
Susceptibility to Predation Varies with Body Mass, Foraging Niche, and Anti-Predator Responses among Bird Species

Independent Researcher, Montreal, QC, Canada
Birds 2023, 4(1), 73-84; https://doi.org/10.3390/birds4010006
Submission received: 12 January 2023
/
Revised: 1 February 2023
/
Accepted: 2 February 2023
/
Published: 5 February 2023
(This article belongs to the Special Issue Feature Papers of Birds 2022–2023)
Abstract
:Simple Summary
Predation can have a major impact on animal populations, and yet the factors that affect susceptibility to predation among species are poorly understood given that predation rate is difficult to measure in the field. Using estimates of predation rates from a large number of published radio-tracking studies in birds, I examined the general expectation that species facing fewer predators or those with effective anti-predator responses experience lower predation rates. I obtained 393 estimates of predation rates from 129 species, and the results generally supported the prediction. To obtain a fuller picture, more research is needed to broaden the range of species and habitats included in radio-tracking studies.
Abstract
Predation is a major source of mortality for many avian species. Species that face more predators, and those with less effective anti-predator responses, are presumably more likely to die from predation over time. Predation rate, as a measure of susceptibility to predation, is difficult to measure in the field. Radio-tracking studies, however, allow researchers to determine the time and cause of death of marked individuals, making it possible to estimate predation rate. I used estimates of predation rates from a large number of published radio-tracking studies in birds to assess in a phylogenetic framework the effect of several potential determinants. I obtained 393 estimates of predation rates from 129 species. Predation rates were lower in areas with fewer predators, such as islands and aquatic habitats, and for species with fewer potential predators, such as larger species. The predation rate was also lower for prey species with effective anti-predator responses, such as those that forage in flocks. Radio-tracking studies provide a unique opportunity to estimate overall predation rate in the field. Broadening the range of species and the range of habitats involved in such studies will help to further elucidate the factors that affect susceptibility to predation.
1. Introduction
Predation is a threat in the life of most animals. A recent review found that predation was responsible for about 55% of deaths in vertebrates [1], by far the leading cause of mortality from natural sources. Thus, the direct effect of predation can have a large impact on survival, population size, and viability [2]. Anti-predator responses by prey species to mitigate predation risk, such as changes in habitat use, vigilance, or group formation, often have large effects as well on animal populations [3,4]. The rate of predation experienced by the young and adults of a species can, in turn, constitute a major driver of life history traits such as fecundity and senescence [5]. A greater understanding of the factors that affect susceptibility to predation among species is, thus, important for ecology and evolution.
Not all species are expected to be equally susceptible to predation. The odds of dying from predation over a period of time represent a predation rate, which is a measure of susceptibility to predation. The predation rate for a prey species depends on inherent predation risk in the habitat and the effectiveness of anti-predator responses to reduce that risk [3]. Susceptibility to predation, therefore, is not a measure of predation risk, but rather the outcome of interactions between predators and prey after adjustments by prey to predation risk. For the same anti-predator responses, for instance, species living in habitats with a lower diversity and abundance of predators should be less susceptible to predation. As an example, islands and areas at higher latitudes typically harbour fewer predators [6,7], which would make prey species living there less susceptible to predation. By contrast, species that forage on the ground are often exposed to a greater diversity of predators than arboreal species or those that can fly [8,9]. Similarly, parental care during the breeding season can increase exposure to predators [10,11]. Thus, the predation rate should be higher for terrestrial species and during the breeding season, if those species cannot mitigate higher risk.
For the same level of predation risk, species that can deploy effective anti-predator responses should also be less susceptible to predation. A large body size, for example, can reduce the predation rate, as larger species can be harder to catch and subdue [2,12]. Living in groups represents another possible anti-predator response, as groups facilitate detection of approaching predators and dilute risk [13]. Living in groups would reduce predation rate, as long as groups are not attacked more often [14].
While the above predictions are straightforward, few tests have been carried out. One of the reasons is that documenting predation rate is challenging. Measuring predation rate requires a cohort of identifiable individuals that are followed over time, and the ability to determine the time and cause of death for each individual that dies. This is difficult to achieve in the field for mobile individuals, especially when predation events are rare. Estimates of predation rates are available for a number of primates, as researchers can follow identifiable groups long enough to document rare predation events [15]. For other mobile species such as birds, other approaches must be used. Susceptibility to predation in birds has been estimated by comparing the numbers of prey of different species brought back to the nest of avian predators, and their availability in the surrounding habitat [16]. This susceptibility index is a ratio that is independent of time, and measures predation for a particular prey by one predator species. Therefore, it is unsuitable to measure predation rate, especially for prey species with multiple predators. Another approach has measured the proportion of all deaths in a marked population caused by predation [1]. This is helpful to identify potential sources of mortality and their relative impact, but as the method yields a proportion independent of time, it cannot be used to estimate predation rate.
Here, I used published radio-tracking studies to estimate predation rates more directly in birds. With the help of small transmitters affixed to birds, researchers in a radio-tracking study can determine the locations of mobile individuals in their natural habitat at frequent intervals [17]. The locations of dead individuals can be determined rapidly, making it possible to ascertain whether predation was involved in each death. With this information, it is possible to calculate the susceptibility to predation as a predation rate, that is, the odds that an individual dies of predation over a set period of time.
In this study, I focused on predation by all sorts of predators, including snakes, birds, and mammals, on independent juvenile and adult birds. I examined the effect of potential ecological and behavioural determinants on predation rate. I tested the general expectation that the predation rate among species of birds should be lower in species with effective anti-predator responses, and in those facing fewer predators. In particular, I predicted that predation rates should be lower in larger species, in species dwelling on islands, in species occurring at higher latitudes, in ground-dwelling species, and in species that regularly form flocks to forage. The rationale for these and other predictions are presented in Table 1.
2. Materials and Methods
2.1. Data Collection
I searched the literature for radio-tracking studies in birds. I started with the most recent reviews of radio-tracking studies in birds [1,26], and updated the dataset up to August 2022 using searches on Web of Science and Google Scholar with the following key word combinations: (radio-telemetry or radio-tracking or radio-tagging) and (mortality or survival). From this set of studies, I selected those that provided information on the number of deaths from predation, as well as the number of days at risk of predation for the radio-marked individuals. I excluded studies based on captive-reared and/or translocated individuals, as observed predation rates might not be representative of the pattern of mortality in wild populations [34,35].
For each included study, I noted the age (juvenile vs. adult or both) of the radio-marked population, when the study took place (breeding season vs. non-breeding season or both), where the study took place (island or mainland, and spatial coordinates), and whether hunting occurred or not during the tracking period. The publication year was taken as a measure of time. I gathered the following additional information from the literature for each included species: adult body mass, diet, foraging substrate, foraging strata, and flocking tendency.
I used published sources for adult body mass, unless they were provided in the study [36,37]. I used a published standardised classification for diet, foraging substrate, and foraging strata [38]. The classification for diet included three categories based on the percentages of occurrence of different types of foods in the diet: carnivore (vertebrate or invertebrate prey and scavenging), herbivore (plant parts or seeds), and omnivore (a mixed category). Foraging substrate was either aquatic or terrestrial, and foraging strata was the percentage of time spent foraging on the ground. Using information from the literature [39], the tendency to form flocks during foraging was assessed during the tracking period. I made a distinction between species that form flocks on a regular basis (flocking) versus those that flock irregularly or not at all (non-flocking). Flocks include two or more individuals, and exclude family parties, mated pairs or cases where individuals congregate at fixed food sources such as fruiting trees or carrion.
2.2. Data Analysis
The first step involved calculating the predation rate in each included study. I considered two possible outcomes for each day at risk for a marked individual: alive or dead. Each day alive is a day at risk of dying from predation. The number of days at risk accrues from the beginning of the tracking period until the individual dies, when the study is terminated, or when the individual is censored. Censoring can occur for various reasons, including loss of transmitter, transmitter failure, or departure from the monitored area. The total number of days at risk was summed across all radio-marked individuals over the tracking period. The total number of deaths from predation over the tracking period was obtained from the papers, and excluded cases where the cause of death could not be determined. Nearly all studies excluded deaths occurring within a preset number of days after marking, in order to reduce the impact of capture-related stress. The number of days in the adaptation period was not standardised, but was typically between one and two weeks.
Viewed as a binomial process, the daily survival rate was calculated as follows: (total number of days at risk−number of deaths from predation)/(total number of days at risk) [40]. This daily survival rate was then expressed on a yearly basis, for ease of comparison, by raising it to the power of 365. Predation rate is the complement of this yearly survival rate, and is expressed as a proportion between 0 and 1. As most studies lasted less than a year and often focused on one season, predation rates from such studies expressed on a yearly basis should not be considered as estimates of annual mortality rates calculated using deaths from one full year. This is especially true when predation rates vary substantially with the time of year. When studies did not provide the total number of days at risk, it was necessary to use an approximation. For the purposes of testing the research hypotheses and as a first approach, I assumed that for a study of length t, an individual death or censorship occurred at t/2, which is reasonable for randomly distributed deaths or censorship.
For the statistical analysis of predation rate, I used a phylogenetic linear mixed model with the pglmm function from the phyr R package [41]. The dependent variable was the arc-sine square-root-transformed predation rate. This transformation, rather than the logit transformation which is often used for survival data [42], was deemed more appropriate, because predation rates of 0 could happen. The set of independent variables was described earlier. In this set, I included body mass in log10 scale, and calculated absolute latitudes of the study sites. Variance inflation factors among the independent variables were less than 2 for each variable, which meant that multicollinearity was not an issue in this model. Species was considered a random factor to account for multiple estimates for the same species. I also included whether days at risk of predation were approximated or provided by the authors as a random factor, in order to account for possible variation in the predation rate induced by methodology. To account for phylogenetic relatedness, I included the variance–covariance distance matrix between the included species. The distance matrix was obtained from a 50% majority consensus tree constructed from a set of 1000 phylogenetic trees [43]. I obtained the consensus tree and branch lengths from the ape R-package [44]. Note that it is not possible to weigh linear models with sample size in the phyr package.
3. Results
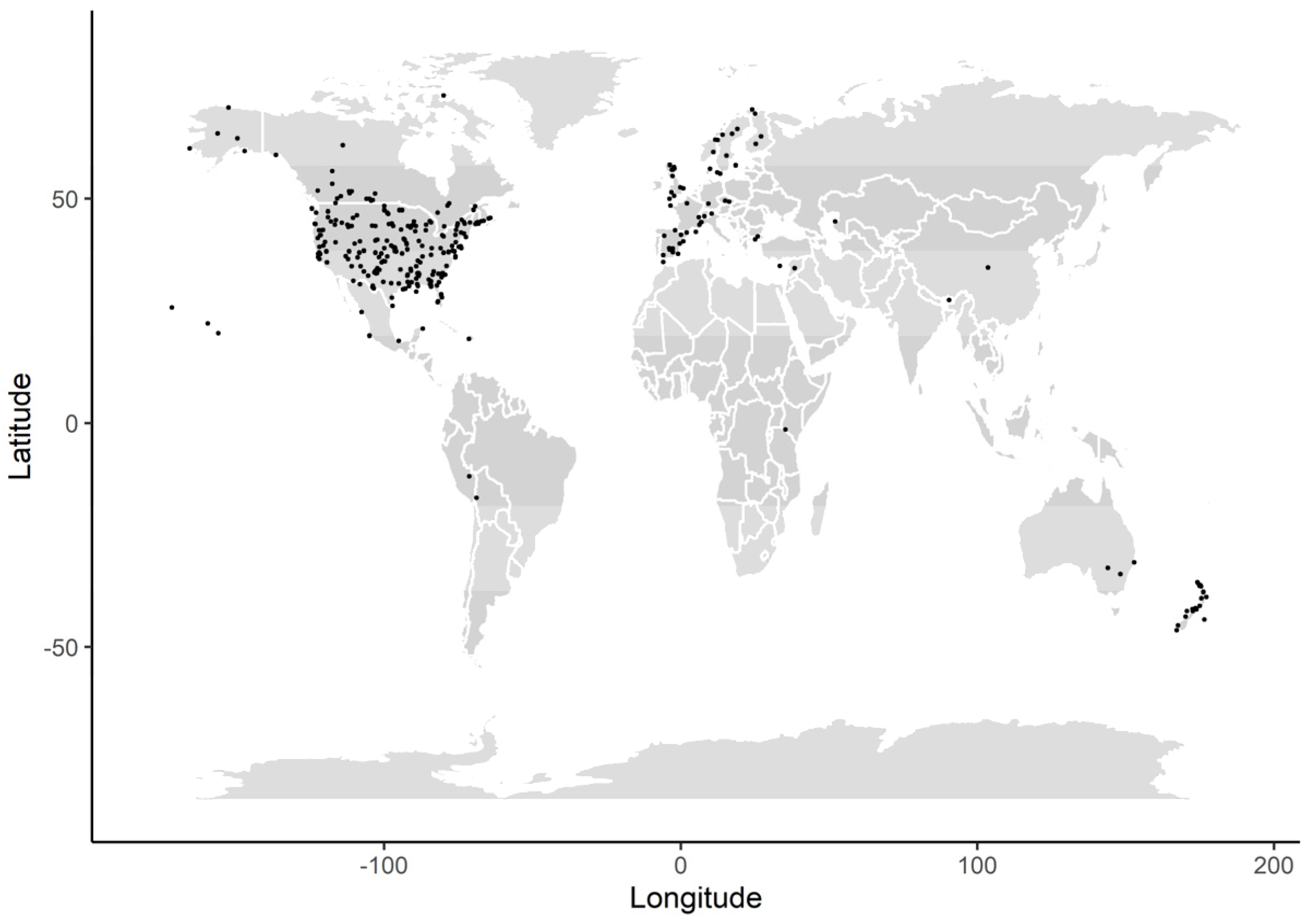
The dataset consisted of 393 estimates of predation rates from 129 species belonging to 38 families. The top five families included Phasianidae, Anatidae, Accipitridae, Odontophoridae, and Scolopacidae, which represented about 74% of the estimates. The top five studied species included Northern Bobwhite (Colinus virginianus), American Woodcock (Scolopax minor), Mallard (Anas platyrhynchos), Wild Turkey (Meleagris gallopavo), and Ruffed Grouse (Bonasa umbellus), which represented about 29% of the estimates. The body mass ranged from 8.9 to 9625 g, with a median across all estimates of about 650 g. The taxonomic coverage included many large harvested species. The coverage included more terrestrial than aquatic species (78% vs. 22%). and more species that forage close to the ground than above ground (80% vs. 20%). More estimates came from the northern than the southern hemisphere (92% vs. 8%), with North America and Europe figuring prominently in the dataset (Figure 1).
The number of days at risk of predation was approximated rather than provided by the researchers in about 58% of the cases. The breakdown of the data with respect to the other independent variables in the model was as follows: flocking present vs. absent (42% vs. 58%), carnivore vs. omnivore vs. herbivore (36% vs. 13% vs. 51%), island-dwelling vs. continent (7% vs. 93%), hunted vs. non-hunted (33% vs. 67%), breeding vs. non-breeding (26% vs. 74%), and juvenile vs. non-juvenile (20% vs. 80%).
The phylogenetic linear mixed model revealed that predation rates in birds decreased with adult body mass, but did not vary with absolute latitude (Table 2).
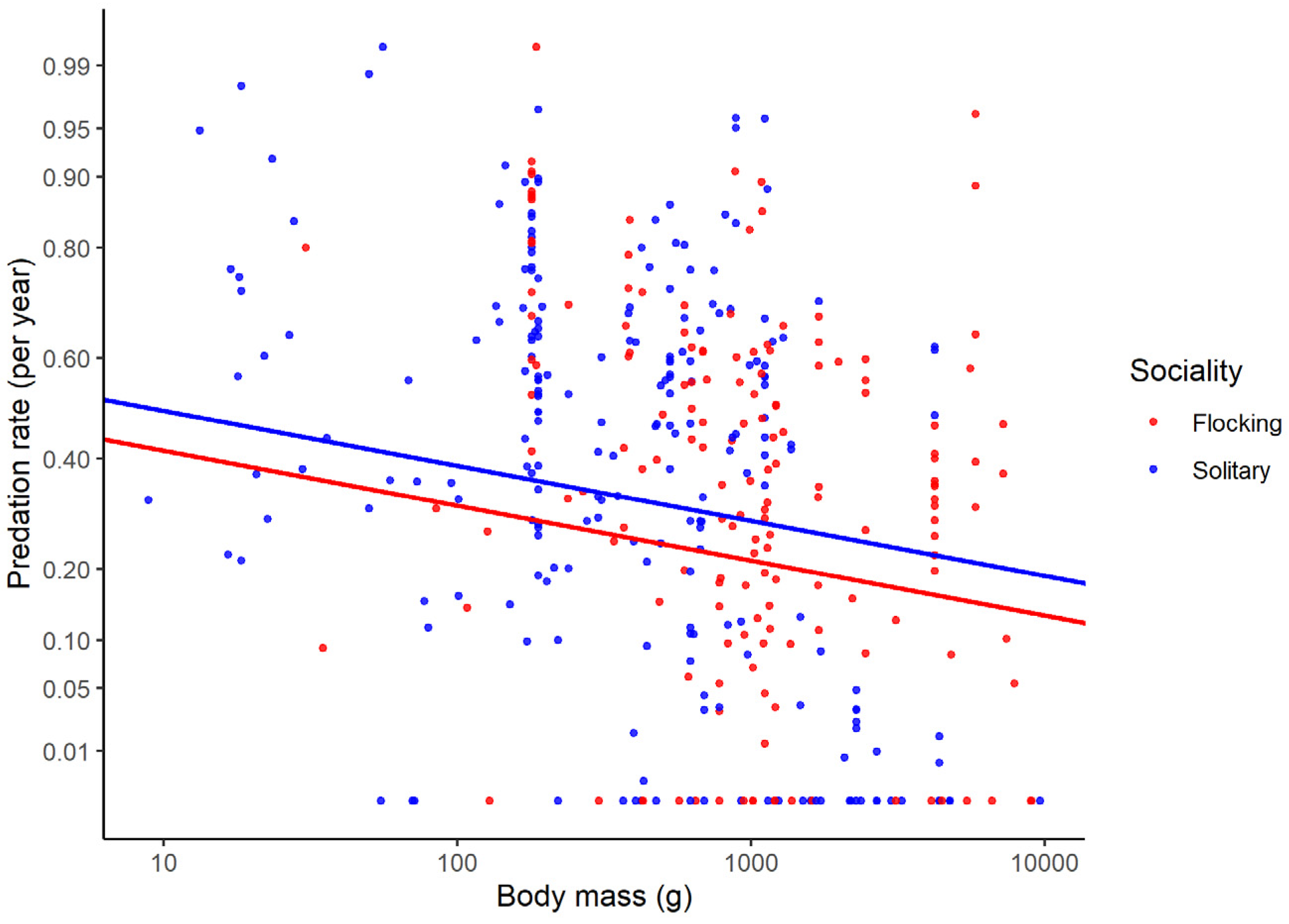
Controlling for these variables, predation rates were associated with several variables. The predation rate was lower in flocking than more solitary species (Figure 2).
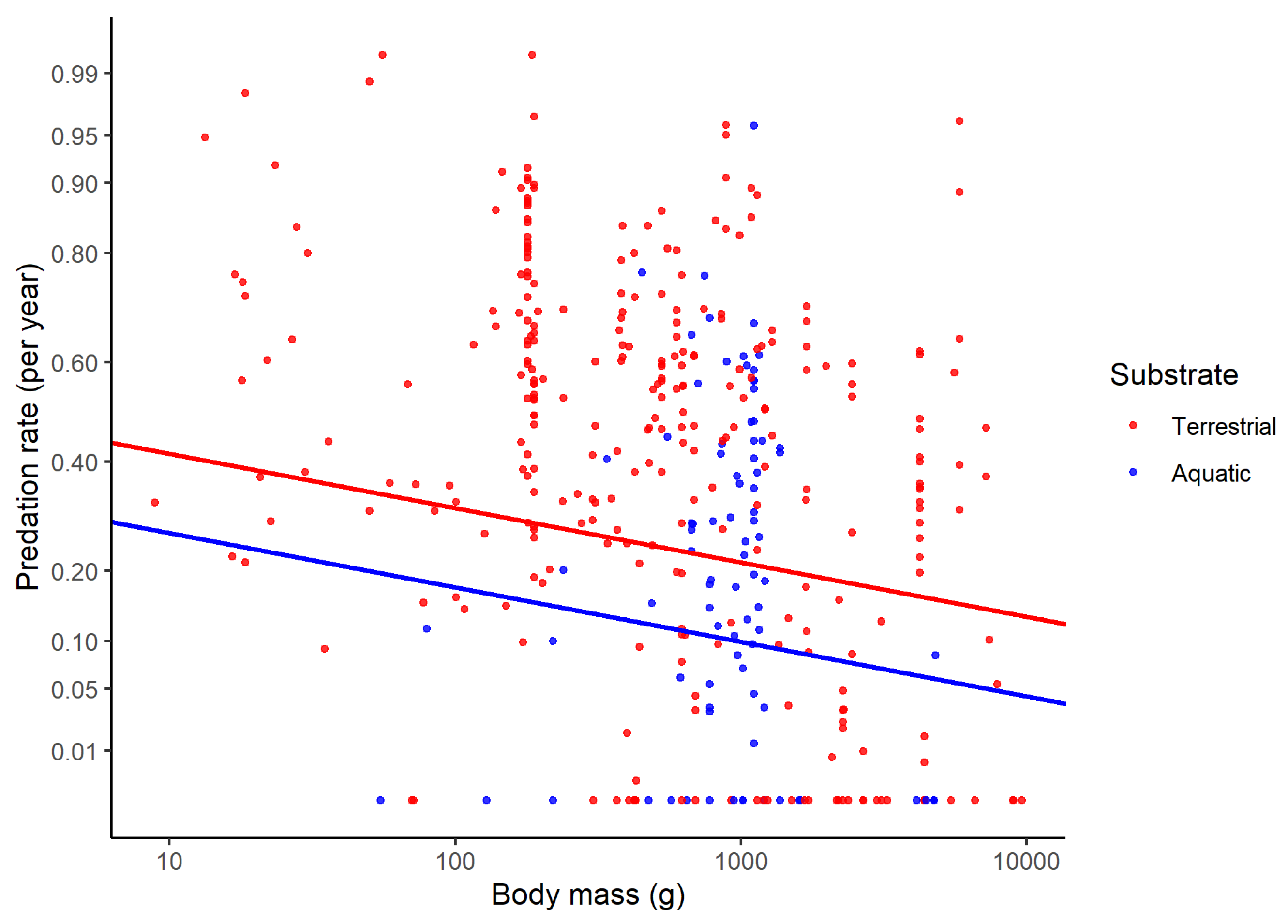
The predation rate was higher for terrestrial than aquatic species (Figure 3), and increased when species foraged closer to the ground. Diet was also important, as predation rates were lower in carnivores than in herbivores. The predation rate was lower for island-dwelling than continental species. The predation rate also tended to increase in populations exposed to hunting. The predation rate did not differ in the breeding and non-breeding seasons, but was higher in juveniles than in adults. Overall, predation rates were higher in earlier times. The methodology to calculate days at risk of predation accounted for little variation in predation rates (Table 2).
4. Discussion
I used published radio-tracking studies to estimate overall predation rates in birds. In a phylogenetic framework including a large number of species, the study revealed that susceptibility to predation, as measured by predation rate, is associated with several ecological and behavioural variables. In particular, body mass, flocking, and the foraging niche of a species were important correlates of predation rates in independent juvenile and adult birds.
The largest contributing factor to susceptibility to predation was adult body mass. In birds, predation rate decreased in larger species. The results support the prediction that species with a larger body mass are harder to catch and subdue [2,12,45,46]. A similar relationship was documented in a small number of primates [15]. Diet-based studies in birds have shown that prey preferences by avian predators decrease with prey body mass [16,47]. The proportion of total mortality attributable to predation also tends to decrease in larger species of birds and mammals [1,48]. The present study extends these findings to a larger set of species, using a measure of predation susceptibility based on time. The relationship between predation rate and body mass might not be linear across the whole range of body masses, as smaller species could be less profitable and avoided as well [16,23,49]. This is difficult to evaluate with radio-tracking studies, which typically focus on large species to minimize the impact of transmitters on survival and reproduction. More radio-tracking studies with smaller species are needed to evaluate the effect of adult body mass on predation rate over a larger spectrum for size.
Biodiversity of birds and mammals is known to increase at lower latitudes [50]. This is also true for the diversity and abundance of raptors, which are main predators of many bird species [7,18]. However, this latitudinal trend was not associated with predation rate. As noted earlier, few radio-tracking studies have been conducted in tropical areas. Further studies in such areas are needed to reach a firmer conclusion.
The lower predation rate in flocking species supports the prediction that flocking is associated with better predator detection and risk dilution [13,51]. Common Redshanks (Tringa totanus) in larger flocks, for instance, detect avian predators more quickly and are less susceptible to predation [52]. Annual adult survival often tends to be higher in flocking species [26,53,54]. Higher annual survival can thus be partly attributed to decreased predation. Predation rate is also a decreasing function for group size in primates [15,47]. These findings support the idea that living in groups represents an adaptation to higher predation risk [55,56,57,58].
The predation rate was higher in terrestrial than in aquatic species. Aquatic species probably face fewer predators overall, as water limits the ability of terrestrial predators to capture such prey. Similarly, nest predation is often lower in aquatic environments due to greater concealment in the marshy vegetation, or because the nests are less accessible [19,20]. Flocking tends to be more prevalent in aquatic species [59], and as flocking is associated with higher annual adult survival [26], lower predation rates in aquatic birds could arise simply through the indirect effect of flocking. However, the association between predation rate and foraging substrate was documented after controlling for the effect of flocking. In birds, aquatic species tend to live longer than terrestrial species [21] and show higher annual survival [22]. The lower predation rates documented here in aquatic birds could partly explain increased longevity and higher annual survival.
In addition to foraging substrate, the strata where species forage was also associated with predation rate. Indeed, predation rates decreased further away from the ground. Avian species foraging on the ground probably face a wider array of predators, including those that are strictly terrestrial. For instance, in addition to large birds of prey, predators of Wild Turkeys (Meleagris gallopavo), a ground-foraging species, include strictly terrestrial mammals such as coyotes (Canis latrans) and bobcats (Lynx rufus) [24]. By contrast, the main predators of small arboreal Passerines are only birds of prey [23]. In a guild of forest primates, predation rates were also higher in terrestrial than in arboreal species [47]. Increased longevity in non-volant arboreal species of birds and mammals [8] might reflect the reduction in predation rate away from the ground.
In terms of diet, herbivores were more susceptible to predation than carnivores. This result controls for flocking, which tends to be closely associated with diet in birds [58,59] and in mammals [55]. In birds, annual adult survival tends to be lower in herbivores than in carnivores [26], which fits with the higher predation rate for such species. Carnivores and omnivores, which capture moving prey, might have better visual detection abilities than herbivores. Thus, such animals would be able to react more quickly to approaching predators and experience lower predation rates [25]. However, in two recent large studies, diet was not associated with the distance at which prey species initiated escape [60,61]. In a study with a smaller sample of species, herbivores, in fact, tended to escape sooner from approaching threats [62]. Thus, why diet influences predation rates requires more study.
The predation rate was lower for island-dwelling species, supporting the idea that island living is associated with relaxed predation [63,64]. Island living is also associated with higher annual adult survival [65], reflecting in part the lower number and diversity of predators on islands [6]. The magnitude of the predation rate advantage was probably underestimated here, as island populations often face introduced predators that can have devastating impacts on survival [66].
The predation rate tended to be higher in harvested populations. Exposure to hunting is known to cause changes in prey behaviour or habitat use, in order to avoid hunters [28,29,30], which could lead to increased susceptibility to predation. Increased predation in harvested populations suggests a depensatory response by prey species to hunting mortality [67]. However, density-dependent processes such as predation could also explain why the predation rate is higher with hunting if hunted populations are larger [68]. Measures of prey abundance would be needed to address this issue. In fact, human activity associated with hunting could have the opposite effect on predation rate as it might displace large carnivores, thus providing a predation shield for the remaining prey [27]. The results suggest that across a large range of species, the shielding effect, if present, is not sufficient to overcome the negative impact of hunting.
The breeding season is often considered riskier in terms of predation [10]. Nevertheless, this higher risk was not associated with a higher predation rate in this study. It is possible that adaptations to higher risk, such as changes in territory location or behaviour [11], are effective in mitigating the higher risk during the breeding season. For instance, birds can avoid nesting close to the nests of avian predators [69]. Territorial species of birds during the breeding season can also benefit from heterospecific neighbours to improve predator detection [70].
The predation rate was higher in juveniles than adults. Annual survival estimates are typically lower in birds during their first year of life [71,72], reflecting in part their lack of efficiency in avoiding predators [31]. In American Black Ducks (Anas rubripes), for instance, hatch-year individuals were more likely to die from terrestrial predators such as racoon (Procyon lotor) and red fox (Vulpes fulva) than older individuals [73]. The effect of age was not very strong compared to other variables, such as diet or flocking. This is probably because this study focused on young birds after independence from parental care. Mortality typically decreases rapidly over time after independence [74,75], suggesting that a stronger effect of age would be found before independence. Future studies could concentrate on predation rates in birds, before or soon after fledging, to determine whether low survival during this period is also associated with increased predation rate.
Predation rates decreased over time. Over time, radio-tracking devices have become smaller and more powerful. The negative impact of these devices on survival might be less pronounced now than in the past, which could explain lower predation rates in more recent studies. Nevertheless, despite technological improvements, a meta-analysis found little changes in the magnitude of the negative effects of transmitters on survival over time [32]. Notice that the meta-analysis focused on survival, and not on mortality caused by predation. Alternatively, it is possible that contemporary radio-tracking studies with high predation rates are less likely to be published, thus indicating a form of publication bias. This is difficult to evaluate without access to unpublished studies. Another possible explanation is that a general decrease in predator populations over the years [33] has caused a decrease in predation rates.
This study has limitations. It is clear that radio-tracking studies focus on a non-random set of species. Indeed, large ground-dwelling species that are harvested are over-represented. Harvesting can have an impact on predation rate, as shown here; therefore, it is important to consider this potential bias in future studies. As radio-transmitters have become smaller over time, technological improvements might help increase data coverage for small arboreal species. The picture is also distorted by the geographical bias in the data, as radio-tracking studies in tropical areas are under-represented. Filling these gaps will help provide a fuller picture of the factors that influence predation rates in birds. This study mostly focused on natural habitats, but with urbanisation increasing rapidly, it might be interesting to compare predation rates in habitats with varying degrees of urbanisation. While anthropogenic mortality is typically higher in urban settings [1], urbanisation could reduce susceptibility to predation [76].
Attaching transmitters to birds can also have an impact on survival and predation rates. To reduce any short-term negative impacts of transmitters, researchers typically ignore mortality in the adaptation period following attachment. Nevertheless, transmitters can have a negative impact on survival in the long run, through factors such as extra weight, attachment type, and even possibly electromagnetic radiation [32,77]. Transmitters do not necessarily affect predation rate [78,79], but more evidence is needed. Individuals that are caught in radio-tracking studies might not represent a random sample of the population [80]. If the likelihood of capture is also linked to survival, this can introduce a bias in predation rate estimates. Assessing the cause of death is not an exact science, especially when carcasses are found many days after death. Errors in the assessment of cause of death will reduce the power to detect biological trends. It is not known how such errors vary along the ecological and behavioural dimensions explored here. Quicker access to carcasses after death, when possible, might reduce such errors. As a first approach, I relied on an approximation for the number of days at risk of predation when this information was not available. Fortunately, the methodology to calculate predation rate accounted for little variation in the results.
5. Conclusions
Despite their shortcomings, radio-tracking studies provide a unique opportunity to estimate overall predation rates in the field. Results from a large number of studies using a phylogenetic framework support the general prediction that species that face fewer predators or those with effective anti-predator responses experience a lower predation rate. Future studies based on more refined calculations in birds and in other taxa will shed more light on the ecological determinants of predation rate in animals.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The dataset is available upon request from the author.
Conflicts of Interest
The author declares no conflict of interest.
References
- Hill, J.E.; DeVault, T.L.; Belant, J.L. Cause-specific mortality of the world’s terrestrial vertebrates. Glob. Ecol. Biogeogr. 2019, 28, 680–689. [Google Scholar] [CrossRef]
- Sinclair, A.R.E.; Mduma, S.A.R.; Brashares, J.S. Patterns of predation in a diverse predator–prey system. Nature 2003, 425, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Creel, S.; Christianson, D. Relationships between direct predation and risk effects. Trends Ecol. Evol. 2008, 23, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D.A.; Bryga, H.; Endler, J.A. Experimentally induced life-history evolution in a natural population. Nature 1990, 346, 357–359. [Google Scholar] [CrossRef]
- Bliard, L.; Paquet, M.; Robert, A.; Dufour, P.; Renoult, J.P.; Grégoire, A.; Crochet, P.-A.; Covas, R.; Doutrelant, C. Examining the link between relaxed predation and bird coloration on islands. Biol. Lett. 2020, 16, 20200002. [Google Scholar] [CrossRef]
- Chapa-Vargas, L.; Ceballos, G.; Tinajero, R.; Torres-Romero, E.J. Latitudinal effects of anthropogenic factors driving raptor species richness across the American continent. J. Biogeogr. 2019, 46, 1948–1958. [Google Scholar] [CrossRef]
- Shattuck, M.R.; Williams, S.A. Arboreality has allowed for the evolution of increased longevity in mammals. Proc. Natl. Acad. Sci. USA 2010, 107, 4635–4639. [Google Scholar] [CrossRef]
- Healy, K.; Guillerme, T.; Finlay, S.; Kane, A.; Kelly, S.B.A.; McClean, D.; Kelly, D.J.; Donohue, I.; Jackson, A.L.; Cooper, N. Ecology and mode-of-life explain lifespan variation in birds and mammals. Proc. R. Soc. Lond B Biol. Sci. 2014, 281, 20140298. [Google Scholar] [CrossRef]
- Magnhagen, C. Predation risk as a cost of reproduction. Trends Ecol. Evol. 1991, 6, 183–186. [Google Scholar] [CrossRef]
- Lima, S.L. Predators and the breeding bird: Behavioral and reproductive flexibility under the risk of predation. Biol. Rev. 2009, 84, 485–513. [Google Scholar] [CrossRef]
- Cheney, D.L.; Wrangham, R.W. Predation. In Primate societies; Smuts, B.B., Cheney, D.L., Seyfarth, R.M., Wrangham, R.W., Eds.; University of Chicago Press: Chicago, IL, USA, 1987; pp. 227–239. [Google Scholar]
- Clark, C.W.; Mangel, M. The evolutionary advantages of group foraging. Theor. Popul. Biol. 1986, 30, 45–75. [Google Scholar] [CrossRef]
- Ioannou, C.C.; Bartumeus, F.; Krause, J.; Ruxton, G.D. Unified effects of aggregation reveal larger prey groups take longer to find. Proc. R. Soc. Lond B Biol. Sci. 2011, 278, 2985–2990. [Google Scholar] [CrossRef] [PubMed]
- Isbell, L.A. Predation on primates: Ecological patterns and evolutionary consequences. Evol. Anthropol. 1994, 3, 61–71. [Google Scholar] [CrossRef]
- Møller, A.P.; Nielsen, J.T. Malaria and risk of predation: A comparative study of birds. Ecology 2007, 88, 871–881. [Google Scholar] [CrossRef]
- Millspaugh, J.J.; Marzluff, J.M. Radio-tracking and animal populations: Past trends and future needs. In Radio Tracking and Animal Populations; Millspaugh, J.J., Marzluff, J.M., Eds.; Academic Press: Cambridge, MA, USA, 2001; pp. 383–393. [Google Scholar]
- Díaz, M.; Møller, A.P.; Flensted-Jensen, E.; Grim, T.; Ibáñez-Álamo, J.D.; Jokimäki, J.; Markó, G.; Tryjanowski, P. The geography of fear: A latitudinal gradient in anti-predator escape distances of birds across Europe. PLoS ONE 2013, 8, e64634. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.E. Avian life history evolution in relation to nest sites, nest predation, and food. Ecology 1995, 65, 101–127. [Google Scholar] [CrossRef]
- Rolland, C.; Danchin, E.; de Fraipont, M. The evolution of coloniality in birds in relation to food, habitat, predation, and life-history traits: A comparative analysis. Am. Nat. 1998, 151, 514–529. [Google Scholar] [CrossRef]
- Beauchamp, G. Group-foraging is not associated with longevity in North American birds. Biol. Lett. 2010, 6, 42–44. [Google Scholar] [CrossRef]
- Bird, J.P.; Martin, R.; Akçakaya, H.R.; Gilroy, J.; Burfield, I.J.; Garnett, S.T.; Symes, A.; Taylor, J.; Şekercioğlu, Ç.H.; Butchart, S.H.M. Generation lengths of the world’s birds and their implications for extinction risk. Conserv. Biol. 2020, 34, 1252–1261. [Google Scholar] [CrossRef]
- Götmark, F.; Post, P. Prey selection by sparrowhawks, Accipiter nisus: Relative predation risk for breeding passerine birds in relation to their size, ecology and behaviour. Phil. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1559–1577. [Google Scholar]
- Hughes, T.W.; Tapley, J.L.; Kennamer, J.E.; Lehman, C.P. The imapct of predation on wild turkeys. Proc. Natl. Wild Turk. Symp. 1997, 9, 117–126. [Google Scholar]
- Blumstein, D.T. Developing an evolutionary ecology of fear: How life history and natural history traits affect disturbance tolerance in birds. Anim. Behav. 2006, 71, 389–399. [Google Scholar] [CrossRef]
- Beauchamp, G. Flocking in birds increases annual adult survival in a global analysis. Oecologia 2021, 197, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Leighton, P.A.; Horrocks, J.A.; Kramer, D.L. Conservation and the scarecrow effect: Can human activity benefit threatened species by displacing predators? . Biol. Conserv. 2010, 143, 2156–2163. [Google Scholar] [CrossRef]
- Ciuti, S.; Northrup, J.M.; Muhly, T.B.; Simi, S.; Musiani, M.; Pitt, J.A.; Boyce, M.S. Effects of humans on behaviour of wildlife exceed those of natural predators in a landscape of fear. PLoS ONE 2012, 7, e50611. [Google Scholar] [CrossRef]
- Crosmary, W.G.; Makumbe, P.; Côté, S.D.; Fritz, H. Vulnerability to predation and water constraints limit behavioural adjustments of ungulates in response to hunting risk. Anim. Behav. 2012, 83, 1367–1376. [Google Scholar] [CrossRef]
- Suraci, J.P.; Clinchy, M.; Zanette, L.Y.; Wilmers, C.C. Fear of humans as apex predators has landscape-scale impacts from mountain lions to mice. Ecol. Lett. 2019, 22, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, K.A. Predation and starvation: Age-specific mortality in juvenile juncos (Junco phaenotus). J. Anim. Ecol. 1989, 58, 275–286. [Google Scholar] [CrossRef]
- Barron, D.G.; Brawn, J.D.; Weatherhead, P.J. Meta-analysis of transmitter effects on avian behaviour and ecology. Methods Ecol. Evol. 2010, 1, 180–187. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.S.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic downgrading of planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Meriggi, A.; Brangi, A.; Cuccus, P.; Della Stella, R.M. High mortality rate in a re-introduced grey partridge population in central Italy. Ital. J. Zool. 2002, 69, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Leighton, K.; Chilvers, D.; Charles, A.; Kelly, A. Post-release survival of hand-reared tawny owls (Strix aluco) based on radio-tracking and leg-band return data. Anim. Welf. 2008, 17, 207–214. [Google Scholar] [CrossRef]
- Lislevand, T.; Figuerola, J.; Székely, T. Avian body sizes in relation to fecundity, mating system, display behaviour, and resource sharing. Ecology 2007, 88, 1605. [Google Scholar] [CrossRef]
- Dunning, J.B. CRC Handbook of Avian Body Masses, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Wilman, H.; Belmaker, J.; Simpson, J.; de la Rosa, C.; Rivadeneira, M.M.; Jetz, W. EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 2014, 95, 2027. [Google Scholar]
- Billerman, S.M.; Keeney, B.K.; Rodewald, P.G.; Schulenberg, T.S. (Eds.) Birds of the World; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2022. [Google Scholar]
- Heisey, D.M.; Fuller, T.K. Evaluation of survival and cause-specific mortality rates using telemetry data. J. Wildl. Manage. 1985, 49, 668–674. [Google Scholar] [CrossRef]
- Li, D.; Dinnage, R.; Nell, L.A.; Helmus, M.R.; Ives, A.R. Phyr: An R package for phylogenetic species-distribution modelling in ecological communities. Methods Ecol. Evol. 2020, 11, 1455–1463. [Google Scholar] [CrossRef]
- Scholer, M.N.; Strimas-Mackey, M.; Jankowski, J.E. A meta-analysis of global avian survival across species and latitude. Ecol. Lett. 2020, 23, 1537–1549. [Google Scholar] [CrossRef]
- Hackett, S.J.; Kimball, R.T.; Reddy, S.; Bowie, R.C.K.; Braun, E.L.; Braun, M.J.; Chojnowski, J.L.; Cox, W.A.; Han, K.L.; Harshman, J.; et al. A phylogenomic study of birds reveals their evolutionary history. Science 2008, 320, 1763–1768. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- Cohen, J.E.; Pimm, S.L.; Yodzis, P.; Sadana, J. Body sizes of animal predators and animal prey in food webs. J. Anim. Ecol. 1993, 62, 67–78. [Google Scholar] [CrossRef]
- Thiollay, J.-M.; Jullien, M. Flocking behaviour of foraging birds in a neotropical rain forest and the antipredator defence hypothesis. Ibis 1998, 140, 382–394. [Google Scholar] [CrossRef]
- Shultz, S.; Noë, R.; McGraw, W.S.; Dunbar, R.I.M. A community-level evaluation of the impact of prey behavioural and ecological characteristics on predator diet composition. Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, 725–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, C.; Kays, R. Causes of mortality in North American populations of large and medium-sized mammals. Anim. Conserv 2011, 14, 474–483. [Google Scholar] [CrossRef]
- Hayward, M.W.; Henschel, P.; O’Brien, J.; Hofmeyr, M.; Balme, G.; Kerley, G.I.H. Prey preferences of the leopard (Panthera pardus). J. Zool. 2006, 270, 298–313. [Google Scholar] [CrossRef]
- Kinlock, N.L.; Prowant, L.; Herstoff, E.M.; Foley, C.M.; Akin-Fajiye, M.; Bender, N.; Umarani, M.; Ryu, H.-Y.; Şen, B.; Gurevitch, J. Explaining global variation in the latitudinal diversity gradient: Meta-analysis confirms known patterns and uncovers new ones. Glob. Ecol. Biogeogr. 2018, 27, 125–141. [Google Scholar] [CrossRef]
- Caraco, T.; Martindale, S.; Pulliam, H.R. Avian flocking in the presence of a predator. Nature 1980, 285, 400–401. [Google Scholar] [CrossRef]
- Cresswell, W. Flocking is an effective anti-predation strategy in redshanks, Tringa totanus. Anim. Behav. 1994, 47, 433–442. [Google Scholar] [CrossRef]
- Jullien, M.; Clobert, J. The survival value of flocking in neotropical birds: Reality or fiction? Ecology 2000, 81, 3416–3430. [Google Scholar] [CrossRef]
- Srinivasan, U. Morphological and behavioral correlates of long-term bird survival in selectively logged forest. Front. Ecol. Evol. 2019, 7, 17. [Google Scholar] [CrossRef]
- Brashares, J.S.; Garland, T.; Arcese, P. Phylogenetic analysis of coadaptation in behavior, diet, and body size in the African antelope. Behav. Ecol. 2000, 11, 452–463. [Google Scholar] [CrossRef]
- Ebensperger, L.A.; Bozinovic, F. Communal burrowing in the hystricognath rodent, Octogon degus: A benefit of sociality? Behav. Ecol. Sociobiol. 2000, 47, 365–369. [Google Scholar] [CrossRef]
- Stankowich, T.; Haverkamp, P.J.; Caro, T. Ecological drivers of antipredator defenses in Carnivores. Evolution 2014, 68, 1415–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Liker, A.; Liu, Y.; Székely, T. Evolution of social organization: Phylogenetic analyses of ecology and sexual selection in Weavers. Am. Nat. 2022, 200, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, G. Flocking in birds is associated with diet, foraging substrate, timing of activity, and life history. Behav. Ecol. Sociobiol. 2022, 76, 74. [Google Scholar] [CrossRef]
- Weston, M.A.; Radkovic, A.; Kirao, L.; Guay, P.-J.; Van Dongen, W.F.D.; Malaki, P.; Blumstein, D.T.; Symonds, M.R.E. Differences in flight initiation distances between African and Australian birds. Anim. Behav. 2021, 179, 235–245. [Google Scholar] [CrossRef]
- Ekanayake, K.B.; Gnanapragasam, J.J.; Ranawana, K.; Vidanapathirana, D.R.; Abeyawardhana, U.T.; Fernando, C.H.; McQueen, A.; Weston, M.A.; Symonds, M.R.E. Ecological and environmental predictors of escape among birds on a large tropical island. Behav. Ecol. Sociobiol. 2022, 76, 31. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Díaz, M.; Grim, T.; Ibáñez-Álamo, J.D.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Tätte, K.; Markó, G.; Jiang, Y.; et al. Contagious fear: Escape behavior increases with flock size in European gregarious birds. Ecol. Evol. 2019, 9, 6096–6104. [Google Scholar] [CrossRef]
- Willis, E.O. Do birds flock in Hawaii, a land without predators? Calif. Birds 1972, 3, 1–9. [Google Scholar]
- Blumstein, D.T.; Daniel, J.C. The loss of anti-predator behaviour following isolation on islands. Proc. R. Soc. Lond. B Biol. Sci. 2005, 272, 1663–1668. [Google Scholar] [CrossRef]
- Beauchamp, G. Do avian species survive better on islands? Biol. Lett. 2021, 16, 20200643. [Google Scholar] [CrossRef] [PubMed]
- Courchamp, F.; Chapuis, J.-L.; Pascal, M. Mammal invaders on islands: Impact, control and control impact. Biol. Rev. 2003, 78, 347–383. [Google Scholar] [CrossRef] [PubMed]
- Péron, G. Compensation and additivity of anthropogenic mortality: Life-history effects and review of methods. J. Anim. Ecol. 2013, 82, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Riecke, T.V.; Lohman, M.G.; Sedinger, B.S.; Arnold, T.W.; Feldheim, C.L.; Koons, D.N.; Rohwer, F.C.; Schaub, M.; Williams, P.J.; Sedinger, J.S. Density-dependence produces spurious relationships among demographic parameters in a harvested species. J. Anim. Ecol. 2022, 91, 2261–2272. [Google Scholar] [CrossRef] [PubMed]
- Suhonen, J.; Norrdahl, K.; Korpimaki, E. Avian predation risk modifies breeding bird community on a farmland area. Ecology 1994, 75, 1626–1634. [Google Scholar] [CrossRef]
- Groom, M.J. Sand-colored nighthawks parasitize the antipredator behavior of three nesting bird species. Ecology 1992, 73, 785–793. [Google Scholar] [CrossRef]
- Sæther, B.-E. Survival rates in relation to body weight in European birds. Ornis Scand. 1989, 20, 13–21. [Google Scholar] [CrossRef]
- Muñoz, A.P.; Kéry, M.; Martins, P.V.; Ferraz, G. Age effects on survival of Amazon forest birds and the latitudinal gradient in bird survival. Auk 2018, 135, 299–313. [Google Scholar] [CrossRef]
- Conroy, M.J.; Costanzo, G.R.; Stotts, D.B. Winter survival of female American Black Ducks on the Atlantic Coast. J. Wildl. Manage. 1989, 53, 99–109. [Google Scholar] [CrossRef]
- Maness, T.J.; Anderson, D.J. Predictors of juvenile survival in birds. Ornithol. Monogr. 2013, 78, 1–55. [Google Scholar] [CrossRef]
- Naef-Daenzer, B.; Grüebler, M.U. Post-fledging survival of altricial birds: Ecological determinants and adaptation. J. Field Ornithol. 2016, 87, 227–250. [Google Scholar] [CrossRef]
- Møller, A.P. Flight distance of urban birds, predation, and selection for urban life. Behav. Ecol. Sociobiol. 2008, 63, 63–75. [Google Scholar] [CrossRef]
- Balmori, A. Radiotelemetry and wildlife: Highlighting a gap in the knowledge on radiofrequency radiation effects. Sci. Tot. Environ. 2016, 543, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Thirgood, S.J.; Redpath, S.M.; Hudson, P.J.; Hurley, M.M.; Aebischer, N.J. Effects of necklace radio transmitters on survival and breeding success of red grouse Lagopus lagopus scoticus. Wildl. Biol. 1995, 1, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Naef-Daenzer, B.; Widmer, F.; Nuber, M. A test for effects of radio-tagging on survival and movements of small birds. Avian Sci. 2001, 1, 15–23. [Google Scholar]
- Biro, P.A.; Dingemanse, N.J. Sampling bias resulting from animal personality. Trends Ecol. Evol. 2009, 24, 66–67. [Google Scholar] [CrossRef]
Figure 1.
Geographical locations of study sites where predation rates were obtained for different bird species. Locations are shown as black dots. Some locations include more than one predation rate estimate.
Figure 1.
Geographical locations of study sites where predation rates were obtained for different bird species. Locations are shown as black dots. Some locations include more than one predation rate estimate.

Figure 2.
Predation rates expressed on a yearly basis as a function of adult body mass for solitary and flocking species of birds. The regression lines were obtained from a phylogenetic linear mixed model. Predation rate is shown in arc-sine square-root scale, and body mass in log10 scale.
Figure 2.
Predation rates expressed on a yearly basis as a function of adult body mass for solitary and flocking species of birds. The regression lines were obtained from a phylogenetic linear mixed model. Predation rate is shown in arc-sine square-root scale, and body mass in log10 scale.

Figure 3.
Predation rate expressed on a yearly basis as a function of adult body mass for terrestrial and aquatic species of birds. The regression lines were obtained from a phylogenetic linear mixed model. Predation rate is shown in arc-sine square-root scale, and body mass in log10 scale.
Figure 3.
Predation rate expressed on a yearly basis as a function of adult body mass for terrestrial and aquatic species of birds. The regression lines were obtained from a phylogenetic linear mixed model. Predation rate is shown in arc-sine square-root scale, and body mass in log10 scale.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Predictions and rationale for variables included as predictors of predation rate in birds.
| Variable | Prediction | Rationale | Source |
|---|---|---|---|
| Body mass | Predation rate decreases with body mass | Large body mass makes prey more difficult to capture and subdue | [2,12] |
| Latitude | Predation rate increases at lower latitudes | Latitudinal gradient in predator diversity and abundance | [7,18] |
| Flocking | Predation rate decreases in flocking species | Living in groups increases the ability to detect predators and dilutes risk | [13] |
| Foraging substrate | Predation rate is lower for aquatic than terrestrial species | Fewer predators in aquatic habitats | [19,20,21,22] |
| Foraging strata | Predation rate increases closer to the ground | Greater diversity of predators closer to the ground | [23,24] |
| Diet | Predation rate is higher for herbivores than carnivores | Better visual detection abilities in carnivores | [25,26] |
| Island living | Predation rate decreases in island-living species | Lower diversity and abundance of predators on islands | [6] |
| Hunting | Predation rate varies for harvested species | Changes in habitat and prey behaviour increases predation risk but hunting might shield prey species from predators | [27,28,29,30] |
| Season | Predation rate is higher in the breeding season | Parental care increases encounters with predators | [11] |
| Age | Predation rate is higher in juveniles than in adults | Relative lack of experience of juveniles with predators | [31] |
| Time | Predation rate decreases over time | Technological improvements over time reduce the negative impact of transmitters or general decrease in predator populations over the years | [32,33] |
Table 2.
Estimates in arc-sine square-root scale from a phylogenetic linear mixed model of various potential correlates of predation rates in birds.
Table 2.
Estimates in arc-sine square-root scale from a phylogenetic linear mixed model of various potential correlates of predation rates in birds.
| Variable | β (SE) | p |
|---|---|---|
| Fixed effects | ||
| Body mass in log10 scale | −0.34 (0.040) | <0.0001 |
| Absolute latitude | 0.0013 (0.0017) | 0.44 |
| Solitary vs. flocking | 0.12 (0.047) | 0.013 |
| Aquatic vs. terrestrial | −0.18 (0.062) | 0.003 |
| Foraging strata | 0.0029 (0.00092) | 0.001 |
| Herbivore vs. carnivore | 0.31 (0.060) | <0.0001 |
| Omnivore vs. carnivore | 0.098 (0.075) | 0.19 |
| Island vs. continent | −0.16 (0.079) | 0.047 |
| Hunting vs. non-hunting | 0.067 (0.035) | 0.058 |
| Breeding vs. non-breeding | 0.053 (0.047) | 0.26 |
| Juvenile vs. non-juvenile | 0.077 (0.036) | 0.03 |
| Time | −0.0042 (0.0014) | 0.002 |
| Random effects | Variance | |
| Species | 0.040 | |
| Methodology | 0.000000076 | |
| Residual variance | 0.051 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Beauchamp, G. Susceptibility to Predation Varies with Body Mass, Foraging Niche, and Anti-Predator Responses among Bird Species. Birds 2023, 4, 73-84. https://doi.org/10.3390/birds4010006
AMA Style
Beauchamp G. Susceptibility to Predation Varies with Body Mass, Foraging Niche, and Anti-Predator Responses among Bird Species. Birds. 2023; 4(1):73-84. https://doi.org/10.3390/birds4010006
Chicago/Turabian StyleBeauchamp, Guy. 2023. "Susceptibility to Predation Varies with Body Mass, Foraging Niche, and Anti-Predator Responses among Bird Species" Birds 4, no. 1: 73-84. https://doi.org/10.3390/birds4010006