
Ten Principles for Bird-Friendly Forestry: Conservation Approaches in Natural Forests Used for Timber Production
Abstract
:Simple Summary
Abstract
1. Introduction
2. Forest Birds as Indicators of Biodiversity and Environmental Change
3. Conservation Approaches in Natural Forests Used for Timber Production
4. Ten Principles of Bird-Friendly Forestry
4.1. Protect and Enhance Vertical Structure through Uneven-Aged Silviculture
4.2. Leave Dead and Dying Trees and Coarse Woody Debris in Different Decay Stages
4.3. Maintain Residual Large, Green Trees
4.4. Establish and Maintain Uncut Reserves, Ideally Connected by Corridors
4.5. Maximize Forest Interior through Retaining Large Contiguous Forest Tracts
4.6. Maintain Buffers around Streams, Rivers, Wetlands, and Known Nesting Areas
4.7. Maintain Horizontal Stand Structure and Enhance Vegetation Diversity in Canopy Gaps
4.8. Extend the Temporal Scale of Logging Cycles through Prolonged Rotations
4.9. Minimize Disturbance to Forests after Logging and during the Bird Breeding Season
4.10. Manage for Focal Species and Guilds
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Global Forest Resources Assessment 2010: Main Report. FAO: Rome, Italy, 2010; Available online: https://www.fao.org/3/i1757e/i1757e00.htm (accessed on 1 February 2023).
- Lehikoinen, A.; Virkkala, R. Population trends and conservation status of forest birds. In Ecology and Conservation of Forest Birds; Mikusiński, G., Roberge, J.-M., Fuller, R.J., Eds.; Cambridge University Press: Cambridge, UK, 2018; pp. 390–426. [Google Scholar]
- Betts, M.G.; Wolf, C.; Ripple, W.J.; Phalan, B.; Millers, K.A.; Duarte, A.; Butchart, S.H.M.; Levi, T. Global forest loss disproportionately erodes biodiversity in intact landscapes. Nature 2017, 547, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Imbeau, L.; Mönkkönen, M.; Desrochers, A. Long-term effects of forestry on birds of the eastern Canadian boreal forests: A comparison with Fennoscandia. Conserv. Biol. 2001, 15, 1151–1162. [Google Scholar] [CrossRef]
- Martin, J.-L.; Drapeau, P.; Fahrig, L.; Lindsay, K.F.; Kirk, D.A.; Smith, A.C.; Villard, M.-A. Birds in cultural landscapes: Actual and perceived differences between northeastern North America and western Europe. In Birds and Habitat Relationships in Changing Landscapes; Fuller, R.J., Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 481–515. [Google Scholar] [CrossRef]
- Sallabanks, R.; Arnett, E.B.; Marzluff, J.M. An evaluation of research on the effects of timber harvest on bird populations. Wildl. Soc. Bull. 2000, 28, 1144–1155. [Google Scholar]
- Angelstam, P.; Roberge, J.-M.; Lõhmus, A.; Bergmanis, M.; Brazaitis, G.; Dönz-Breuss, M.; Edenius, L.; Kosinski, Z.; Kurlavicius, P.; Lârmanis, V.; et al. Habitat modelling as a tool for landscape-scale conservation—A review of parameters for focal forest birds. Ecol. Bull. 2004, 51, 427–453. [Google Scholar]
- Sallabanks, R.; Arnett, E.B. Accommodating Birds in Managed Forests of North America: A Review of Bird-Forestry Relationships. USDA Forest Service General Technical Report, PSW-GTR-191. 2005. Available online: https://www.fs.usda.gov/research/treesearch/31705 (accessed on 8 November 2022).
- Angelstam, P.; Manton, M.; Green, M.; Jonsson, B.-G.; Mikusiński, G.; Svensson, J.; Sabatini, F.M. Sweden does not meet agreed national and international forest biodiversity targets: A call for adaptive landscape planning. Landsc. Urban Plan. 2020, 202, 103838. [Google Scholar] [CrossRef]
- Zhao, Q.; Azeria, E.T.; Le Blanc, M.-L.; Lemaître, J.; Fortin, D. Landscape-scale disturbances modified bird community dynamics in successional forest environment. PLoS ONE 2013, 8, e81358. [Google Scholar] [CrossRef] [PubMed]
- Buřivalová, Z.; Şekercioğlu, C.H.; Koh, L.P. Thresholds of logging intensity to maintain tropical forest biodiversity. Curr. Biol. 2014, 24, 1893–1898. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, B.G.; Svensson, J.; Mikusiński, G.; Manton, M.; Angelstam, P. European Union’s last intact forest landscape is at a value chain crossroad between multiple use and intensified wood production. Forests 2019, 10, 564. [Google Scholar] [CrossRef] [Green Version]
- Drapeau, P.; Leduc, A.; Giroux, J.-F.; Savard, J.-P.; Bergeron, Y.; Vickery, W. Landscape-scale disturbances and changes in bird communities of boreal mixed-wood forests. Ecol. Monogr. 2000, 70, 423–444. [Google Scholar] [CrossRef]
- Venier, L.A.; Thompson, I.D.; Fleming, R.; Malcolm, J.; Aubin, I.; Trofymow, J.A.; Langor, D.; Sturrock, R.; Patry, C.; Outerbridge, R.O.; et al. Effects of natural resource development on the terrestrial biodiversity of Canadian boreal forests. Environ. Rev. 2014, 22, 457–490. [Google Scholar] [CrossRef]
- Buřivalová, Z.; Rosin, C.; Buchner, J.; Radeloff, V.C.; Ocampo-Peñuela, N. Conservation responsibility for bird species in tropical logged forests. Conserv. Lett. 2022, 15, e12903. [Google Scholar] [CrossRef]
- Convention on Biological Diversity. 2010. Available online: https://www.cbd.int/forest/definitions.shtml (accessed on 12 February 2023).
- Smith, D.M. The Practice of Silviculture; Wiley: New York, NY, USA, 1986. [Google Scholar]
- USDA Forest Service. Ottawa National Forest Land and Resource Management Plan Appendix C: Harvest Cutting Methods. 2006. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5110681.pdf (accessed on 7 November 2022).
- Angelstam, P.; Naumov, V.; Elbakidze, M.; Manton, M.; Priednieks, J.; Rendenieks, Z. Wood production and biodiversity conservation are rival forestry objectives in Europe’s Baltic Sea Region. Ecosphere 2018, 9, e02119. [Google Scholar] [CrossRef] [Green Version]
- Naumov, V.; Manton, M.; Elbakidze, M.; Rendenieks, Z.; Priednieks, J.; Uhlianets, S.; Yamelynets, T.; Anton Zhivotov, A.; Angelstam, P. How to reconcile wood production and biodiversity conservation? The Pan-European boreal forest history gradient as an “experiment”. J. Environ. Manag. 2018, 218, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Czeszczewik, D.; Zub, K.; Stanski, T.; Sahel, M.; Kapusta, A.; Walankiewicz, W. Effects of forest management on bird assemblages in the Bialowieza Forest, Poland. iForest 2014, 8, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Mönkkönen, M. Managing Nordic boreal forest landscapes for biodiversity: Ecological and economic perspectives. Biodivers. Conserv. 1999, 8, 85–99. [Google Scholar] [CrossRef]
- Seymour, R.S.; White, A.S.; de Maynadier, P.G. Natural disturbance regimes in northeastern North America—Evaluating silvicultural systems using natural scales and frequencies. For. Ecol. Manag. 2002, 155, 357–367. [Google Scholar] [CrossRef]
- Franklin, J.F.; Spies, T.A.; Pelt, R.; Van Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C.; et al. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Angelstam, P.; Kuuluvainen, T. Boreal forest disturbance regimes, successional dynamics and landscape structures—A European perspective. Ecol. Bull. 2004, 51, 117–136. [Google Scholar]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [Green Version]
- Viljur, M.; Abella, S.R.; Adámek, M.; Alencar, J.B.R.; Barber, N.A.; Beudert, B.; Burkle, L.A.; Cagnolo, L.; Campos, B.R.; Chao, A.; et al. The effect of natural disturbances on forest biodiversity: An ecological synthesis. Biol. Rev. 2022, 97, 1930–1947. [Google Scholar] [CrossRef]
- Jokimäki, J.; Huhta, E. Effects of landscape matrix and habitat structure on a bird community in northern Finland: A multi-scale approach. Ornis Fenn. 1996, 73, 97–113. [Google Scholar]
- Forsman, J.T.; Reunanen, P.; Jokimäki, J.; Mönkkönen, M. The effects of small-scale disturbance on forest birds: A meta-analysis. Can. J. For. Res. 2010, 40, 1833–1842. [Google Scholar] [CrossRef]
- Mönkkönen, M.; Juutinen, A.; Mazziotta, A.; Miettinen, K.; Podkopaev, D.; Reunanen, P.; Salminen, H.; Tikkanen, O.-P. Spatially dynamic forest management to sustain biodiversity and economic returns. J. Environ. Manag. 2014, 134, 80–89. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Ramage, B.S. Silviculture in an uncertain world: Utilizing multi-aged management systems to integrate disturbance. Forestry 2013, 86, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Pommerening, A.; Murphy, S.T. A review of the history, definitions and methods of continuous cover forestry with special attention to afforestation and restocking. Forestry 2004, 77, 27–44. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Martínez Pastur, G.; Messier, C.; et al. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Kuuluvainen, T.; Angelstam, P.; Frelich, L.; Jõgiste, K.; Koivula, M.; Kubota, Y.; Lafleur, B.; Macdonald, E. Natural disturbance-based forest management: Moving beyond retention and continuous-cover forestry. Front. For. Glob. Chang. 2021, 4, 629020. [Google Scholar] [CrossRef]
- Diaci, J.; Kerr, G.; O’Hara, K. Twenty-first century forestry: Integrating ecologically based, uneven-aged silviculture with increased demands on forests. Forestry 2011, 84, 463–465. [Google Scholar] [CrossRef] [Green Version]
- Larsen, J.B.; Angelstam, P.; Bauhus, J.; Carvalho, J.F.; Diaci, J.; Dobrowolska, D.; Gazda, A.; Gustafsson, L.; Krumm, F.; Knoke, T.; et al. Closer-to-Nature Forest Management: From Science to Policy 12; European Forest Institute: Joensuu, Finland, 2022. [Google Scholar] [CrossRef]
- Sharma, S.; Kreye, M.M. Social value of bird conservation on private forest lands in Pennsylvania, USA. Ecol. Econ. 2022, 196, 107426. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Hobbs, R.J.; Montague Drake, R.; Alexandra, J.; Bennett, A.; Burgman, M.; Cale, P.; Calhoun, A.; Cramer, V.; Cullen, P.; et al. A checklist for ecological management of landscapes for conservation. Ecol. Lett. 2008, 11, 78–91. [Google Scholar] [CrossRef]
- Karr, J.R. Interactions between forest birds and their habitats: A comparative synthesis. In Biogeography and Ecology of Forest Bird Communities; Keast, A., Ed.; SPB Academic Publishing: The Hague, The Netherlands, 1990; pp. 379–386. [Google Scholar]
- Jokimäki, J.; Solonen, T. Habitat associations of old forest bird species in managed boreal forests characterized by forest inventory data. Ornis Fenn. 2011, 88, 57–70. [Google Scholar]
- Virkkala, R. Bird species dynamics in a managed southern boreal forest in Finland. For. Ecol. Manag. 2004, 195, 151–163. [Google Scholar] [CrossRef]
- Betts, M.G.; Villard, M.-A. Landscape thresholds in species occurrence as quantitative targets in forest management: Generality in space and time? In Setting Conservation Targets in Managed Forest Landscapes; Villard, M.-A., Jonsson, B.-G., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 185–206. [Google Scholar]
- Lindenmayer, D.B.; Franklin, J.F.; Lõhmus, A.; Baker, S.C.; Bauhus, J.; Beese, W.; Brodie, A.; Kiehl, B.; Kouki, J.; Martínez Pastur, G.; et al. A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv. Lett. 2012, 5, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Drapeau, P.; Villard, M.-A.; Leduc, A.; Hannon, S.J. Natural disturbance regimes as templates for the response of bird species assemblages to contemporary forest management. Divers. Distrib. 2016, 22, 385–399. [Google Scholar] [CrossRef] [Green Version]
- Lõhmus, A.; Nellis, R.; Pullerits, M.; Leivits, M. The potential for long-term sustainability in seminatural forestry: A broad perspective based on woodpecker populations. Environ. Manag. 2016, 57, 558–571. [Google Scholar] [CrossRef]
- Kuuluvainen, T. Forest management and biodiversity conservation based natural ecosystem dynamics in northern Europe: The complexity challenge. AMBIO 2009, 38, 209–315. [Google Scholar] [CrossRef]
- Angelstam, P.; Axelsson, R.; Elbakidze, M.; Laestadius, L.; Lazdinis, M.; Nordberg, M.; Pătru-Stupariu, I.; Smith, M. Knowledge production and learning for sustainable forest management on the ground: Pan-European landscapes as a time machine. Forestry 2011, 84, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Partners in Flight. Forests for the Birds Webinar Series. 2023. Available online: https://partnersinflight.org/resources/forest-for-the-birds-webinar-series/ (accessed on 5 September 2022).
- Arcilla, N.; Holbech, L.H.; O’Donnell, S. Severe declines of forest understory birds follow illegal logging in Ghana, West Africa. Biol. Conserv. 2015, 188, 41–49. [Google Scholar] [CrossRef]
- Carrillo-Rubio, E.; Kéry, M.; Morreale, S.J.; Sullivan, P.J.; Gardner, B.; Cooch, E.G.; Lassoie, J.P. Use of multispecies occupancy Models to evaluate the response of bird communities to forest degradation associated with logging. Conserv. Biol. 2014, 28, 1034–1044. [Google Scholar] [CrossRef]
- Burton, P.J.; Messier, C.; Weetman, G.F.; Prepas, E.E.; Adamowicz, W.L.; Tittler, R. The current state of boreal forestry and the drive for change. In Towards Sustainable Management of the Boreal Forest; Burton, P.J., Messier, C., Smith, D.W., Adamowicz, W.L., Eds.; NRC Research Press: Ottawa, ON, Canada, 2003; Chapter 1; pp. 1–40. [Google Scholar]
- Niederberger, M.; Spranger, J. Delphi Technique in health sciences: A map. Front. Public Health 2020, 8, 457. [Google Scholar] [CrossRef]
- Stokland, J. Representativeness and efficiency of bird and insect conservation in Norwegian boreal forest reserves. Conserv. Biol. 1997, 11, 101–111. [Google Scholar] [CrossRef]
- Niemi, G.J.; Hanowski, J.M.; Helle, P.; Howe, R.W.; Mönkkönen, M.; Venier, L.A.; Welsh, D.A. Ecological sustainability of birds in boreal forests. Ecol. Soc. 1998, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Voigt, D.; Baker, J.; Thompson, I.; Rempel, R. Forest vertebrate responses to landscape-level changes in Ontario. In Ecology of a Managed Terrestrial Landscape: Patterns and Processes of Forest Landscapes in Ontario; Perera, A., Euler, D., Thompson, I., Eds.; UBC Press: Vancouver, BC, Canada, 2000; pp. 198–233. [Google Scholar]
- Roberge, J.M.; Angelstam, P. Indicator species among resident forest birds—A cross-regional evaluation in northern Europe. Biol. Conserv. 2006, 130, 134–147. [Google Scholar] [CrossRef]
- Venier, L.A.; Pearce, J.L. Birds as indicators of sustainable forest management. For. Chron. 2004, 80, 61–66. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A. The impact of conservation on the status of the world’s vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef] [Green Version]
- Järvinen, O.; Väisänen, R.A. Changes in bird populations as criteria of environmental changes. Holarct. Ecol. 1979, 2, 75. [Google Scholar] [CrossRef]
- Fraixedas, S.; Lindén, A.; Piha, M.; Cabeza, M.; Gregory, R.; Lehikoinen, A. A state-of-the-art review on birds as indicators of biodiversity: Advances, challenges, and future directions. Ecol. Indic. 2020, 118, 106728. [Google Scholar] [CrossRef]
- Gregory, R.D.; Vorisek, P.; Van Strien, A.; Gmelig Meyling, A.W.; Jiguet, F.; Fornasari, L.; Reif, J.; Chylarecki, P.; Burfield, I.J. Population trends of widespread woodland birds in Europe. IBIS 2007, 149, 78–97. [Google Scholar] [CrossRef]
- Voříšek, P.; Klvaňová, A.; Wotton, S.; Gregory, R.D. A Best Practice Guide for Wild Bird Monitoring Schemes; Czech Society for Ornithology (CSO): Prague, Czech Republic, 2008; 150p. [Google Scholar]
- Pan-European Common Bird Monitoring Scheme (PECBMS). 2022. Available online: https://pecbms.info/ (accessed on 14 April 2023).
- Finnish Common Forest Bird Indicator. 2023. Available online: https://www.biodiversity.fi/en/habitats/forests/fo10-forest-birds (accessed on 22 February 2023).
- Rosenberg, K.V.; Kennedy, J.A.; Dettmers, R.; Ford, R.P.; Reynolds, D.; Alexander, J.D.; Beardmore, C.J.; Blancher, P.J.; Bogart, R.E.; Butcher, G.S.; et al. Partners in Flight Landbird Conservation Plan: 2016 Revision for Canada and Continental United States. Partners in Flight Science Committee. 2016. Available online: https://www.partnersinflight.org/wp–content/uploads/2016/08/pif-continental-plan-final-spread-single.pdf (accessed on 25 May 2023).
- IUCN Red List. 2023. Available online: https://www.iucnredlist.org/en (accessed on 15 May 2023).
- European Environment Agency. 2023. Available online: https://www.eea.europa.eu/data-and-maps/data/european-red-lists-7 (accessed on 3 May 2023).
- Committee on the Status of Endangered Wildlife in Canada (COSEWIC). 2022. Available online: https://www.cosewic.ca/index.php/en-ca/assessment-process/detailed-version-december-2022.html (accessed on 9 April 2023).
- US Fish and Wildlife Service. 2023. Available online: https://fws.gov/program/endangered-species (accessed on 16 April 2023).
- Billerman, S.M.; Keeney, B.K.; Rodewald, P.G.; Schulenberg, T.S. (Eds.) Birds of the World; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2022; Available online: https://birdsoftheworld-org.proxy.birdsoftheworld.org/bow/home (accessed on 25 May 2023).
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef]
- Schäublin, S.; Bollmann, K. Winter habitat selection and conservation of Hazel Grouse (Bonasa bonasia) in mountain forests. J. Ornithol. 2011, 152, 179–192. [Google Scholar] [CrossRef]
- Uimaniemi, L.; Orell, M.; Mönkkönen, M.; Huhta, E.; Jokimäki, J.; Lumme, J. Genetic diversity in the Siberian Jay Perisoreus infaustus in fragmented old-growth forests of Fennoscandia. Ecography 2000, 23, 669–677. [Google Scholar] [CrossRef]
- Robbins, C.S.; Fitzpatrick, J.W.; Hamel, P.B. A warbler in trouble: Dendroica cerulea. In Ecology and Conservation of Neotropical Migrant Landbirds; Hagan, J.M., III, Johnston, D.W., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1992; pp. 549–562. [Google Scholar]
- Angelstam, P.; Mikusinski, G. Woodpecker assemblages in natural and managed boreal and hemiboreal forest—A review. Ann. Zool. Fenn. 1994, 31, 157–172. [Google Scholar]
- Crocker-Bedford, D.C. Goshawk reproduction and forest management. Wildl. Soc. Bull. 1990, 18, 262–269. [Google Scholar]
- Treinys, R.; Lõhmus, A.; Stoncius, D.; Skuja, S.; Drobelis, E.; Šablevičius, B.; Rumbutis, S.; Dementavičius, D.; Naruševičius, V.; Petraška, A. At the border of ecological change: Status and nest sites of Lithuanian Black Stork Ciconia nigra population 2000–2006 versus 1976–1992. J. Ornithol. 2008, 149, 75–81. [Google Scholar] [CrossRef]
- Strazds, M. Conservation Ecology of the Black Stork in Latvia. Ph.D. Thesis, University of Latvia, Riga, Latvia, 2011; 96p. [Google Scholar]
- Bond, M.L.; Chi, T.Y.; Bradley, C.M.; DellaSala, D.A. Forest management, barred owls, and wildfire in northern spotted owl territories. Forests 2022, 13, 1730. [Google Scholar] [CrossRef]
- Drapeau, P.; Nappi, A.; Imbeau, L.; Saint-Germain, M. Standing deadwood for keystone bird species in the eastern boreal forest: Managing for snag dynamics. For. Chron. 2009, 85, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Spühler, L.; Krüsi, B.O.; Pasinelli, G. Do Oaks Quercus spp., dead wood and fruiting Common Ivy Hedera helix affect habitat selection of the middle-spotted woodpecker Dendrocopos medius. Bird Study 2015, 62, 115–119. [Google Scholar] [CrossRef]
- Pettersson, B. Extinction of an isolated population of the middle-spotted woodpecker Dendrocopos medius (L.) in Sweden and its relation to general theories on extinction. Biol. Conserv. 1985, 32, 335–353. [Google Scholar] [CrossRef]
- Troy, J.R.; Jones, C.D. The ongoing narrative of Ivory-billed Woodpecker rediscovery and support for declaring the species extinct. IBIS 2023, 165, 340–351. [Google Scholar] [CrossRef]
- Kelly, J.F.; Pletschet, S.M.; David, M.; Leslie, D.M., Jr. Habitat associations of Red-cockaded Woodpecker cavity trees in an old-growth forest of Oklahoma. J. Wildl. Manag. 1993, 57, 122–128. [Google Scholar] [CrossRef]
- Kilgo, J.C.; Vukovich, M.A. Can snag creation benefit a primary cavity nester: Response to an experimental pulse in snag abundance. Biol. Conserv. 2014, 171, 21–28. [Google Scholar] [CrossRef]
- Virkkala, R.; Lehikoinen, A. Birds on the move in the face of climate change: High species turnover in northern Europe. Ecol. Evol. 2017, 7, 8201–8209. [Google Scholar] [CrossRef] [PubMed]
- Rosamond, K.M.; Goded, S.; Soultan, A.; Kaplan, R.H.; Glass, A.; Kim, D.H.; Arcilla, N. Not singing in the rain: Linking migratory songbird declines with increasing precipitation and brood parasitism vulnerability. Front. Ecol. Evol. 2020, 8, 536769. [Google Scholar] [CrossRef]
- Jirinec, V.; Burner, R.C.; Aramal, B.R.; Bierregaard, R., Jr.; Fernández-Arellano, G.; Hernández-Palma, A.; Johnson, E.I.; Lovejoy, T.E.; Powell, L.L.; Rutt, C.L.; et al. Morphological consequences of climate change for resident birds in intact Amazonian rainforest. Sci. Adv. 2021, 7, abk1743. [Google Scholar] [CrossRef] [PubMed]
- Van Tatenhove, A.; Filiberti, E.; Sillett, T.S.; Rodenhouse, N.; Hallworth, M. Climate-related distribution shifts of migratory songbirds and sciurids in the White Mountain National Forest. Forests 2019, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Virkkala, R. Long-term decline of southern boreal forest birds: Consequence of habitat alteration or climate change? Biodivers. Conserv. 2016, 25, 151–167. [Google Scholar] [CrossRef]
- Ram, D.; Axelsson, A.-L.; Green, M.; Smith, H.G.; Lindström, Å. What drives current population trends in forest birds—Forest quantity, quality or climate? A large-scale analysis from northern Europe. For. Ecol. Manag. 2017, 385, 177–188. [Google Scholar] [CrossRef]
- Strazds, M.; Ķerus, V. Conservation Action Plan for Hazel Grouse (Bonasa bonasia) 2017–2026 [In Latvia]. Latvijas Ornitoloģijas biedrība, Rīga. In Latvian with English Summary; 2017. Available online: https://www.daba.gov.lv/lv/media/5922/download?attachment (accessed on 4 November 2022).
- Forsman, J.T.; Reunanen, P.; Jokimäki, J.; Mönkkönen, M. Effects of canopy gap disturbance on forest birds in boreal forests. Ann. Zool. Fenn. 2013, 50, 316–326. [Google Scholar] [CrossRef]
- Biber, P.; Borges, J.G.; Moshammer, R.; Barreiro, S.; Botequim, B.; Brodrechtová, Y.; Brukas, V.; Chirici, G.; Cordero-Debets, R.; Corrigan, E.; et al. How sensitive are ecosystem services in European forest landscapes to silvicultural treatment? Forests 2015, 6, 1666–1695. [Google Scholar] [CrossRef] [Green Version]
- Morissette, J.L.; Cobb, T.P.; Brigham, R.M.; James, P.C. The response of boreal forest songbird communities to fire and post-fire harvesting. Can. J. For. Res. 2002, 32, 2169–2183. [Google Scholar] [CrossRef]
- Verner, J.; Larson, T.A. Richness of breeding bird species in mixed-conifer forests of the Sierra Nevada, California. Auk 1989, 106, 447–463. [Google Scholar]
- Vanha-Majamaa, I.; Jalonen, J. Green tree retention in Fennoscandian forestry. Scand. J. For. Res. 2001, 3, 79–90. [Google Scholar] [CrossRef]
- Rosenvald, R.; Lõhmus, A. For what, when, and where is green-tree retention better than clear-cutting? A review of the biodiversity aspects. For. Ecol. Manag. 2008, 255, 1–15. [Google Scholar] [CrossRef]
- Linder, P.; Elfving, B.; Zackrisson, O. Stand structure and successional trends in virgin boreal forest reserves in Sweden. For. Ecol. Manag. 1997, 98, 17–33. [Google Scholar] [CrossRef]
- Schmiegelow, F.K.A.; Mönkkönen, M. Habitat loss and fragmentation in dynamic landscapes: Avian perspectives from the boreal forest. Ecol. Appl. 2002, 12, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Huhta, E.; Jokimäki, J.; Rahko, P. Breeding success of pied flycatchers in artificial forest edges: The effect of a suboptimally shaped foraging area. Auk 1999, 116, 528–535. [Google Scholar] [CrossRef]
- Angelstam, P.; Andersson, L. Estimates of the needs for nature reserves in Sweden. Scand. J. For. Suppl. 2001, 3, 38–51. [Google Scholar] [CrossRef]
- Wade, C.M.; Austin, K.G.; Cajka, J.; Lapidus, D.; Everett, K.H.; Galperin, D.; Maynard, R.; Sobel, A. What is threatening forests in protected areas? A global assessment of deforestation in protected areas, 2001–2018. Forests 2020, 11, 539. [Google Scholar] [CrossRef]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Peres, C.A.; Bradshaw, C.J.; Laurance, W.F.; Lovejoy, T.E.; et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2011, 478, 378–381. [Google Scholar] [CrossRef]
- Roy, C.; Michel, N.L.; Handel, C.M.; Van Wilgenburg, S.L.; Burkhalter, J.C.; Gurney, K.E.B.; Messmer, D.J.; Princé, K.; Rushing, C.S.; Saracco, J.F.; et al. Monitoring boreal avian populations: How can we estimate trends and trajectories from noisy data? Avian Conserv. Ecol. 2019, 14, 8. [Google Scholar] [CrossRef] [Green Version]
- Lewandowski, P.; Przepióra, F.; Ciach, M. Single dead trees matter: Small-scale canopy gaps increase the species richness, diversity and abundance of birds breeding in a temperate deciduous forest. For. Ecol. Manag. 2021, 481, 118693. [Google Scholar] [CrossRef]
- Drolet, B.; Desrochers, A.; Fortin, M.-J. Effects of landscape structure on nesting songbird distribution in a harvested boreal forest. Condor 1999, 101, 699–704. [Google Scholar] [CrossRef]
- Virkkala, R.; Lehikoinen, A.; Rajasärkkä, A. Can protected areas buffer short-term population changes of resident bird species in a period of intensified forest harvesting? Biol. Conserv. 2020, 244, 108526. [Google Scholar] [CrossRef]
- Donázar, J.A.; Blanco, G.; Hiraldo, F.; Soto-Largo, E.; Oria, J. Effects of forestry and other land-use practices on the conservation of Cinerous Vultures. Ecol. Appl. 2002, 12, 1445–1456. [Google Scholar] [CrossRef]
- Askins, R.A. Open corridors in a heavily forested landscape: Impact on shrubland and forest-interior birds. Wildl. Soc. Bull. 1994, 22, 339–347. [Google Scholar]
- Hannon, S.J.; Schmiegelow, F. Corridors may not improve the conservation value of small reserves for most boreal birds. Ecol. Appl. 2002, 12, 1457–1468. [Google Scholar] [CrossRef]
- Kroodsma, R.L. Effect of edge on breeding forest bird species. Wilson Bull. 1984, 96, 426–436. [Google Scholar]
- Andrén, H.; Angelstam, P. Elevated predation rates as an edge effect in habitat islands. Ecology 1988, 69, 544–547. [Google Scholar] [CrossRef]
- Parker, T.H.; Stansberry, B.M.; Becker, C.D.; Gipson, P.S. Edge and area effects on the occurrence of migrant forest songbirds. Conserv. Biol. 2005, 19, 1157–1167. [Google Scholar] [CrossRef]
- LandOwner Resource Centre. Conserving the Forest Interior: A Threatened Wildlife Habitat. Extension Notes Ontario. Ontario Ministry of Natural Resources. 2000. Available online: http://www.lrconline.com/Extension_Notes_English/pdf/forInterior.pdf (accessed on 23 May 2023).
- Pfeifer, M.; Lefebvre, V.; Peres, C.; Banks-Leite, C.; Wearn, O.R.; Marsh, C.J.; Butchart, S.H.M.; Arroyo-Rodríguez, V.; Barlow, J.; Cerezo, A.; et al. Creation of forest edges has a global impact on forest vertebrates. Nature 2017, 551, 187–191. [Google Scholar] [CrossRef] [Green Version]
- McGarigal, K.; McComb, W.C. Relationships between landscape structure and breeding birds in the Oregon Coast Range. Ecol. Monogr. 1996, 65, 235–260. [Google Scholar] [CrossRef] [Green Version]
- Huhta, E.; Jokimäki, J.; Helle, P. Predation on artificial nests in a forest dominated landscape—The effects of nest type, patch size and edge structure. Ecography 1998, 21, 464–471. [Google Scholar] [CrossRef]
- Darveau, M.; Beauchesne, P.; Belanger, L.; Huot, J.; Larue, P. Riparian forest strips as habitat for breeding birds in boreal forest. J. Wildl. Manag. 1995, 59, 67–78. [Google Scholar] [CrossRef]
- Basile, M.; Storch, I.; Mikusiński, G. Abundance, species richness and diversity of forest bird assemblages—The relative importance of habitat structures and landscape context. Ecol. Indic. 2021, 133, 108402. [Google Scholar] [CrossRef]
- Villard, M.A.; Trzcinski, M.K.; Merriam, G. Fragmentation effects on forest birds: Relative influence of woodland cover and configuration on landscape occupancy. Conserv. Biol. 1999, 13, 774–783. [Google Scholar] [CrossRef] [Green Version]
- Mattsson, B.J. Louisiana Waterthrush Ecology and Conservation in the Georgia Piedmont. Ph.D. Thesis, University of Georgia, Athens, Georgia, 2006. Available online: https://www.researchgate.net/publication/242134898_LOUISIANA_WATERTHRUSH_ECOLOGY_AND_CONSERVATION_IN_THE_GEORGIA_PIEDMONT#fullTextFileContent (accessed on 17 May 2023).
- Ortega, Y.K.; Capen, D.E. Roads as edges: Effects on birds in forested landscapes. For. Sci. 2002, 48, 381–390. [Google Scholar]
- Przepióra, F.; Loch, J.; Ciach, M. Bark beetle infestation spots as biodiversity hotspots: Canopy gaps resulting from insect outbreaks enhance the species richness, diversity and abundance of birds breeding in coniferous forests. For. Ecol. Manag. 2020, 473, 118280. [Google Scholar] [CrossRef]
- Curtis, R. Some Simulation Estimates of Mean Annual Increment of Douglas Fir: Results, Limitations, and Implications for Management; PNW-RP-471; Pacific Northwest Research Station, USDA Forest Service: Olympia, WA, USA, 1994.
- Gruntorad, M.P.; Graham, K.A.; Arcilla, N.; Chizinski, C.J. Is hay for the birds? Investigating landowner willingness to time hay harvests for grassland bird conservation. Animals 2021, 11, 1030. [Google Scholar] [CrossRef]
- Björklund, H.; Valkama, J. Putting Forest Raptor Conservation into Practice: Guidelines for Forest Management and Voluntary Conservation; Finnish Museum of Natural History Luomus, University of Helsinki: Helsinki, Finland, 2017; Available online: https://tuhat.helsinki.fi/ws/portalfiles/portal/88678580/EOU_Turku2017_HeidiBj_rklund.pdf (accessed on 28 May 2023).
- Betts, M.G.; Harris, S.H. Selecting among land sparing, sharing and Triad in a temperate rainforest depends on biodiversity and timber production targets. J. Appl. Ecol. 2023, 60, 737–750. [Google Scholar] [CrossRef]
- Felton, A.; Löfroth, T.; Angelstam, P.; Gustafsson, L.; Hjältén, J.; Felton, A.M.; Simonsson, P.; Dahlberg, A.; Lindbladh, M.; Svensson, J.; et al. Keeping pace with forestry: Multi-scale conservation in a changing production forest matrix. AMBIO 2020, 49, 1050–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
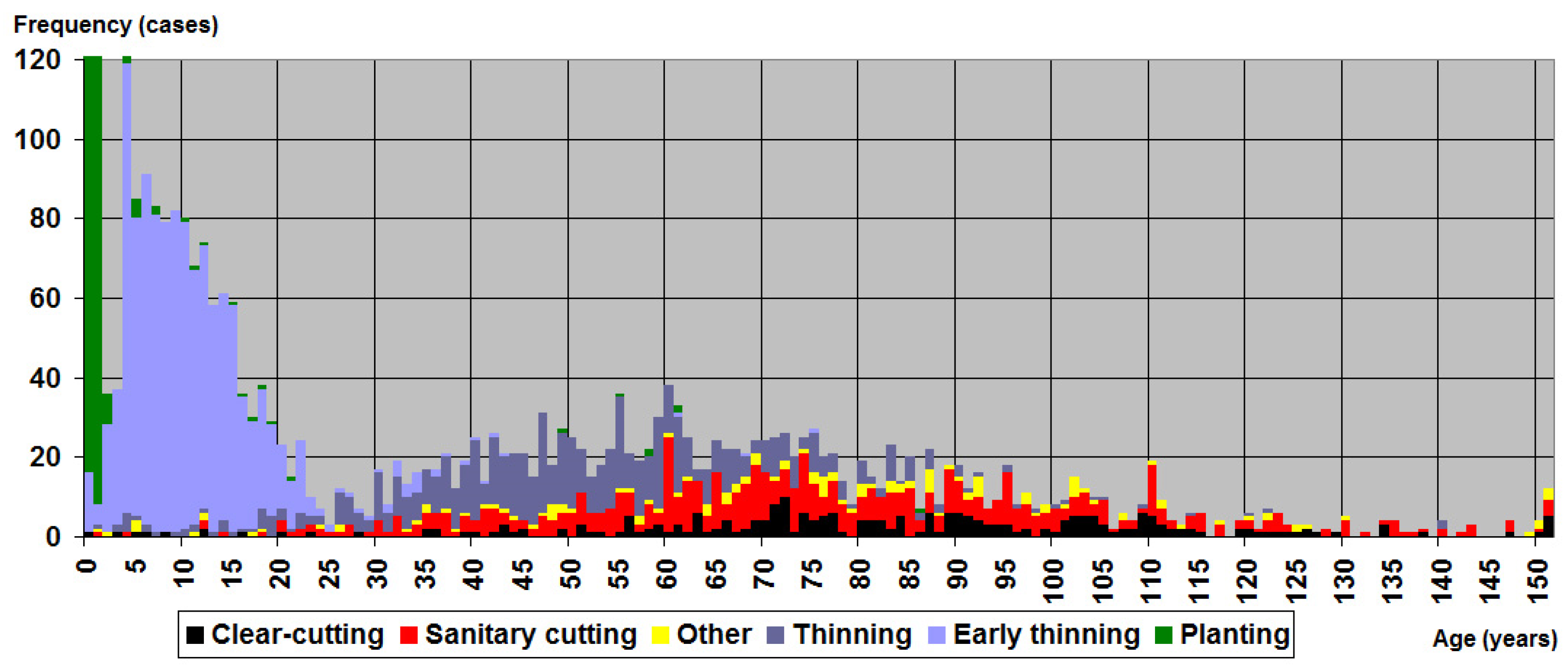{kind=link}
{kind=link}
{kind=link}
| Species | Average Age | Minimum Age | Maximum Age | Logging Age | Sample Size |
|---|---|---|---|---|---|
| Pine (Pinus sylvestris) | 206 | 81 | 430 | 101 | 455 |
| Oak (Quercus robur) | 169 | 135 | 190 | 101 | 30 |
| Aspen (Populus tremula) | 100 | 70 | 135 | 41 | 252 |
| Black Alder (Alnus glutinosa) | 120 | 85 | 159 | 71 | 26 |
| Spruce (Picea abies) | 157 | 119 | 210 | 81 | 17 |
| Principle | Practice | References |
|---|---|---|
| 1. Protect and enhance vertical structure | Employ uneven-aged silviculture | [8,9,23,24,28,29,33] |
| 2. Leave dying and dead trees and coarse woody debris | Retain snags, avoid crushing logs, and scatter tops and limbs | [7,8,21,81,82,107] |
| 3. Maintain residual large green trees | Protect mature trees and stands | [43,53,98,99,108,128] |
| 4. Integrate conservation areas into production forests | Establish and maintain uncut reserves and corridors | [62,87,103,104,109] |
| 5. Maximize forest interior and minimize fragmentation | Retain contiguous stands and reduce and “soften” edge areas | [6,7,8,74,102,112,119] |
| 6. Maintain buffers around riparian areas and nests | Protect riparian and nest sites | [6,7,8,79,120,123] |
| 7. Maintain horizontal stand structure | Establish small canopy caps | [14,25,57,94,107,125] |
| 8. Lengthen logging rotations | Increase cutting age of trees that have high value to birds | [11,40,49,79,108,125] |
| 9. Minimize post-logging and breeding season disturbance | Allow logging roads and trails to regenerate forest | [79,80,96,105,127,128] |
| 10. Manage for focal species and guilds | Identify priority species and quantitative objectives | [5,6,7,8,9,10,11,21,42,64,66,71] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arcilla, N.; Strazds, M. Ten Principles for Bird-Friendly Forestry: Conservation Approaches in Natural Forests Used for Timber Production. Birds 2023, 4, 245-261. https://doi.org/10.3390/birds4020021
Arcilla N, Strazds M. Ten Principles for Bird-Friendly Forestry: Conservation Approaches in Natural Forests Used for Timber Production. Birds. 2023; 4(2):245-261. https://doi.org/10.3390/birds4020021
Chicago/Turabian StyleArcilla, Nico, and Māris Strazds. 2023. "Ten Principles for Bird-Friendly Forestry: Conservation Approaches in Natural Forests Used for Timber Production" Birds 4, no. 2: 245-261. https://doi.org/10.3390/birds4020021