.; Yang, R. Isolation of deoxynivalenol-transforming bacteria from the chicken intestines using the approach of PCR-DGGE guided microbial selection. BMC Microbiol. 2010, 10, 182. [Google Scholar] [CrossRef]
- Li, X.-Z.; Zhu, C.; de Lange, C.; Zhou, T.; He, J.; Yu, H.; Gong, J.; Young, J.C. Efficacy of detoxification of deoxynivalenol-contaminated corn by Bacillus sp LS100 in reducing the adverse effects of the mycotoxin on swine growth performance. Food Addit. Contam. Part A 2011, 28, 894–901. [Google Scholar] [CrossRef]
- Gao, X.; Mu, P.; Wen, J.; Sun, Y.; Chen, Q.; Deng, Y. Detoxification of trichothecene mycotoxins by a novel bacterium, Eggerthella sp. DII-9. Food Chem Toxicol. 2018, 112, 310–319. [Google Scholar] [CrossRef]
- Gao, X.; Mu, P.; Zhu, X.; Chen, X.; Tang, S.; Wu, Y.; Miao, X.; Wang, X.; Wen, J.; Deng, Y. Dual function of a novel bacterium, Slackia sp. D-G6: Detoxifying deoxynivalenol and producing the natural estrogen analogue, equol. Toxins 2020, 12, 85. [Google Scholar] [CrossRef]
- Binder, J.; Horvath, E.M.; Schatzmayr, G.; Ellend, N.; Danner, H.; Krska, R.; Braun, R. Screening for deoxynivalenol-detoxifying anaerobic rumen microorganisms. Cereal Res. Commun. 1997, 25, 343–346. [Google Scholar] [CrossRef]
- Fuchs, E.; Binder, E.M.; Heidler, D.; Krska, R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit. Contam. 2002, 19, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Schatzmayr, G.; Zehner, F.; Täubel, M.; Schatzmayr, D.; Klimitsch, A.; Loibner, A.P.; Binder, E.M. Microbiologicals for deactivating mycotoxins. Mol. Nutr. Food Res. 2006, 50, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Sayyari, A.; Fæste, C.K.; Hansen, U.; Uhlig, S.; Framstad, T.; Schatzmayr, D.; Sivertsen, T. Effects and biotransformation of the mycotoxin deoxynivalenol in growing pigs fed with naturally contaminated pelleted grains with and without the addition of Coriobacteriaceum DSM 11798. Food Addit. Contam. Part A 2018, 35, 1394–1409. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, J.; Huang, L.; Chen, H.; Wang, C. Effects of adding Clostridium sp. WJ06 on intestinal morphology and microbial diversity of growing pigs fed with natural deoxynivalenol contaminated wheat. Toxins 2017, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Shima, J.; Takase, S.; Takahashi, Y.; Iwai, Y.; Fujimoto, H.; Yamazaki, M.; Ochi, K. Novel detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated by enrichment culture. Appl. Environ. Microbiol. 1997, 63, 3825–3830. [Google Scholar] [CrossRef] [PubMed]
- He, J.W.; Hassan, Y.I.; Perilla, N.; Li, X.-Z.; Boland, G.J.; Zhou, T. Bacterial epimerization as a route for deoxynivalenol detoxification: The influence of growth and environmental conditions. Front. Microbiol. 2016, 7, 572. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Y.I.; He, J.W.; Perilla, N.; Tang, K.; Karlovsky, P.; Zhou, T. The enzymatic epimerization of deoxynivalenol by Devosia mutans proceeds through the formation of 3-keto-DON intermediate. Sci. Rep. 2017, 7, 6929. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, Y.; Ji, F.; Xu, L.; Yu, M.; Shi, J.; Xu, J. Biodegradation of deoxynivalenol and its derivatives by Devosia insulae A16. Food Chem. 2019, 276, 436–442. [Google Scholar] [CrossRef]
- Gao, H.; Niu, J.; Yang, H.; Lu, Z.; Zhou, L.; Meng, F.; Lu, F.; Chen, M. Epimerization of deoxynivalenol by the Devosia strain A6-243 assisted by pyrroloquinoline quinone. Toxins 2022, 14, 16. [Google Scholar] [CrossRef]
- He, W.J.; Shi, M.M.; Yang, P.; Huang, T.; Zhao, Y.; Wu, A.B.; Dong, W.B.; Li, H.P.; Zhang, J.B.; Liao, Y.C. A quinone-dependent dehydrogenase and two NADPH-dependent aldo/keto reductases detoxify deoxynivalenol in wheat via epimerization in a Devosia strain. Food Chem. 2020, 321, 126703. [Google Scholar] [CrossRef] [PubMed]
- Sato, I.; Ito, M.; Ishizaka, M.; Ikunaga, Y.; Sato, Y.; Yoshida, S.; Koitabashi, M.; Tsushima, S. Thirteen novel deoxynivalenol-degrading bacteria are classified within two genera with distinct degradation mechanisms. FEMS Microbiol. Lett. 2011, 327, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Morimura, H.; Uesaka, K.; Ito, M.; Yoshida, S.; Koitabashi, M.; Tsushima, S.; Sato, I. Draft genome sequence of deoxynivalenol-degrading actinomycete Nocardioides sp. strain LS1, isolated from wheat leaves in Japan. Microbiol. Resour. Announc. 2019, 8, e01650-18. [Google Scholar] [CrossRef] [PubMed]
- Ikunaga, Y.; Sato, I.; Grond, S.; Numaziri, N.; Yoshida, S.; Yamaya, H.; Hiradate, S.; Hasegawa, M.; Toshima, H.; Koitabashi, M.; et al. Nocardioides sp. strain WSN05-2, isolated from a wheat field, degrades deoxynivalenol, producing the novel intermediate 3-epi deoxynivalenol. Appl. Microbiol. Biotechnol. 2011, 89, 419–427. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, H.; Qin, X.; Wang, X.; Wang, Y.; Bin, Y.; Xie, X.; Zheng, F.; Luo, H. Biodegradation of deoxynivalenol by Nocardioides sp. ZHH-013: 3-keto-deoxynivalenol and 3-epi-deoxynivalenol as intermediate products. Front. Microbiol. 2021, 12, 658421. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Zhao, C.; Han, Y.; Liu, Y.; Zhang, X. Isolation and characterisation of a novel deoxynivalenol- transforming strain Paradevosia shaoguanensis DDB001 from wheat field soil. Lett. Appl. Microbiol. 2017, 65, 414–422. [Google Scholar] [CrossRef]
- He, W.J.; Shi, M.M.; Yang, P.; Huang, T.; Yuan, Q.S.; Yi, S.Y.; Wu, A.B.; Li, H.P.; Gao, C.B.; Zhang, J.B.; et al. Novel soil bacterium strain Desulfitobacterium sp. PGC-3-9 detoxifies trichothecene mycotoxins in wheat via de-epoxidation under aerobic and anaerobic conditions. Toxins 2020, 12, 363. [Google Scholar] [CrossRef]
- Wang, S.; Hou, Q.; Guo, Q.; Zhang, J.; Sun, Y.; Wei, H.; Shen, L. Isolation and characterization of a deoxynivalenol-degrading bacterium Bacillus licheniformis YB9 with the capability of modulating intestinal microbial flora of mice. Toxins 2020, 12, 184. [Google Scholar] [CrossRef]
- Zhang, J.; Qin, X.; Guo, Y.; Zhang, Q.; Ma, Q.; Ji, C.; Zhao, L. Enzymatic degradation of deoxynivalenol by a novel bacterium, Pelagibacterium halotolerans ANSP101. Food Chem. Toxicol. 2020, 140, 111276. [Google Scholar] [CrossRef]
- He, W.J.; Zhang, L.; Yi, S.Y.; Tang, X.L.; Yuan, Q.S.; Guo, M.W.; Wu, A.B.; Qu, B.; Li, H.P.; Liao, Y.C. An aldo-keto reductase is responsible for Fusarium toxin-degrading activity in a soil Sphingomonas strain. Sci. Rep. 2017, 7, 9549. [Google Scholar] [CrossRef]
- Murtaza, B.; Li, X.; Dong, L.; Saleemi, M.R.; Iqbal, M.; Majeed, S.; Ali, A.; Li, G.; Jin, B.; Wang, L.; et al. In-vitro assessment of a novel plant rhizobacterium, Citrobacter freundii, for degrading and biocontrol of food mycotoxin deoxynivalenol. Toxicon 2023, 27, 107095. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Zhong, L.; Gao, H.; Lu, Z.; Bie, X.; Zhao, H.; Zhang, C.; Lu, F. Detoxification of deoxynivalenol by a mixed culture of soil bacteria with 3-epi-deoxynivalenol as the main intermediate. Front. Microbiol. 2019, 10, 2172. [Google Scholar] [CrossRef]
- Carere, J.; Hassan, Y.I.; Lepp, D.; Zhou, T. The enzymatic detoxification of the mycotoxin deoxynivalenol: Identification of DepA from the DON epimerization pathway. Microb. Biotechnol. 2018, 11, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- Carere, J.; Hassan, Y.I.; Lepp, D.; Zhou, T. The identification of DepB: An enzyme responsible for the final detoxification step in the deoxynivalenol epimerization pathway in Devosia mutans 17-2-E-8. Front. Microbiol. 2018, 9, 1573. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zhang, J.; Liu, Y.; Guo, Y.; Tang, Y.; Zhang, Q.; Ma, Q.; Ji, C.; Zhao, L. A quinoprotein dehydrogenase from Pelagibacterium halotolerans ANSP101 oxidizes deoxynivalenol to 3-keto-deoxynivalenol. Food Control 2022, 136, 108834. [Google Scholar] [CrossRef]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Vekiru, E.; Krska, R.; Schatzmayr, G.; Moll, W.-D.; Grabherr, R. Degradation of fumonisin B1 by the consecutive action of two bacterial enzymes. J. Biotechnol. 2010, 145, 120–129. [Google Scholar] [CrossRef]
- Blackwell, B.A.; Gilliam, J.T.; Savard, M.E.; Miller, D.; Duvick, J.P. Oxidative deamination of hydrolyzed fumonisin B1 (AP1) by cultures of Exophiala spinifera. Nat. Toxins 1999, 7, 31–38. [Google Scholar] [CrossRef]
- Qi, T.F.; Renaud, J.B.; McDowell, T.; Seifert, K.A.; Yeung, K.K.-C.; Sumarah, M.W. Diversity of mycotoxin-producing black aspergilli in Canadian vineyards. J. Agric. Food Chem. 2016, 64, 1583–1589. [Google Scholar] [CrossRef]
- Burgess, K.M.N.; Renaud, J.B.; McDowell, T.; Sumarah, M.W. Mechanistic insight into the biosynthesis and detoxification of fumonisin mycotoxins. ACS Chem. Biol. 2016, 11, 2618–2625. [Google Scholar] [CrossRef]
- Garnham, C.P.; Butler, S.G.; Telmer, P.G.; Black, F.E.; Renaud, J.B.; Sumarah, M.W. Identification and characterization of an Aspergillus niger amine oxidase that detoxifies intact fumonisins. J. Agric. Food Chem. 2020, 68, 13779–13790. [Google Scholar] [CrossRef]
- Duvick, J.; Rood, T.; Wang, N. Fumonisin Detoxification Enzymes. U.S. Patent 5716820, 10 February 1998. [Google Scholar]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Schatzmayr, G.; Moll, W.D.; Grabherr, R. An aminotransferase from bacterium ATCC 55552 deaminates hydrolyzed fumonisin B-1. Biodegradation 2011, 22, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, R.; Nazzi, F.; Locci, R.; Firrao, G. Degradation of fumonisin B1 by a bacterial strain isolated from soil. Biodegradation 2006, 17, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Keawmanee, P.; Rattanakreetakul, C.; Pongpisutta, R. Microbial reduction of fumonisin B1 by the new isolate Serratia marcescens 329-2. Toxins 2021, 13, 638. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Gong, A.; Liu, N.; Chen, S.; Zhao, X.; Li, X.; Chen, L.; Zhou, C.; Wang, J. Biodegradation of mycotoxin fumonisin B1 by a novel bacterial consortium SAAS79. Appl. Microbiol. Biotechnol. 2019, 103, 7129–7140. [Google Scholar] [CrossRef]
- Scientific Opinion on the safety and efficacy of fumonisin esterase (FUMzyme®) as a technological feed additive for pigs. EFSA J. 2014, 12, 3667.
- Moll, D. Enzyme technology for detoxification of mycotoxins in animal feed. In Industrial Enzyme Applications; Vogel, A., May, O., Eds.; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2019; pp. 219–254. [Google Scholar] [CrossRef]
- Grenier, B.; Schwartz-Zimmermann, H.E.; Gruber-Dorninger, C.; Dohnal, I.; Aleschko, M.; Schatzmayr, G.; Moll, W.D.; Applegate, T.J. Enzymatic hydrolysis of fumonisins in the gastrointestinal tract of broiler chickens. Poult. Sci. 2017, 96, 4342. [Google Scholar] [CrossRef] [PubMed]
- Alberts, J.; Schatzmayr, G.; Moll, W.-D.; Davids, I.; Rheeder, J.; Burger, H.-M.; Shephard, G.; Gelderblom, W. Detoxification of the fumonisin mycotoxins in maize: An enzymatic approach. Toxins 2019, 11, 523. [Google Scholar] [CrossRef]
- Duvick, J.; Maddox, J.; Gilliam, J. Compositions and Methods for Fumonisin Detoxification. PCT Patent Application WO200004158, 27 January 2000. [Google Scholar]
- Li, Z.; Wang, Y.; Liu, Z.; Jin, S.; Pan, K.; Liu, H.; Liu, T.; Li, X.; Zhang, C.; Luo, X.; et al. Biological detoxification of fumonisin by a novel carboxylesterase from Sphingomonadales bacterium and its biochemical characterization. Int. J. Biol. Macromol. 2021, 169, 18–27. [Google Scholar] [CrossRef]
- Liu, Q.; Huang, L.; Cui, Z.; Qiao, B.; Li, F.; Wang, C. FumDSB can alleviate the inflammatory response induced by fumonisin B1 in growing pigs. Food Addit. Contam. Part A 2022, 39, 1619–1633. [Google Scholar] [CrossRef]
- Hartinger, D.; Schwartz, H.; Hametner, C.; Schatzmayr, G.; Haltrich, D.; Moll, W.D. Enzyme characteristics of aminotransferase FumI of Sphingopyxis sp. MTA144 for deamination of hydrolyzed fumonisin B1. Appl. Microbiol. Biotechnol. 2011, 91, 757–768. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, J.; Zhang, M.; Pan, K.; Liu, T.; Zhang, T.; Luo, X.; Zhao, J.; Li, Z. Detoxification of fumonisins by three novel transaminases with diverse enzymatic characteristics coupled with carboxylesterase. Foods 2023, 12, 416. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Yu, S.; Yu, D.; Lin, H.; Liu, N.; Wu, A. Biodegradation of fumonisins by the consecutive action of a fusion enzyme. Toxins 2022, 14, 266. [Google Scholar] [CrossRef]
- Dänicke, S.; Valenta, H.; Döll, S.; Ganter, M.; Flachowsky, G. On the effectiveness of a detoxifying agent in preventing fusario-toxicosis in fattening pigs. Anim. Feed Sci. Technol. 2004, 114, 141–157. [Google Scholar] [CrossRef]
- Sandlin, N.; Russell Kish, D.; Kim, J.; Zaccaria, M.; Momeni, B. Current and emerging tools of computational biology to improve the detoxification of mycotoxins. Appl. Environ. Microbiol. 2022, 88, e0210221. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.; Jiang, L.; Huang, H. Improvement of the enzymatic detoxification activity towards mycotoxins through structure-based engineering. Biotechnol. Adv. 2022, 56, 107927. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, W.; Chen, C.-C.; Li, Q.; Huang, J.-W.; Ko, T.-P.; Liu, G.; Liu, W.; Peng, W.; Cheng, Y.-S.; et al. Enhanced α-zearalenol hydrolyzing activity of a mycoestrogen-detoxifying lactonase by structure-based engineering. ACS Catal. 2016, 6, 7657–7663. [Google Scholar] [CrossRef]
- Fu, X.; Xu, M.; Li, T.; Zhang, H.; Zhang, C. The improved expression and stability of zearalenone lactonohydrolase from Escherichia coli BL21 (DE3). Appl. Biochem. Microbiol. 2021, 57, 79–85. [Google Scholar] [CrossRef]
- Wang, X.; Bai, Y.; Huang, H.; Tu, T.; Wang, Y.; Wang, Y.; Luo, H.; Yao, B.; Su, X. Degradation of aflatoxin B1 and zearalenone by bacterial and fungal laccases in presence of structurally defined chemicals and complex natural mediators. Toxins 2019, 11, 609. [Google Scholar] [CrossRef]
- Xia, Y.; Wu, Z.; He, R.; Gao, Y.; Qui, Y.; Cheng, Q.; Ma, X.; Wang, Z. Simultaneous degradation of two mycotoxins enabled by a fusion enzyme in food-grade recombinant Kluyveromyces lactis. Bioresour. Bioprocess. 2021, 8, 62. [Google Scholar] [CrossRef]
- Azam, M.S.; Yu, D.; Liu, N.; Wu, A. Degrading ochratoxin A and zearalenone mycotoxins using a multifunctional recombinant enzyme. Toxins 2019, 11, 301. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Fusariotoxin Occurrence, % of Samples | ||
|---|---|---|---|
| Zearalenone | Deoxynivalenol | Fumonisins | |
| Europe | 46 | 52 | 40 |
| Middle East | 65 | 58 | 90 |
| Pacific Asia ** | 71 | 80 | 90 |
| North America ** | 32 | 69 | 73 |
| Central America | 73 | 68 | - |
| South America | 40 | 46 | 66 |
| Africa | 44 | 81 | 51 |
| Species | Type of Experiment and ZEN Concentration | ZEN Degradation Level, % | Approaches Used for Evaluation of Biotransformation Efficiency/Mechanism of Transformation | Data From |
|---|---|---|---|---|
| Bacteria | ||||
| Bacillus subtilis 168 | In Vitro, LB medium; 20 μg/L | 81 | MS analysis (lack of estrogenic-type metabolites)/Unknown (includes decarboxylation) | [44] |
| B. natto CICC 24640 | In Vitro, LB medium; 20 μg/L | 100 | MS analysis (lack of estrogenic-type metabolites)/Unknown (includes decarboxylation) | [44] |
| B. pumilis ES-21 | In Vitro, LB medium; 17.9 μg/mL | 95.7 | LC-TOF-MS/MS analysis (non-estrogenic metabolite)/Lactone ring cleavage | [45] |
| B. velezensis ANSB01E | In Vitro, LB medium; 2 μg/mL | 95 | BLAST analysis (revealing of genes encoding proteins similar to confirmed ZEN-transforming enzymes)/Unknown (possibly cracking of a dihydroxybenzene ring) | [46] |
| In Vitro, corn meal medium; 0.74 μg/mL | ~75 | [46] | ||
| Rhodococcus ruber N361 | In Vitro, LB medium; 2 μg/mL | 60 | ELISA test for ZEN metabolites, BLYES bioassay for estrogenicity (40–60% reduction)/Unknown | [49] |
| R. erythropolis NI1 | In Vitro, LB medium; 2 μg/mL | 50 | ELISA test for ZEN metabolites, BLYES bioassay for estrogenicity (40–60% reduction)/Unknown | [49] |
| R. pyridinivorans K402 | In Vitro, LB medium; 2 μg/mL | 70.11 | ELISA test for ZEN metabolites, BLYES bioassay for estrogenicity (100% ceasing)/Unknown | [49] |
| R. pyridinivorans K404 | In Vitro, LB medium; 2 μg/mL | 72.3 | ELISA test for ZEN metabolites, BLYES bioassay for estrogenicity (100% ceasing)/Unknown | [49] |
| R. pyridinivorans K408 | In Vitro, LB medium; 2 μg/mL | 80.3 | ELISA test for ZEN metabolites, BLYES bioassay for estrogenicity (100% ceasing)/Unknown | [49] |
| In Vitro, LB medium; 5 μg/mL | 87.2 | BLYES bioassay (81.75% reduction of estrogenicity) | [50] | |
| In Vivo, female rats | - | Immature uterotrophic assay (lack of estrogenicity) | [51] | |
| R. percolatus JCM 10087T | In Vitro, LB medium; 1 μg/mL | 95 | BLYES bioassay for estrogenicity (70% reduction)/Unknown | [52] |
| R. erythropolis PFA D8-1 | In Vitro | - | Bioassays with the MCF cell line and estrogen reporter yeast strain YZHB817 (estrogenicity), metabolite identification/Lactone ring cleavage | [53] |
| Acinetobacter sp. SM04 | In Vitro, M1 medium; 20 μg/mL; In Vitro, MCF-7 cells | ~100 | HPLC and MS for ZEN metabolites; MTT cell proliferation assay for estrogenic activity (no estrogenic effect)/Cracking of a dihydroxybenzene ring | [54] |
| Pseudomonas sp. ZEA-1 | In Vitro, M1 medium; 100 μg/mL | 100 | TLC analysis for ZEN metabolites/Unknown | [55] |
| In Vivo, Artemisia salina larvae | - | Toxicity tests | [55] | |
| Fungi and yeasts | ||||
| Clonostachys rosea (Gliocladeum roseum) NRRL1829 | In Vitro, 250 μg/mL | 80–90 | MS, NMR, and IR spectral analysis of ZEN metabolites; inhibition of estradiol binding to estrogen receptors to evaluate estrogenic activity/Lactone ring cleavage | [56] |
| C. rosea IFO 7063 | In Vitro, 100 μg/mL | nd * | NMR and MS analysis of ZEN metabolites; MCF-7 cell culture assay to evaluate estrogenic activity/Lactone ring cleavage | [57] |
| C. rosea GRZ7 | In Vitro, 0.5 μg/mL | 68 | Isolation of lactonohydrolase with the confirmed target activity/Lactone ring cleavage | [59] |
| Trichosporon mycotoxinivorans | In Vitro, saline medium, 10 μg/mL; In Vivo, yeast bioassayIn Vitro, estrogen receptor binding assay | 95 | LC-MS/MS, LC-DAD, TOF MS, NMR analysis of ZEN metabolites; indicator yeast bioassay and human estrogen receptor protein binding assay for estrogenicity evaluation/Cracking of a C6′ ketone carbonyl group | [65] |
| Species | Experimental Conditions and DON Concentration | DON Degradation Level, % | Approaches Used for Evaluation of Biotransformation Efficiency (Metabolite) | Data From |
|---|---|---|---|---|
| Anaerobic bacteria | ||||
| Bacillus sp. LS100 | In Vitro, AIM medium, 100 μg/mL | 100 | LS-MS analysis of DON metabolites (DOM-1) | [105] |
| In Vivo (pigs), 124 μg/g of corn | - | LS-MS analysis of DON metabolites; feed intake, weight gain, and feed efficiency to evaluate toxicity | [106] | |
| Eggerthella (Raoultibacter) sp. DII-9 | In Vitro, BHI broth, 100 μg/mL | 100 | LC-MS analysis of DON metabolites (DOM-1) | [107] |
| Slackia sp. D-G6 | In Vitro, BHI broth, 100 μg/mL | >90 | HPLC analysis of DON metabolites (DOM-1) | [108] |
| Coriobacteriaceum DSM 11798 | In Vitro, M10 medium, 100 ppm | 100 | RP-HPLC and TLC analysis of DON metabolites; LPA assay for toxicity evaluation (DOM-1) | [109,111] |
| Clostridium sp. WJ06 | In Vitro, L10 medium, 20 ppm | >90 | LC-MS/MS analysis of DON metabolites (DOM-1) | [113] |
| In Vivo (pigs), 1.9 mg/kg of feed | - | Toxicity evaluation (growth performance, intestinal morphology, relative organ weight, intestinal flora) | [113] | |
| Aerobic bacteria | ||||
| E3-39 isolate (Agrobacterium–Rhizobium group) | In Vitro, BYE medium, 200 μg/mL | 100 | TLC and NMR analysis of DON metabolites; mouse spleen lymphocyte proliferation assay to evaluate immunosuppressive activity (3-keto-DON) | [114] |
| Devosia mutans 17-2-E-8 | In Vitro, CMB medium, 100 μg/mL | 95 | HPLC analysis of DON metabolites (3-epi-DON) | [115] |
| Devosia insulae A16 | In Vitro, MSM medium, 20 μg/mL | 88 | HPLC, LC-TOF-MS and NMR analyses of DON metabolites (3-keto-DON) | [117] |
| Devosia A6-243 | In Vitro, MSM medium + PQQ, 100 μg/mL | 100 | QTOF LC–MS and NMR analyses of DON metabolites (3-epi-DON) | [118] |
| In Vitro, wheat grain, 6.7 µg/g | 100 | QTOF LC–MS and NMR analyses of DON metabolites (3-epi-DON) | [118] | |
| Devosia sp. D6-9 | In Vitro, 500 μg/mL | 100 | HPLS and GC-MS identification of DON metabolites (3-keto-DON and 3-epi-DON) | [119] |
| In Vitro, wheat grain, 11.5 µg/g | 100 | HPLS and GC-MS identification of DON metabolites (3-keto-DON and 3-epi-DON) | [119] | |
| Devosia sp. SS5, RS1, NKK1, NKJ1 | In Vitro, MSM medium, 100 μg/mL | ~99.5% | HPLC analysis of DON metabolites (3-epi-DON) | [120] |
| Nocardioides sp. PFS1, YMN1, SS1, SS2, SS3, SS4, LS1, LS2, YUL1 | In Vitro, MSM medium; 100 μg/mL (pre-incubation with DON) | ~99.5% | HPLC analysis of DON metabolites (3-epi-DON) | [120] |
| Nocardioides sp. WSN05-2 | In Vitro, MSM medium, 1000 μg/mL | 100 | LC-MS and NMR analyses of DON metabolites (3-epi-DON) | [122] |
| In Vitro, wheat grain, 2 μg/100 grains | 100 | LC-MS and NMR analyses of DON metabolites (3-epi-DON) | [122] | |
| Nocardioides sp. ZHH-013 | In Vitro, MSM medium, 168.74 μM | 80 | LC-MS analysis of DON metabolites (3-epi-DON) | [123] |
| Paradevosia shaoguanensis DDB001 | In Vitro, MSM medium, 200 μg/mL | 100 | LC-MS analysis of DON metabolites (3-epi-DON) | [124] |
| Desulfitobacterium sp. PGC-3-9 | In Vitro, MMYPF medium, 500 μg/mL | 99/95 (anaerobic/aerobic) | GC-MS analysis of DON metabolites (DOM-1) | [125] |
| In Vitro, wheat grain, 11.2 μg/g | 92 | GC-MS analysis of DON metabolites (DOM-1) | [125] | |
| Bacillus licheniformis YB9 | In Vitro, MSM medium, 1 μg/L | 82.67 | ELISA (DON Plus Test Kit) for DON quantification, use of DON as a sole carbon source | [126] |
| Pelagibacterium halotolerans ANSP101 | In Vitro, MMB2216 medium, 50 µg/mL | 80 | UPLC-MS/MS analysis of DON degradation products (3-keto-DON) | [127] |
| Sphingomonas sp. S3-4 | In Vitro, MSM medium, 100 µg/mL | 100 | GC-MS and NMR analysis of degradation products (3-oxo-DON (=3-keto-DON) and 3-epi-DON) | [128] |
| In Vitro, wheat grain, ~115 µg/g | 100 | GC-MS and NMR analysis of degradation products (3-oxo-DON (=3-keto-DON) and 3-epi-DON) | [128] | |
| Citrobacter freundii | In Vitro, LB broth, 10 µg/mL | 93.5 | HPLC and UPLC-MS/MS analysis of DON degradation products (3-keto-DON, DOM-1) | [129] |
| Pseudomonas sp. Y1 + Lysobacter sp. S1 | In Vitro, MSM medium, 50 µg/mL | 100 | HPLC for identification of degradation products (3-epi-DON) | [130] |
| Species | Experimental Conditions and FB1 Concentration | FB1 Degradation Level, % | Approaches Used for Evaluation of Biotransformation Efficiency | Data From |
|---|---|---|---|---|
| Sphingopyxis MTA144 | In Vitro, liquid medium | nd * | LC-MS analysis of FB1 metabolites | [133] |
| Sphingomonas sp. ATCC 5552 | In Vitro, liquid medium, 0.5–1.0 mg/mL | nd | TLC and radiochemical analysis of FB1 metabolites | [139,140] |
| Delftia/Comamonas NCB 1492 | In Vitro, liquid medium, 0.5 mg/mL | 100 | HPLC, GS-MS analysis of FB1 metabolites | [141] |
| Serratia marcescens 329-2 | In Vitro, cell-free extract, 5 μg/mL | 30.29 | ELISA assay for residual FB1, MS-based proteomic analysis for enzyme expression | [142] |
| In Vitro, cell-free extract, corn grain, 5 μg/mL | 37 | ELISA assay for residual FB1, MS-based proteomic analysis to evaluate enzyme expression | [142] | |
| Bacterial consortium SAAS79 | In Vitro, liquid medium, 50 μg/mL | ~90 | LS-MS/MS analysis of FB1 metabolites; MTT and MARC-145 bioassays to evaluate cytotoxicity | [143] |
| Exophiala spinifera 2141.10 | In Vitro, MSM medium, 0.5–1 μg/mL | nd | NMR and mass spectroscopy analysis of FB1 metabolites | [135] |
| Aspergillus welwitschiae DAOMC 250207 | nd | nd | LS-MS, NMR, isotope labeling to study FB1 metabolites; Lemna minor bioassay to evaluate toxicity | [136] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Statsyuk, N.V.; Popletaeva, S.B.; Shcherbakova, L.A. Post-Harvest Prevention of Fusariotoxin Contamination of Agricultural Products by Irreversible Microbial Biotransformation: Current Status and Prospects. BioTech 2023, 12, 32. https://doi.org/10.3390/biotech12020032
Statsyuk NV, Popletaeva SB, Shcherbakova LA. Post-Harvest Prevention of Fusariotoxin Contamination of Agricultural Products by Irreversible Microbial Biotransformation: Current Status and Prospects. BioTech. 2023; 12(2):32. https://doi.org/10.3390/biotech12020032
Chicago/Turabian StyleStatsyuk, Natalia V., Sophya B. Popletaeva, and Larisa A. Shcherbakova. 2023. "Post-Harvest Prevention of Fusariotoxin Contamination of Agricultural Products by Irreversible Microbial Biotransformation: Current Status and Prospects" BioTech 12, no. 2: 32. https://doi.org/10.3390/biotech12020032