
Highly Conserved Microchromosomal Organization in Passeriformes Birds Revealed via BAC-FISH Analysis

 ,
,  , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species, Chromosome Preparation and Karyotype Description
2.2. Bacterial Artificial Chromosomes (BACs) FISH Experiments
3. Results
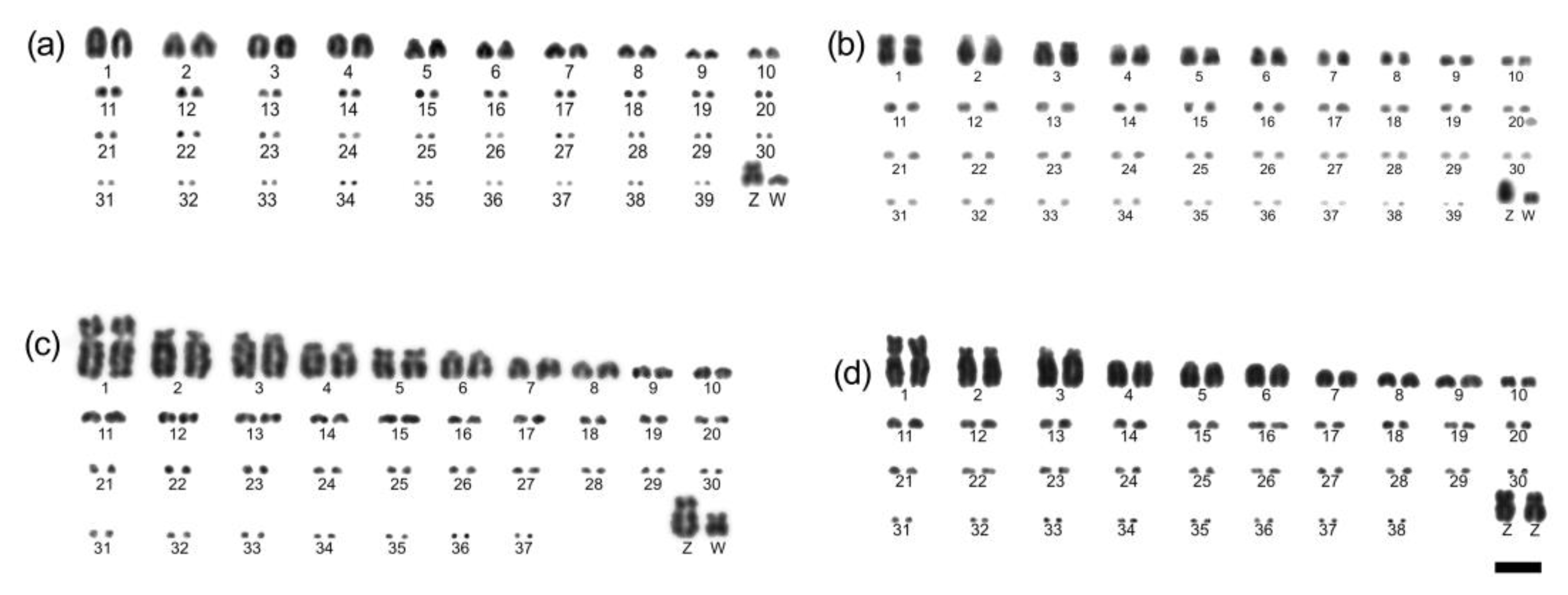
3.1. The Karyotype Description
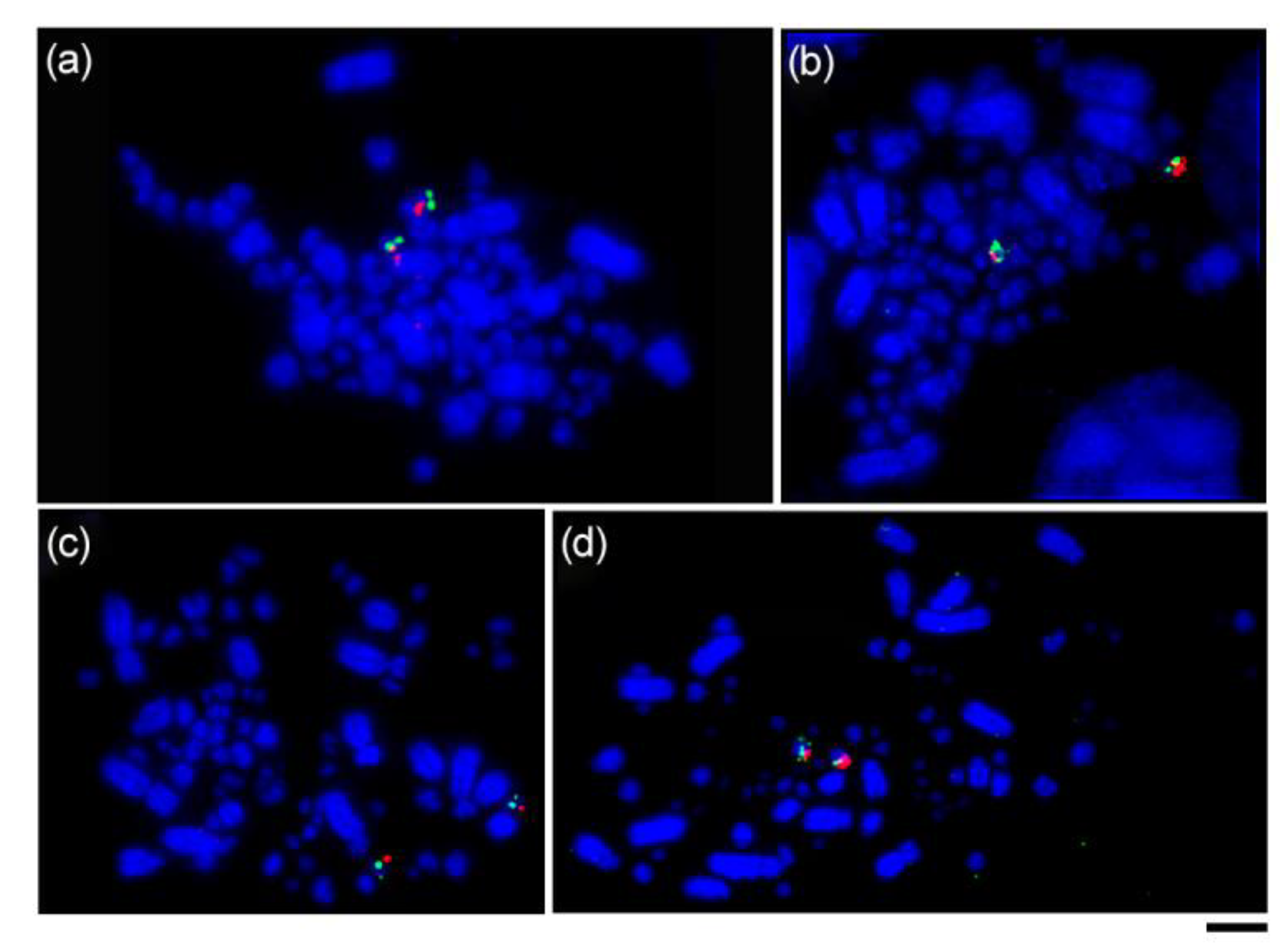
3.2. Bacterial Artificial Chromosomes Fluorescence In Situ Hybridization (BAC-FISH) Experiments
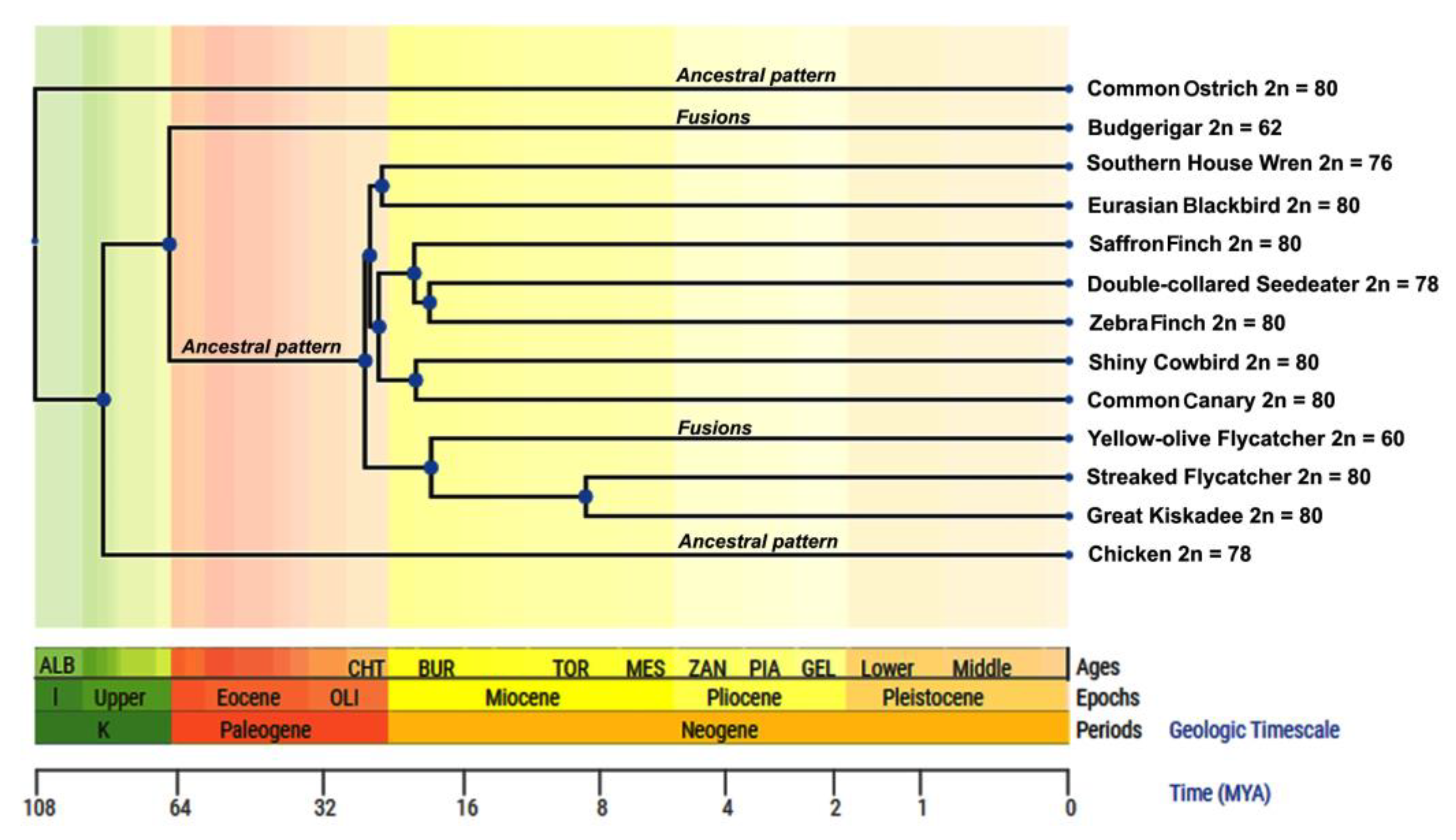
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sibley, C.G.; Monroe, B.L. Distribution and Taxonomy of Birds of the World; Yale University Press: New Haven, CT, USA, 1990. [Google Scholar]
- Cracraft, J.; Barker, F.K.; Hedges, S.B.; Kumar, S. Passerine birds (Passeriformes). In The Timetree of Life; Oxford University Press: New York, NY, USA, 2009; pp. 423–431. [Google Scholar]
- O’Connor, R.E.; Farré, M.; Joseph, S.; Damas, J.; Kiazim, L.; Jennings, R.; Bennett, S.; Slack, E.A.; Allanson, E.; Larkin, D.M.; et al. Chromosome-level assembly reveals extensive rearrangement in saker falcon and budgerigar, but not ostrich, genomes. Genome Biol. 2018, 19, 171. [Google Scholar] [CrossRef] [Green Version]
- Christidis, L. Chordata 3 B: Aves. In Animal Cytogenetics, 4th ed.; John, B., Ed.; Gebrüder Borntraeger: Berlin, Germany, 1990; Volume 4, 116p. [Google Scholar]
- Christidis, L. Extensive chromosomal repatterning in two congeneric species: Pytilia melba, L. and Pytilia phoenicoptera Swainson (Estrildidae; Aves). Cytogenet. Genome Res. 1983, 36, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Piccinni, E.; Stella, M. Some avian karyograms. Caryologia 1970, 23, 189–202. [Google Scholar]
- Bulatova, N.S.; Panov, E.N. Comparative analysis of karyotypes of 18 species family Turdidae (Aves). Caryologia 1973, 26, 229–244. [Google Scholar] [CrossRef]
- Bulatova, N.S. A comparative karyological study of passerine birds. Acta. Sci. Nat. Acad. Brno. 1981, 15, 1–44. [Google Scholar]
- Li, Q. Studies of bird karyotypes XI: Karyotypes in 15 Turdinae species. Hereditas 1989, 11, 17–20. [Google Scholar]
- Burt, D.W. Origin and evolution of avian microchromosomes. Cytogenet. Genome Res. 2002, 96, 97–112. [Google Scholar] [CrossRef]
- Nakatani, Y.; Takeda, H.; Kohara, Y.; Morishita, S. Reconstruction of the vertebrate ancestral genome reveals dynamic genome reorganization in early vertebrates. Genome Res. 2007, 17, 1254–1265. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.K.; Robertson, L.B.W.; Tempest, H.G.; Skinner, B.M. The evolution of the avian genome as revealed by comparative molecular cytogenetics. Cytogenet. Genome Res. 2007, 117, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Guttenbach, M.; Nanda, I.; Feichtinger, W.; Masabanda, J.S.; Griffin, D.K.; Schmid, M. Comparative chromosome painting of chicken autosomal paints 1–9 in nine different bird species. Cytogenet. Genome Res. 2003, 103, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Derjusheva, S.; Kurganova, A.; Haberman, F.; Gaginskaia, E. High chromosome conservation detected by comparative chromosome painting in chicken, pigeon and passerine birds. Chromosome Res. 2004, 12, 715–723. [Google Scholar] [CrossRef]
- Itoh, Y.; Arnold, A.P. Chromosomal polymorphism and comparative painting analysis in the zebra finch. Chromosome Res. 2005, 13, 47–56. [Google Scholar] [CrossRef]
- de Oliveira, E.H.C.; Tagliarini, M.M.; Nagamachi, C.Y.; Pieczarka, J.C. Genomic comparison in birds using chromosome-specific probes. Rev. Br. Ornit. 2006, 14, 47–52. [Google Scholar]
- Nanda, I.; Benisch, P.; Fetting, D.; Haaf, T.; Schmid, M. Synteny conservation of chicken macrochromosomes 1–10 in different avian lineages revealed by cross-species chromosome painting. Cytogenet. Genome Res. 2011, 132, 165–181. [Google Scholar] [CrossRef]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.V.; Furo, I.O.; O’Brien, P.C.M.; Fergunson-Smith, M.A.; de Oliveira, E.H.C. Molecular Cytogenetic Characterization of Multiple Intrachromosomal Rearrangements in Two Representatives of the Genus Turdus (Turdidae, Passeriformes). PLoS ONE 2014, 9, e103338. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; de Oliveira, E.H.C.; dos Santos, M.S.; Furo, I.O.; O’Brien, P.C.M.; Fergunson-Smith, M.A.; Garnero, A.V.; Gunski, R.J. Chromosome mapping of the large elaenia (Elaenia spectabilis): Evidence for a cytogenetic signature for passeriform birds? Biol. J. Linn. Soc. 2015, 115, 391–398. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, M.S.; Kretschmer, R.; Silva, F.A.O.; Ledesma, M.A.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Garnero, A.D.V.; de Oliveira, E.H.C.; Gunski, R.J. Intrachromosomal rearrangements in two representatives of the genus Saltator (Thraupidae, Passeriformes) and the occurrence of heteromorphic Z chromosomes. Genetica 2015, 143, 535–543. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, M.S.; Kretschmer, R.; Frankl-Vilches, C.; Bakker, A.; Gahr, M.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Comparative Cytogenetics between Two Important Songbird, Models: The Zebra Finch and the Canary. PLoS ONE 2017, 12, e0170997. [Google Scholar] [CrossRef] [Green Version]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.d.V.; de Freitas, T.R.O.; Toma, G.A.; Cioffi, M.d.B.; Oliveira, E.H.C.d.; O’Connor, R.E.; Griffin, D.K. Chromosomal Analysis in Crotophaga ani (Aves, Cuculiformes) Reveals Extensive Genomic Reorganization and an Unusual Z-Autosome Robertsonian Translocation. Cells 2021, 10, 4. [Google Scholar] [CrossRef]
- Kiazim, L.G.; O’Connor, R.E.; Larkin, D.M.; Romanov, M.N.; Narushin, V.G.; Brazhnik, E.A.; Griffin, D.K. Comparative Mapping of the Macrochromosomes of Eight Avian Species Provides Further Insight into Their Phylogenetic Relationships and Avian Karyotype Evolution. Cells 2021, 10, 362. [Google Scholar] [CrossRef]
- Furo, I.O.; Kretschmer, R.; dos Santos, M.S.; de Lima, C.A.C.; Gunski, R.J.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Cioffi, M.B.; de Oliveira, E.H.C. Chromosomal mapping of repetitive DNAs in Myiopsitta monachus and Amazona aestiva (Psittaciformes, Psittacidae) with emphasis on the sex chromosomes. Cytogenet. Genome Res. 2017, 151, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Barcellos, S.A.; de Souza, M.S.; Tura, V.; Pereira, L.R.; Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V. Direct Chromosome Preparation Method in Avian Embryos for Cytogenetic Studies: Quick, Easy and Cheap. DNA 2022, 2, 2. [Google Scholar] [CrossRef]
- Guerra, M.S. Reviewing the chromosome nomenclature of Levan et al. Rev. Bras. Genet. 1986, 9, 741–743. [Google Scholar]
- Griffith, S.C.; Ton, R.; Hurley, L.L.; McDiarmid, C.S.; Pacheco-Fuentes, H. The Ecology of the Zebra Finch Makes It a Great Laboratory Model but an Outlier amongst Passerine Birds. Birds 2021, 2, 60–76. [Google Scholar] [CrossRef]
- O’Connor, R.E.; Kiazim, L.; Skinner, B.; Fonseka, G.; Joseph, S.; Jennings, R.; Larkin, D.M.; Griffin, D.K. Patterns of microchromosome organization remain highly conserved throughout avian evolution. Chromosoma 2019, 128, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Damas, J.; O’connor, R.; Farré, M.; Lenis, V.P.E.; Martell, H.J.; Mandawala, A.; Fowler, K.; Joseph, S.; Swain, M.T.; Griffin, D.K.; et al. Upgrading short-read animal genome assemblies to chromosome level using comparative genomics and a universal probe set. Genome Res. 2017, 27, 875–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, M.S.; Barcellos, S.A.; dos Santos, M.d.S.; Gunski, R.J.; Garnero, A.d.V.; de Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K.; Kretschmer, R. Microchromosome BAC-FISH Reveals Different Patterns of Genome Organization in Three Charadriiformes Species. Animals 2022, 12, 3052. [Google Scholar] [CrossRef]
- Gunski, R.J.; Cabanne, G.S.; Ledesma, M.A.; Garnero, A.D.V. Análisis cariotípico de siete especies de Tiránidos (Tyrannidae). Hornero 2000, 15, 103–109. [Google Scholar] [CrossRef]
- Carvalho, M.V.P. Cytogenetic Studies in the Family Fringillidae (Passeriforms-Aves); Universidade Federal do Rio Grande do Sul: Porto Alegre, Brazil, 1989. [Google Scholar]
- De Lucca, E.J.; Waldrigues, A. Karyotypes of nine species of Passeriformes. Egypt. J. Genet. Cytol. 1985, 14, 41–50. [Google Scholar]
- Degrandi, T.M.; Barcellos, S.A.; Costa, A.L.; Garnero, A.D.; Hass, I.; Gunski, R.J. Introducing the bird chromosome database: An overview of cytogenetic studies in birds. Cytogenet. Genome Res. 2020, 160, 199–205. [Google Scholar] [CrossRef]
- Santos, L.P.; Gunski, R.J. Revisão de dados citogenéticos sobre a avifauna brasileira. Rev. Br. Ornit. 2006, 14, 35–45. [Google Scholar]
- Kretschmer, R.; Rodrigues, B.S.; Barcellos, S.A.; Costa, A.L.; Cioffi, M.d.B.; Garnero, A.d.V.; Gunski, R.J.; de Oliveira, E.H.C.; Griffin, D.K. Karyotype Evolution and Genomic Organization of Repetitive DNAs in the Saffron Finch, Sicalis flaveola (Passeriformes, Aves). Animals 2021, 11, 1456. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Franz, I.; de Souza, M.S.; Garnero, A.D.V.; Gunski, R.J.; de Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K.; de Freitas, T.R.O. Cytogenetic Evidence Clarifies the Phylogeny of the Family Rhynchocyclidae (Aves: Passeriformes). Cells 2021, 10, 2650. [Google Scholar] [CrossRef] [PubMed]
- Seroussi, E.; Knytl, M.; Pitel, F.; Elleder, D.; Krylov, V.; Leroux, S.; Morisson, M.; Yosefi, S.; Miyara, S.; Ganesan, S.; et al. Avian Expression Patterns and Genomic Mapping Implicate Leptin in Digestion and TNF in Immunity, Suggesting That Their Interacting Adipokine Role Has Been Acquired Only in Mammals. Int. J. Mol. Sci. 2019, 20, 4489. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Xu, Z.; Bai, H.; Huang, Y.; Kang, N.; Ding, X.; Liu, J.; Luo, H.; Yang, C.; Chen, W.; et al. Evolutionary analysis of a complete chicken genome. Proc. Natl. Acad. Sci. USA 2023, 120, e2216641120. [Google Scholar] [CrossRef]
- Kretschmer, R.; de Lima, V.L.C.; de Souza, M.S.; Costa, A.L.; O’Brien, P.C.; Ferguson-Smith, M.A.; Gunski, R.J.; Garnero, A.D.V. Multidirectional chromosome painting in Synallaxis frontalis (Passeriformes, Furnariidae) reveals high chromosomal reorganization, involving fissions and inversions. Comp. Cytogenet. 2018, 12, 97. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, B.S.; Kretschmer, R.; Gunski, R.J.; Garnero, A.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Chromosome Painting in Tyrant Flycatchers Confirms a Set of Inversions Shared by Oscines and Suboscines (Aves, Passeriformes). Cytogenet. Genome Res. 2017, 153, 205–212. [Google Scholar] [CrossRef]
- Romanov, M.N.; Farré-Belmonte, M.; Lithgow, P.E.; O’Connor, R.; Fowler, K.E.; Larkin, D.M.; Griffin, D.K. In Silico Reconstruction of Chromosomal Rearrangements and an Avian Ancestral Karyotype. In Proceedings of the International Plant and Animal Genome XXII Conference, San Diego, CA, USA, 11–16 January 2014. [Google Scholar]
- Olalla-Tárraga, M.A.; Amado, T.F.; Bini, L.M.; Martínez, P.A.; Morales-Castilla, I.; Torres-Romero, E.J.; Villalobos, F. Biological traits, phylogeny and human footprint signatures on the geographical range size of passerines (Order Passeriformes) worldwide. Glob. Ecol. Biogeogr. 2019, 28, 1183–1194. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Common Name | Scientific Name | Family | Suborder | N and Sex | Locality in Brazil |
|---|---|---|---|---|---|
| Streaked Flycatcher | Myiodynastes maculatus | Tyrannidae | Tyranni | 2 ♀ | Porto Vera Cruz-RS |
| Shiny Cowbird | Molothrus bonariensis | Icteridae | Passeri | 1 ♂ and 1 ♀ | São Gabriel-RS |
| Southern House Wren | Troglodytes aedon | Troglodytidae | Passeri | 1 ♂ and 2 ♀ | São Gabriel-RS |
| Double-collared Seedeater | Sporophila caerulescens | Thraupidae | Passeri | 2 ♂ | São Gabriel-RS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza, M.S.; Barcellos, S.A.; Tura, V.; Bobrowski, V.L.; Garnero, A.D.V.; Gunski, R.J.; Griffin, D.K.; Kretschmer, R. Highly Conserved Microchromosomal Organization in Passeriformes Birds Revealed via BAC-FISH Analysis. Birds 2023, 4, 236-244. https://doi.org/10.3390/birds4020020
de Souza MS, Barcellos SA, Tura V, Bobrowski VL, Garnero ADV, Gunski RJ, Griffin DK, Kretschmer R. Highly Conserved Microchromosomal Organization in Passeriformes Birds Revealed via BAC-FISH Analysis. Birds. 2023; 4(2):236-244. https://doi.org/10.3390/birds4020020
Chicago/Turabian Stylede Souza, Marcelo Santos, Suziane Alves Barcellos, Victoria Tura, Vera Lúcia Bobrowski, Analía Del Valle Garnero, Ricardo José Gunski, Darren K. Griffin, and Rafael Kretschmer. 2023. "Highly Conserved Microchromosomal Organization in Passeriformes Birds Revealed via BAC-FISH Analysis" Birds 4, no. 2: 236-244. https://doi.org/10.3390/birds4020020