
Flight Type and Seasonal Movements Are Important Predictors for Avian Collisions in Wind Farms

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
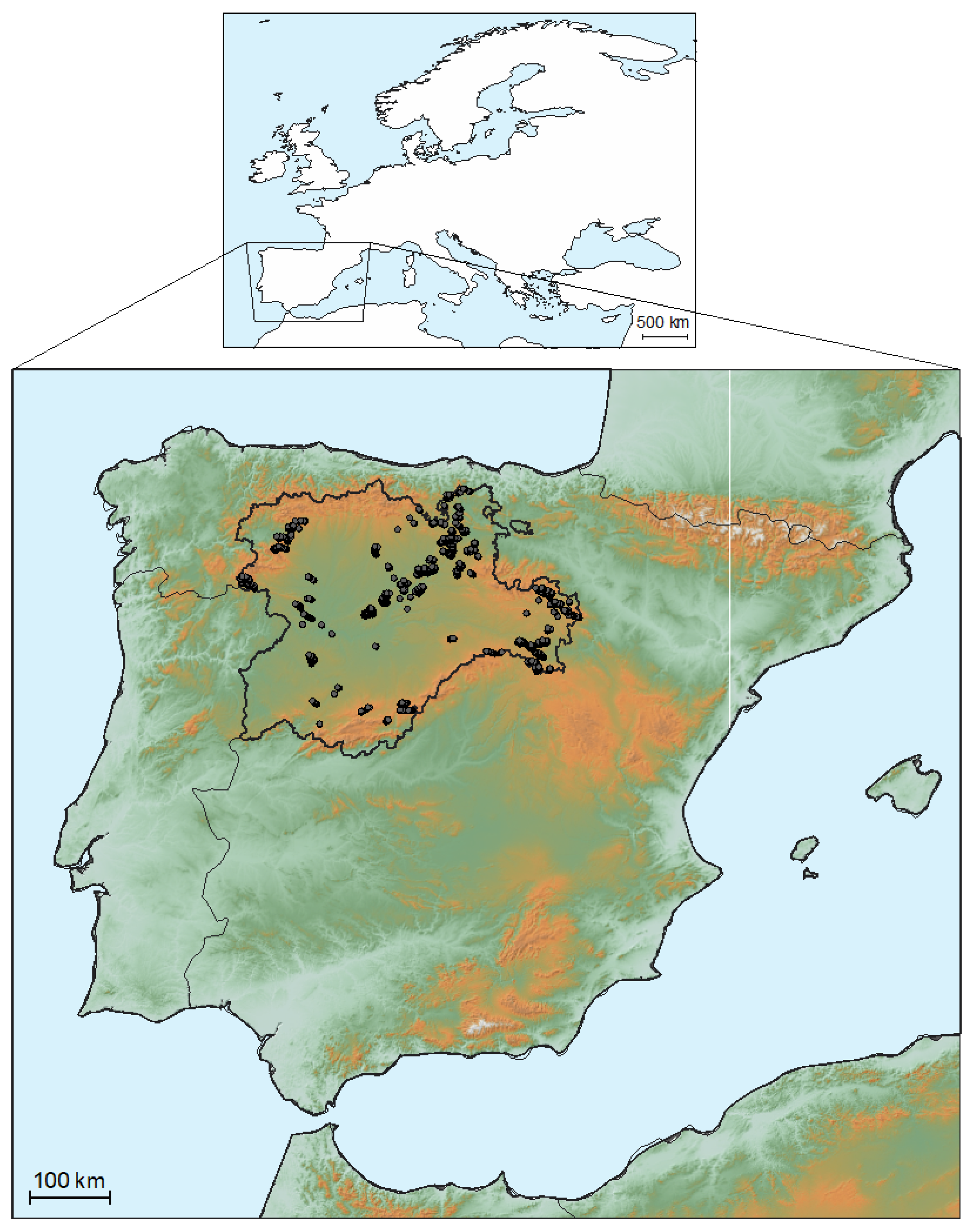
2.1. Study Area and Materials
2.2. Statistical Analyses
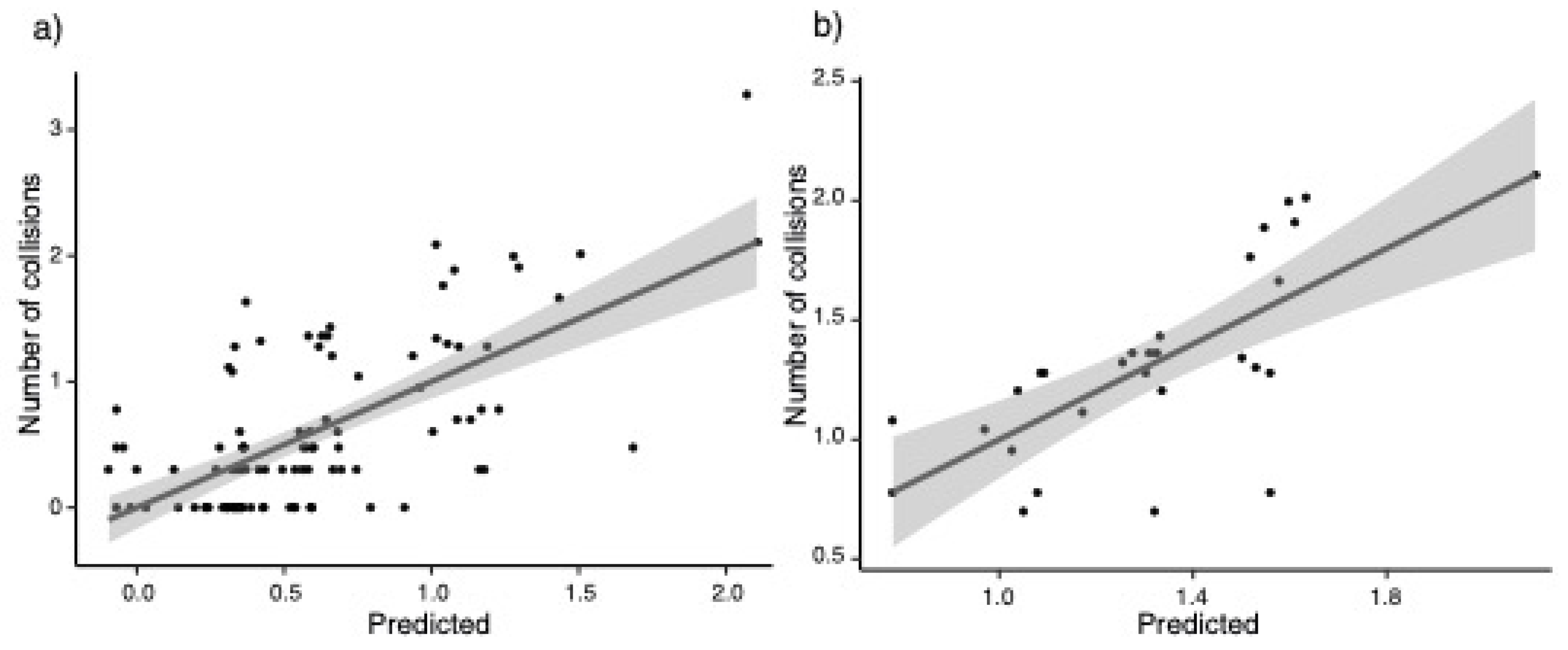
3. Results
4. Discussion
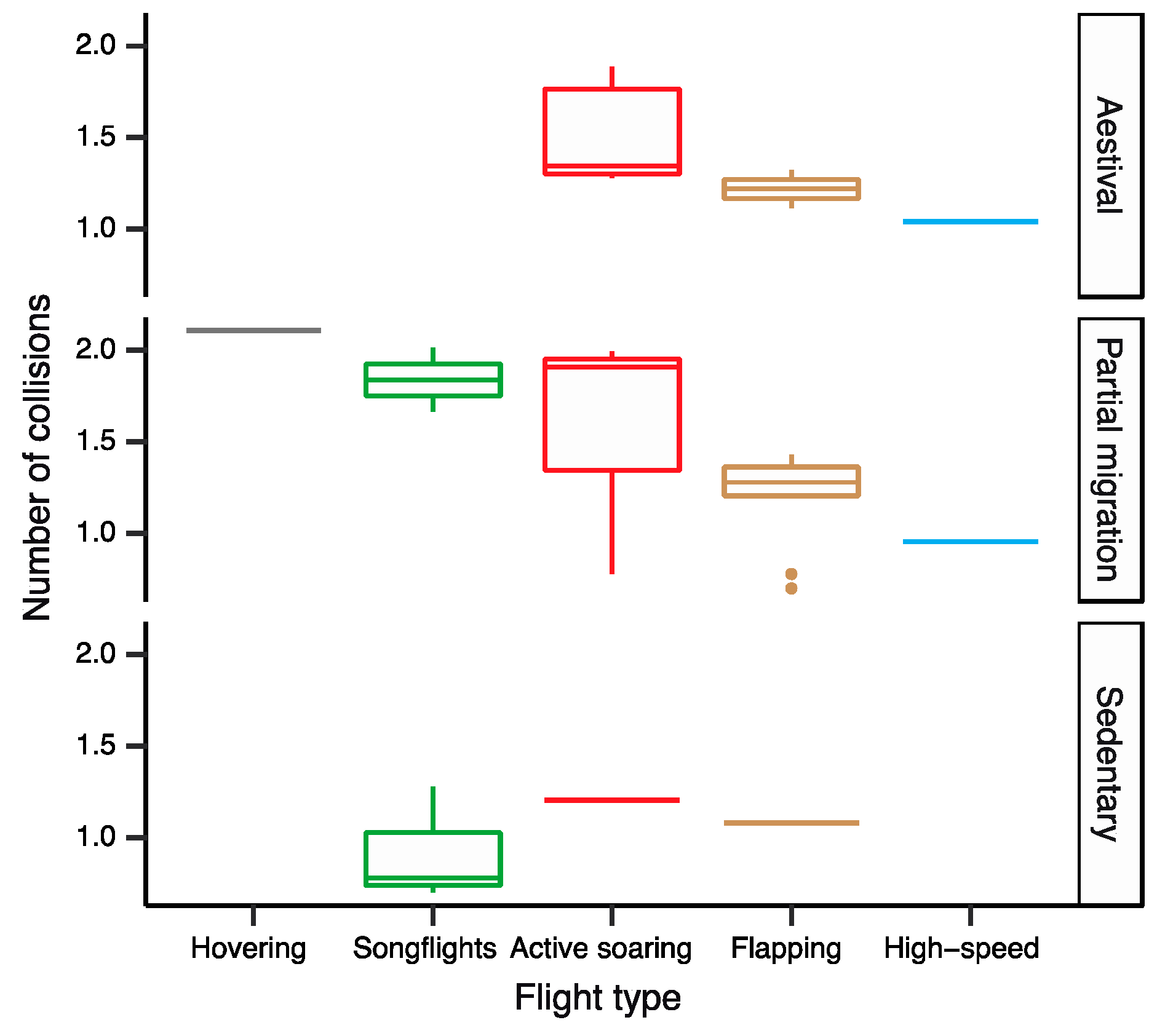
4.1. Flight Type
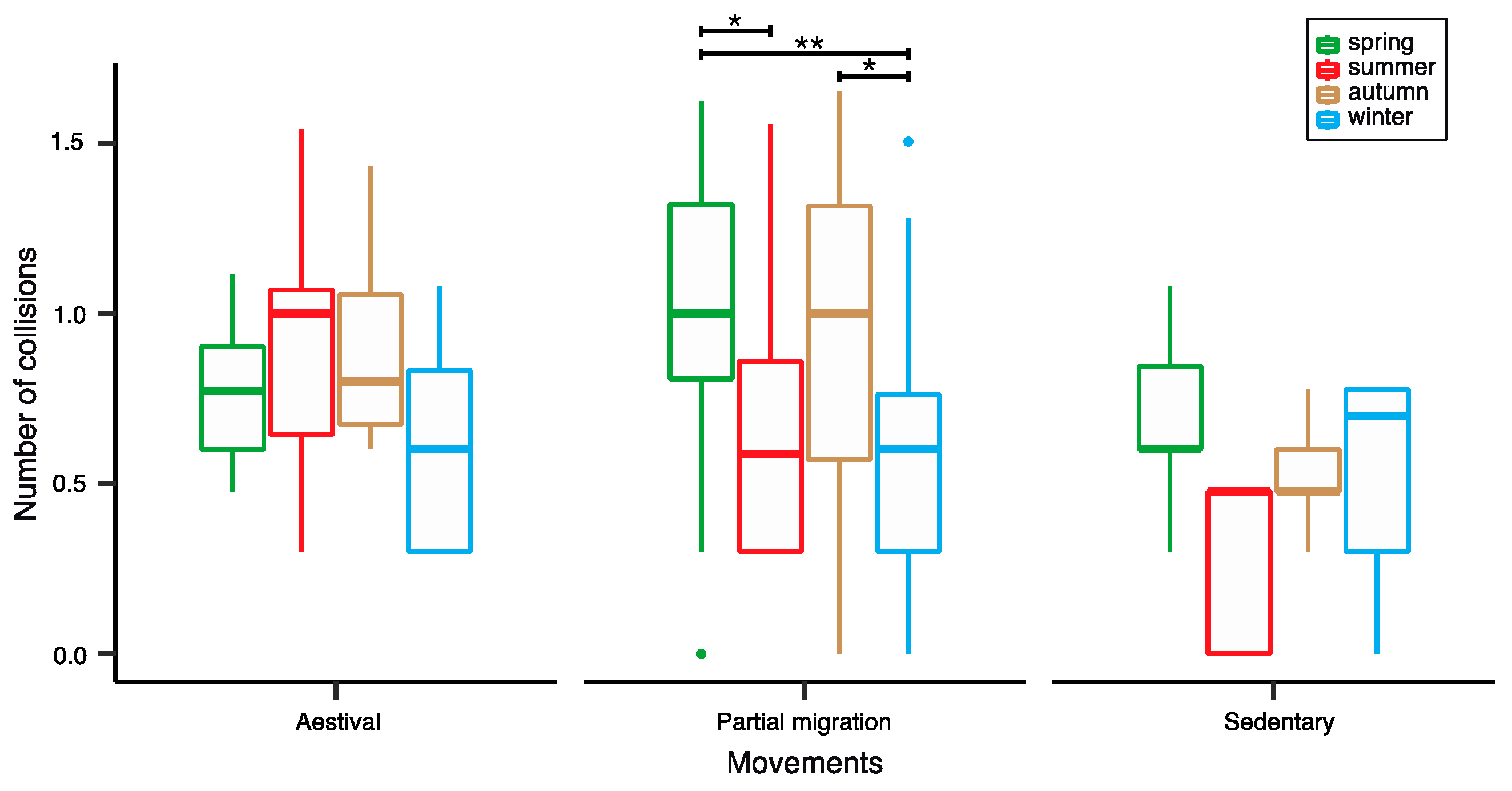
4.2. Seasonality
4.3. Species and Group
4.4. Abundance and Size
4.5. Sex
4.6. Modelling Approaches to Quantify the Theoretical Risk of Collision in Wind Farms and Ways to Avoid it
4.7. Limitations of this Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; et al. The Impact of Conservation on the Status of the World’s Vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, N.S.; Ehrlich, P.R. (Eds.) Conservation Biology for All; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Loss, S.R.; Will, T.; Marra, P.P. Direct human-caused mortality of birds: Improving quantification of magnitude and assessment of population impact. Front. Ecol. Environ. 2012, 10, 357–364. [Google Scholar] [CrossRef]
- Stewart, G.B.; Pullin, A.S.; Coles, C.F. Poor evidence-base for assessment of wind farm impacts on birds. Environ. Conserv. 2007, 34, 1–11. [Google Scholar] [CrossRef]
- Thaxter, C.B.; Buchanan, G.M.; Carr, J.; Butchart, S.H.; Newbold, T.; Green, R.E.; Tobias, J.A.; Foden, W.B.; O’Brien, S.; Pearce-Higgins, J.W. Bird and bat species’ global vulnerability to collision mortality at wind farms revealed through a trait-based assessment. Proc. R. Soc. B 2017, 284, 20170829. [Google Scholar] [CrossRef]
- Carrete, M.; Sánchez-Zapata, J.A.; Benítez, J.R.; Lobón, M.; Donázar, J.A. Large scale risk-assessment of wind-farms on population viability of a globally endangered long-lived raptor. Biol. Conserv. 2009, 142, 2954–2961. [Google Scholar] [CrossRef]
- Drewitt, A.L.; Langston, R.H.W. Assessing the impacts of wind farms on birds. IBIS 2006, 148, 29–42. [Google Scholar] [CrossRef]
- Farfán, M.A.; Vargas, J.M.; Duarte, J.D.; Real, R. What is the impact of wind farms on birds? A case of study in southern Spain. Biodivers. Conserv. 2009, 18, 3743–3758. [Google Scholar] [CrossRef]
- Tellería, J.L. Wind power plants and the conservation of birds and bats in Spain: A geographical assessment. Biodivers. Conserv. 2008, 18, 1781–1791. [Google Scholar] [CrossRef]
- Saidur, R.; Rahim, N.A.; Islam, M.R.; Solangi, K.H. Environmental impact of wind energy. Renew. Sustain. Energy Rev. 2011, 15, 2423–2430. [Google Scholar] [CrossRef]
- Williams, D.R.; Child, M.F.; Dicks, L.V.; Ockendon, N.; Pople, R.G.; Showler, D.A.; Walsh, J.C.; Ermgassen, E.K.H.J.; Sutherland, W.J. Bird Conservation. In What Works in Conservation 2019; Sutherland, W., Dicks, L., Ockendon, N., Petrovan, S., Eds.; Open Book Publishers: Cambridge, UK, 2019; pp. 141–290. [Google Scholar]
- Serrano, D.; Margalida, A.; Pérez-García, J.M.; Juste, J.; Traba, J.; Valera, F.; Carrete, M.; Aihartza, J.; Real, J.; Mañosa, S.; et al. Renewables in Spain threaten biodiversity. Science 2020, 370, 1282–1283. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Rehfisch, M.R.; Fox, A.D.; Desholm, M.; Anthony, S.J. The effect of avoidance rates on bird mortality predictions made by wind turbine collision risk models. IBIS 2006, 148, 198–202. [Google Scholar] [CrossRef]
- Larsen, J.K.; Guillemette, M. Effects of wind turbines on flight behaviour of wintering common eiders: Implications for habitat use and collision risk. J. Appl. Ecol. 2007, 44, 516–522. [Google Scholar] [CrossRef]
- Marques, A.T.; Santos, C.D.; Hanssen, F.; Muñoz, A.R.; Onrubia, A.; Wikelski, M.; Moreira, F.; Palmeirim, J.M.; Silva, J.P. Wind turbines cause functional habitat loss for migratory soaring birds. J. Anim. Ecol. 2020, 89, 93–103. [Google Scholar] [CrossRef]
- Marques, A.T.; Batalha, H.; Bernardino, J. Bird Displacement by Wind Turbines: Assessing Current Knowledge and Recommendations for Future Studies. Birds 2021, 2, 460–475. [Google Scholar] [CrossRef]
- Masden, E.; Cook, A. Avian collision risk models for wind energy impact assessments. Environ. Impact Assess. Rev. 2015, 56, 43–49. [Google Scholar] [CrossRef]
- Tanskanen, A. Impact on breeding birds of a semi-offshore island-based windmill park in Åland, Northern Baltic Sea. Ornis Svec. 2012, 22, 9–15. [Google Scholar] [CrossRef]
- Balotari-Chiebao, F.; Valkama, J.; Byholm, P. Assessing the vulnerability of breeding bird populations to onshore wind-energy developments in Finland. Ornis Fenn. 2021, 98, 59–73. [Google Scholar]
- Zimmerling, J.R.; Pomeroy, A.C.; D’Entremont, M.V.; Francis, C.M. Canadian Estimate of Bird Mortality Due to Collisions and Direct Habitat Loss Associated with Wind Turbine Developments. Avian Conserv. Ecol. 2013, 8, 10. [Google Scholar] [CrossRef]
- Dahl, E.L.; Bevanger, K.; Nygård, T.; Røskaft, E.; Stokke, B.G. Reduced breeding success in white-tailed eagles at Smøla windfarm, western Norway, is caused by mortality and displacement. Biol. Conserv. 2012, 145, 79–85. [Google Scholar] [CrossRef]
- De Lucas, M.; Ferrer, M.; Bechard, M.J.; Muñoz, A.R. Griffon vulture mortality at wind farms in southern Spain: Distribution of fatalities and active mitigation measures. Biol. Conserv. 2012, 147, 184–189. [Google Scholar] [CrossRef]
- Carrete, M.; Sánchez-Zapata, J.A.; Benítez, J.R.; Lobón, M.; Montoya, F.; Donázar, J.A. Mortality at wind-farms is positively related to large-scale distribution and aggregation in griffon vultures. Biol. Conserv. 2012, 145, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.; de Lucas, M.; Janss, G.F.E.; Casado, E.; Muñoz, A.R.; Bechard, M.J.; Calabuig, C.P. Weak relationship between risk assessment studies and recorded mortality in wind farms. J. Appl. Ecol. 2011, 49, 38–46. [Google Scholar] [CrossRef]
- Gómez-Catasús, J.; Garza, V.; Traba, J. Wind farms affect the occurrence, abundance and population trends of small passerine birds: The case of the Dupont’s lark. J. Appl. Ecol. 2018, 55, 2033–2042. [Google Scholar] [CrossRef]
- Martin, G.; Shaw, J. Bird collisions with power lines: Failing to see the way ahead? Biol. Conserv. 2010, 143, 2695–2702. [Google Scholar] [CrossRef]
- Fernández-López, J.; Blanco-Aguiar, J.A.; Vicente, J.; Acevedo, P. Can we model distribution of population abundance from wildlife–vehicles collision data? Ecography 2022, 2022, e06113. [Google Scholar] [CrossRef]
- Osborn, R.G.; Dieter, C.D.; Higgins, K.F.; Usgaard, R.E. Bird Flight Characteristics Near Wind Turbines in Minnesota. Am. Midl. Nat. 1998, 139, 29–38. [Google Scholar] [CrossRef]
- Marques, A.T.; Batalha, H.; Rodrigues, S.; Costa, H.; Pereira, M.J.R.; Fonseca, C.; Mascarenhas, M.; Bernardino, J. Understanding bird collisions at wind farms: An updated review on the causes and possible mitigation strategies. Biol. Conserv. 2014, 179, 40–52. [Google Scholar] [CrossRef]
- Smallwood, K.S.; Rugge, L.; Morrison, M.L. Influence of Behavior on Bird Mortality in Wind Energy Developments. J. Wildl. Manag. 2009, 73, 1082–1098. [Google Scholar] [CrossRef]
- Global Wind Energy Council Data. Available online: http://www.gwec.net/wp-content/uploads/2017/04/5_Top-10-cumulative-capacity-Dec-2016-1.jpg (accessed on 31 January 2020).
- AEE (Asociación Empresarial Eólica). 2017 La Voz del Sector. Available online: https://www.aeeolica.org/es/comunicacion/publicaciones-aee/anuarios (accessed on 31 January 2020).
- Balmori, A. Endangered bird mortality by gunshots: Still a current problem. Biodivers. Conserv. 2019, 28, 2555–2564. [Google Scholar] [CrossRef]
- Peterson, R.; Mountfort, G.; Hollom, P.A.D. Guía de Campo de las Aves de España y de Europa; Omega: Barcelona, Spain, 1973. [Google Scholar]
- Madders, M.; Whitfield, D.P. Upland raptors and the assessment of wind farm impacts. IBIS 2006, 148, 43–56. [Google Scholar] [CrossRef]
- Mullarney, T.; Svensson, L.; Zetterström, D.; Grant, P.J. The Complete Guide to the Birds of Europe; Harper Collins Publishers: London, UK, 2000. [Google Scholar]
- Heath, M.F.; Hagemeijer, W.J. European Bird Populations: Estimates and Trends; (No. 10); BirdLife International: Cambridge, UK, 2000. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 31 January 2020).
- Rydell, J.; Bach, L.; Dubourg-Savage, M.-J.; Green, M.; Rodrigues, L.; Hedenström, A. Mortality of bats at wind turbines links to nocturnal insect migration? Eur. J. Wildl. Res. 2010, 56, 823–827. [Google Scholar] [CrossRef] [Green Version]
- De Barrios, L.; Rodriguez, A. Behavioural and environmental correlates of soaring-bird mortality at on-shore wind turbines. J. Appl. Ecol. 2004, 41, 72–81. [Google Scholar] [CrossRef]
- Thaxter, C.B.; Ross-Smith, V.H.; Bouten, W.; Clark, N.A.; Conway, G.J.; Masden, E.A.; Clewley, G.D.; Barber, L.J.; Burton, N.H.K. Avian vulnerability to wind farm collision through the year: Insights from lesser black-backed gulls (Larus fuscus) tracked from multiple breeding colonies. J. Appl. Ecol. 2019, 56, 2410–2422. [Google Scholar] [CrossRef]
- Desholm, M. Avian sensitivity to mortality: Prioritising migratory bird species for assessment at proposed wind farms. J. Environ. Manag. 2009, 90, 2672–2679. [Google Scholar] [CrossRef]
- Aschwanden, J.; Stark, H.; Peter, D.; Steuri, T.; Schmid, B.; Liechti, F. Bird collisions at wind turbines in a mountainous area related to bird movement intensities measured by radar. Biol. Conserv. 2018, 220, 228–236. [Google Scholar] [CrossRef]
- De Kikuchi, R. Adverse impacts of wind power generation on collision behaviour of birds and anti-predator behaviour of squirrels. J. Nat. Conserv. 2008, 16, 44–55. [Google Scholar] [CrossRef]
- Tellería, J.L. Potential impacts of wind farms on migratory birds crossing Spain. Bird Conserv. Int. 2009, 19, 131–136. [Google Scholar] [CrossRef]
- De Lucas, M.; Janss, G.F.; Whitfield, D.P.; Ferrer, M. Collision fatality of raptors in wind farms does not depend on raptor abundance. J. Appl. Ecol. 2008, 45, 1695–1703. [Google Scholar] [CrossRef]
- Duriez, O.; Pilard, P.; Saulnier, N.; Boudarel, P.; Besnard, A. Windfarm collisions in medium-sized raptors: Even increasing populations can suffer strong demographic impacts. Anim. Conserv. 2022. [Google Scholar] [CrossRef]
- Morinha, F.; Travassos, P.J.; Seixas, F.; Martins, A.; Bastos, R.; Carvalho, D.; Magalhães, P.C.L.F.D.S.; Santos, M.; Bastos, E.; Cabral, J. Differential mortality of birds killed at wind farms in Northern Portugal. Bird Study 2014, 61, 255–259. [Google Scholar] [CrossRef]
- Stienen, E.W.M.; Courtens, W.; Everaert, J.; Van De Walle, M. Sex-Biased Mortality of Common Terns in Wind Farm Collisions. Condor 2008, 110, 154–157. [Google Scholar] [CrossRef] [Green Version]
- Band, W.; Madders, M.; Whitfield, D.P. Developing field and analytical methods to assess avian collision risk at wind farms. In Birds and Wind Power; De Lucas, M., Janss, G., Ferrer, M., Eds.; Lynx Edicions: Barcelona, Spain, 2005. [Google Scholar]
- Kunz, T.H.; Arnett, E.B.; Cooper, B.M.; Erickson, W.P.; Larkin, R.P.; Mabee, T.; Morrison, M.L.; Strickland, M.D.; Szewczak, J.M. Assessing impacts of wind-energy development on nocturnally active birds and bats: A guidance document. J. Wildl. Manag. 2007, 71, 2449–2483. [Google Scholar] [CrossRef]
- Atienza, J.C.; Martín, I.; Infante, O.; Valls, J.; Domínguez, J. Directrices para la Evaluación del Impacto de los Parques Eólicos en aves y Murciélagos (Versión 3.0); SEO/BirdLife: Madrid, Spain, 2012. [Google Scholar]
- González, F.; Alcalde, J.T.; Ibáñez, C. Directrices Básicas para el Estudio del Impacto de Instalaciones Eólicas Sobre Poblaciones de Murciélagos en España; SECEMU: Barbastella, Spain, 2013; Volume 6, pp. 1–31. [Google Scholar]
- Tikkanen, H.; Balotari-Chiebao, F.; Laaksonen, T.; Pakanen, V.M.; Rytkönen, S. Habitat use of flying subadult White-tailed Eagles (Haliaeetus albicilla): Implications for land use and wind power plant planning. Ornis Fenn. 2018, 95, 137–150. [Google Scholar]
- Tikkanen, H.; Rytkönen, S.; Karlin, O.-P.; Ollila, T.; Pakanen, V.-M.; Tuohimaa, H.; Orell, M. Modelling golden eagle habitat selection and flight activity in their home ranges for safer wind farm planning. Environ. Impact Assess. Rev. 2018, 71, 120–131. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. Estimates of bird collision mortality at wind facilities in the contiguous United States. Biol. Conserv. 2013, 168, 201–209. [Google Scholar] [CrossRef]
- Rodríguez, B.; Rodríguez, A.; Siverio, F.; Siverio, M. Causes of Raptor Admissions to a Wildlife Rehabilitation Center in Tenerife (Canary Islands). J. Raptor Res. 2010, 44, 30–39. [Google Scholar] [CrossRef]
- Schuster, E.; Bulling, L.; Köppel, J. Consolidating the State of Knowledge: A Synoptical Review of Wind Energy’s Wildlife Effects. Environ. Manag. 2015, 56, 300–331. [Google Scholar] [CrossRef]
- Del Valle, J.D.; Peralta, F.C.; Arjona, M.I.J. Factors affecting carcass detection at wind farms using dogs and human searchers. J. Appl. Ecol. 2020, 57, 1926–1935. [Google Scholar] [CrossRef]
- Erickson, W.P.; Wolfe, M.M.; Bay, K.J.; Johnson, U.H.; Gehring, J.L. A Comprehensive Analysis of Small-Passerine Fatalities from Collision with Turbines at Wind Energy Facilities. PLoS ONE 2014, 9, e107491. [Google Scholar] [CrossRef]
- Soares, M.; Bernardino, J.; Nogueira, A.J.A.; Bispo, R. An avian relative fatality risk index for Iberian species on wind farms based on zero inflated count models. Bachelor’s Degree Thesis, University of Aveiro, Aveiro, Portugal, 2014; pp. 21–42. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Group | Seasonal Movement Type | IUCN Category | Flight Type | Length | Spanish Population | European Population | Number of Collisions | Spring | Summer | Autumn | Winter | Predicted Values |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accipiter gentilis | Bird of prey | Sedentary | Least concern | Flapping | 53 | 2650 | 155,000 | 1 | 0 | 1 | 0 | 0 | 1.1 |
| Accipiter nisus | Bird of prey | Partial migration | Least concern | Flapping | 32 | 3500 | 330,000 | 19 | 8 | 4 | 3 | 4 | 2.2 * |
| Aegypius monachus | Bird of prey | Sedentary | Near threatened | Passive soaring | 107 | 1100 | 1400 | 3 | 1 | 2 | 0 | 0 | 48.4 |
| Alauda arvensis | Passerine | Partial migration | Least concern | Songflights | 17 | 4,000,000 | 40,000,000 | 103 | 37 | 26 | 22 | 18 | 32.1 * |
| Alectoris rufa | Galliforme | Sedentary | Least concern | Flapping | 34 | 2,600,000 | 3,300,000 | 43 | 11 | 5 | 16 | 11 | 2.3 |
| Anthus campestris | Passerine | Aestival | Least concern | Flapping | 17 | 520,000 | 1,000,000 | 4 | 3 | 0 | 1 | 0 | 2.2 |
| Anthus pratensis | Passerine | Partial migration | Near threatened | Flapping | 15 | 0 | 15,000,000 | 6 | 2 | 1 | 0 | 3 | 0.9 * |
| Anthus spinoletta | Passerine | Partial migration | Least concern | Flapping | 16 | 24,000 | 450,000 | 1 | 1 | 0 | 0 | 0 | 2.7 |
| Anthus trivialis | Passerine | Aestival | Least concern | Flapping | 15 | 350,000 | 26,000,000 | 1 | 0 | 0 | 1 | 0 | 2.1 |
| Apus apus | Apodiforme | Aestival | Least concern | High-speed | 18 | 525,000 | 7,500,000 | 123 | 30 | 27 | 35 | 31 | 10.4 |
| Apus melba | Apodiforme | Aestival | Least concern | High-speed | 22 | 5850 | 165,000 | 1 | 0 | 1 | 0 | 0 | 6.2 |
| Apus pallidus | Apodiforme | Aestival | Least concern | High-speed | 17 | 2200 | 28,000 | 2 | 0 | 0 | 1 | 1 | 5.6 |
| Aquila chrysaetos | Bird of prey | Sedentary | Least concern | Active soaring | 86 | 1230 | 9000 | 16 | 6 | 0 | 5 | 5 | 8.6 * |
| Ardea cinerea | Pelecaniforme | Partial migration | Least concern | Flapping | 93 | 1510 | 180,000 | 1 | 1 | 0 | 0 | 0 | 2.0 |
| Asio flammeus | Bird of prey | Partial migration | Least concern | Flapping | 37 | 3 | 70,000 | 2 | 1 | 0 | 0 | 1 | 1.0 |
| Asio otus | Bird of prey | Partial migration | Least concern | Flapping | 34 | 5700 | 450,000 | 1 | 1 | 0 | 0 | 0 | 2.3 |
| Bubo bubo | Bird of prey | Sedentary | Least concern | Flapping | 65 | 560 | 28,000 | 3 | 2 | 0 | 1 | 0 | 0.9 |
| Burhinus oedicnemus | Caradriforme | Partial migration | Least concern | Flapping | 42 | 26,000 | 100,000 | 1 | 0 | 1 | 0 | 0 | 2.7 |
| Buteo buteo | Bird of prey | Partial migration | Least concern | Active soaring | 52 | 5250 | 850,000 | 81 | 23 | 20 | 30 | 8 | 19.8 * |
| Caprimulgus europaeus | Caprimulgiformes | Aestival | Least concern | Flapping | 26 | 100,000 | 535,000 | 2 | 1 | 0 | 1 | 0 | 1.9 |
| Carduelis cannabina | Passerine | Partial migration | Least concern | Flapping | 13 | 2,500,000 | 14,500,000 | 27 | 12 | 4 | 10 | 1 | 4.5 * |
| Carduelis carduelis | Passerine | Partial migration | Least concern | Flapping | 13 | 1,850,000 | 16,000,000 | 5 | 1 | 1 | 2 | 1 | 4.4 * |
| Ciconia ciconia | Ciconiforme | Aestival | Least concern | Active soaring | 103 | 7901 | 140,000 | 5 | 1 | 4 | 0 | 0 | 12.2 |
| Ciconia nigra | Ciconiforme | Aestival | Least concern | Active soaring | 98 | 210 | 7900 | 1 | 0 | 1 | 0 | 0 | 8.1 |
| Circaetus gallicus | Bird of prey | Aestival | Least concern | Active soaring | 65 | 1900 | 9900 | 22 | 3 | 9 | 7 | 3 | 10.4 * |
| Circus aeruginosus | Bird of prey | Partial migration | Least concern | Active soaring | 49 | 500 | 70,000 | 2 | 1 | 1 | 0 | 0 | 15.1 |
| Circus cyaneus | Bird of prey | Partial migration | Least concern | Active soaring | 50 | 350 | 26,500 | 2 | 1 | 1 | 0 | 0 | 14.5 |
| Circus pygargus | Bird of prey | Aestival | Least concern | Active soaring | 45 | 4100 | 38,000 | 20 | 7 | 9 | 3 | 1 | 11.3 * |
| Clamator glandarius | Cuculiformes | Aestival | Least concern | Flapping | 37 | 60,000 | 65,000 | 1 | 0 | 0 | 1 | 0 | 1.8 |
| Coccothraustes coccothraustes | Passerine | Partial migration | Least concern | Flapping | 17 | 4500 | 1,500,000 | 1 | 0 | 1 | 0 | 0 | 2.2 |
| Columba livia | Columbiformes | Sedentary | Least concern | Flapping | 32 | 2,140,000 | 13,000,000 | 3 | 2 | 0 | 1 | 0 | 2.3 |
| Columba palumbus | Columbiformes | Partial migration | Least concern | Flapping | 40 | 220,000 | 11,500,000 | 2 | 1 | 0 | 1 | 0 | 3.4 |
| Corvus corax | Passerine | Sedentary | Least concern | Flapping | 60 | 75,000 | 830,000 | 1 | 0 | 0 | 0 | 1 | 1.6 |
| Corvus corone | Passerine | Sedentary | Least concern | Flapping | 48 | 425,000 | 13,000,000 | 3 | 1 | 2 | 0 | 0 | 1.9 |
| Coturnix coturnix | Galliforme | Partial migration | Least concern | Flapping | 17 | 380,000 | 1,600,000 | 3 | 0 | 0 | 2 | 1 | 3.7 |
| Cuculus canorus | Cuculiformes | Aestival | Least concern | Flapping | 34 | 230,000 | 2,700,000 | 1 | 0 | 1 | 0 | 0 | 2.0 |
| Delichon urbica | Passerine | Aestival | Least concern | Flapping | 14 | 2,150,000 | 21,000,000 | 21 | 6 | 2 | 4 | 9 | 2.6 * |
| Emberiza cia | Passerine | Sedentary | Least concern | Flapping | 16 | 1,400,000 | 2,600,000 | 2 | 0 | 0 | 1 | 1 | 2.2 |
| Emberiza cirlus | Passerine | Sedentary | Least concern | Flapping | 16 | 650,000 | 2,800,000 | 1 | 1 | 0 | 0 | 0 | 2.0 |
| Emberiza citrinella | Passerine | Partial migration | Least concern | Flapping | 16 | 155,000 | 80,000,000 | 1 | 1 | 0 | 0 | 0 | 3.3 |
| Erithacus rubecula | Passerine | Partial migration | Least concern | Flapping | 13 | 2,100,000 | 100,000,000 | 23 | 9 | 4 | 9 | 1 | 4.4 * |
| Falco columbarius | Bird of prey | Partial migration | Least concern | High-speed | 30 | 0 | 46,000 | 3 | 0 | 1 | 0 | 2 | 3.9 |
| Falco naumanni | Bird of prey | Aestival | Least concern | Active soaring | 30 | 6500 | 15,000 | 77 | 12 | 34 | 26 | 5 | 11.9 * |
| Falco peregrinus | Bird of prey | Partial migration | Least concern | High-speed | 45 | 1680 | 9300 | 9 | 6 | 1 | 2 | 0 | 9.2 * |
| Falco subbuteo | Bird of prey | Aestival | Least concern | High-speed | 32 | 2500 | 90,000 | 11 | 2 | 4 | 4 | 1 | 5.6 * |
| Falco tinnunculus | Bird of prey | Partial migration | Least concern | Hovering | 34 | 27,500 | 370,000 | 128 | 41 | 35 | 44 | 8 | 128.0 * |
| Ficedula hypoleuca | Passerine | Aestival | Least concern | Flapping | 13 | 240,000 | 10,000,000 | 13 | 3 | 1 | 8 | 1 | 2.0 * |
| Fringilla coelebs | Passerine | Partial migration | Least concern | Flapping | 15 | 4,500,000 | 160,000,000 | 3 | 0 | 1 | 0 | 2 | 4.8 |
| Galerida cristata | Passerine | Sedentary | Least concern | Songflights | 18 | 700,000 | 4,600,000 | 5 | 1 | 2 | 2 | 0 | 13.6 * |
| Galerida theklae | Passerine | Sedentary | Least concern | Songflights | 16 | 1,500,000 | 1,550,000 | 6 | 3 | 0 | 2 | 1 | 14.8 * |
| Gallinago gallinago | Caradriformes | Partial migration | Least concern | Flapping | 26 | 70 | 6,400,000 | 1 | 1 | 0 | 0 | 0 | 1.4 |
| Garrulus glandarius | Passerine | Partial migration | Least concern | Flapping | 34 | 800,000 | 13,500,000 | 3 | 1 | 0 | 1 | 1 | 4.0 |
| Grus grus | Gruiformes | Partial migration | Least concern | Flapping | 107 | 0 | 67,000 | 3 | 2 | 1 | 0 | 0 | 0.9 |
| Gyps fulvus | Bird of prey | Partial migration | Least concern | Passive soaring | 102 | 8074 | 10,200 | 1901 | 693 | 305 | 518 | 385 | 117.8 |
| Hieraaetus pennatus | Bird of prey | Aestival | Least concern | Active soaring | 47 | 3000 | 5200 | 58 | 7 | 18 | 22 | 11 | 10.9 * |
| Hirundo rustica | Passerine | Aestival | Least concern | Flapping | 19 | 795,000 | 26,000,000 | 2 | 0 | 0 | 0 | 2 | 2.3 |
| Jynx torquilla | Piciformes | Aestival | Least concern | Flapping | 17 | 49,000 | 550,000 | 1 | 0 | 0 | 0 | 1 | 1.7 |
| Lanius collurio | Passerine | Aestival | Least concern | Flapping | 17 | 370,000 | 4,500,000 | 1 | 0 | 1 | 0 | 0 | 2.2 |
| Lanius meridionalis | Passerine | Partial migration | Least concern | Flapping | 24 | 225,000 | 300,000 | 1 | 1 | 0 | 0 | 0 | 3.4 |
| Locustella naevia | Passerine | Aestival | Least concern | Flapping | 13 | 275 | 1,000,000 | 1 | 0 | 0 | 0 | 1 | 1.0 |
| Loxia curvirostra | Passerine | Sedentary | Least concern | Flapping | 16 | 165,000 | 2,000,000 | 1 | 1 | 0 | 0 | 0 | 1.7 |
| Lullula arborea | Passerine | Partial migration | Least concern | Songflights | 14 | 900,000 | 2,200,000 | 46 | 19 | 6 | 19 | 2 | 27.1 * |
| Luscinia megarhynchos | Passerine | Aestival | Least concern | Flapping | 16 | 1,100,000 | 9,500,000 | 1 | 0 | 1 | 0 | 0 | 2.4 |
| Melanocorypha calandra | Passerine | Sedentary | Least concern | Songflights | 18 | 2,200,000 | 10,000,000 | 19 | 11 | 2 | 1 | 5 | 15.5 * |
| Miliaria calandra | Passerine | Partial migration | Least concern | Flapping | 18 | 2,800,000 | 11,000,000 | 16 | 7 | 2 | 4 | 3 | 4.6 * |
| Milvus migrans | Bird of prey | Aestival | Least concern | Active soaring | 53 | 9000 | 85,000 | 19 | 4 | 9 | 3 | 3 | 12.4 * |
| Milvus milvus | Bird of prey | Partial migration | Near threatened | Active soaring | 67 | 3700 | 21,500 | 99 | 28 | 7 | 33 | 31 | 19.0 * |
| Motacilla alba | Passerine | Partial migration | Least concern | Flapping | 18 | 200,000 | 15,000,000 | 1 | 0 | 1 | 0 | 0 | 3.4 |
| Neophron percnopterus | Bird of prey | Partial migration | Endangered | Active soaring | 60 | 1350 | 5000 | 6 | 0 | 1 | 2 | 3 | 16.9 * |
| Oenanthe oenanthe | Passerine | Aestival | Least concern | Flapping | 15 | 345,000 | 9,000,000 | 1 | 0 | 0 | 1 | 0 | 2.1 |
| Otis tarda | Gruiformes | Sedentary | Vulnerable | Flapping | 90 | 18,000 | 29,500 | 2 | 1 | 1 | 0 | 0 | 1.3 |
| Parus major | Passerine | Partial migration | Least concern | Flapping | 14 | 3,000,000 | 110,000,000 | 2 | 0 | 0 | 0 | 2 | 4.6 |
| Passer domesticus | Passerine | Sedentary | Least concern | Flapping | 15 | 9,600,000 | 120,000,000 | 2 | 0 | 0 | 2 | 0 | 2.7 |
| Pernis apivorus | Bird of prey | Aestival | Least concern | Active soaring | 55 | 1500 | 125,000 | 4 | 0 | 1 | 2 | 1 | 10.1 |
| Petronia petronia | Passerine | Sedentary | Least concern | Flapping | 16 | 1,000,000 | 1,100,000 | 12 | 3 | 2 | 3 | 4 | 2.1 * |
| Phoenicurus ochruros | Passerine | Partial migration | Least concern | Flapping | 14 | 650,000 | 4,500,000 | 1 | 0 | 1 | 0 | 0 | 3.9 |
| Phylloscopus bonelli | Passerine | Aestival | Least concern | Flapping | 11 | 1,900,000 | 2,600,000 | 2 | 0 | 0 | 1 | 1 | 2.6 |
| Phylloscopus collybita | Passerine | Partial migration | Least concern | Flapping | 11 | 550,000 | 80,000,000 | 23 | 9 | 1 | 9 | 4 | 3.8 * |
| Phylloscopus trochilus | Passerine | Aestival | Least concern | Flapping | 12 | 60 | 100,000,000 | 2 | 0 | 0 | 0 | 2 | 0.8 |
| Pica pica | Passerine | Sedentary | Least concern | Flapping | 45 | 700,000 | 21,000,000 | 1 | 1 | 0 | 0 | 0 | 2.0 |
| Podiceps cristatus | Podicipediformes | Partial migration | Least concern | Flapping | 48 | 2800 | 800,000 | 1 | 0 | 0 | 0 | 1 | 2.1 |
| Prunella modularis | Passerine | Partial migration | Least concern | Flapping | 14 | 700,000 | 17,000,000 | 1 | 0 | 0 | 1 | 0 | 3.9 |
| Ptyonoprogne rupestris | Passerine | Partial migration | Least concern | Flapping | 15 | 92,000 | 300,000 | 2 | 0 | 0 | 1 | 1 | 3.1 |
| Regulus ignicapillus | Passerine | Partial migration | Least concern | Flapping | 10 | 1,400,000 | 4,100,000 | 23 | 14 | 2 | 7 | 0 | 4.2 * |
| Regulus regulus | Passerine | Partial migration | Least concern | Flapping | 9 | 370,000 | 70,000,000 | 2 | 0 | 0 | 1 | 1 | 3.6 |
| Saxicola torquata | Passerine | Partial migration | Least concern | Flapping | 12 | 500,000 | 2,100,000 | 3 | 0 | 0 | 3 | 0 | 3.8 |
| Scolopax rusticola | Caradriformes | Partial migration | Least concern | Flapping | 35 | 660,000 | 2,200,000 | 1 | 0 | 0 | 0 | 1 | 3.9 |
| Serinus serinus | Passerine | Partial migration | Least concern | Flapping | 12 | 5,300,000 | 9,600,000 | 2 | 0 | 1 | 1 | 0 | 4.9 |
| Sturnus unicolor | Passerine | Sedentary | Least concern | Flapping | 21 | 2,125,000 | 2,800,000 | 3 | 0 | 1 | 1 | 1 | 2.3 |
| Sturnus vulgaris | Passerine | Partial migration | Least concern | Flapping | 21 | 600,000 | 80,000,000 | 4 | 0 | 1 | 2 | 1 | 3.9 |
| Sylvia atricapilla | Passerine | Partial migration | Least concern | Flapping | 14 | 1,150,000 | 30,000,000 | 19 | 4 | 1 | 10 | 4 | 4.2 * |
| Sylvia borin | Passerine | Aestival | Least concern | Flapping | 14 | 550,000 | 14,000,000 | 2 | 1 | 1 | 0 | 0 | 2.3 |
| Sylvia communis | Passerine | Aestival | Least concern | Flapping | 14 | 525,000 | 15,000,000 | 1 | 0 | 0 | 1 | 0 | 2.2 |
| Sylvia hortensis | Passerine | Aestival | Least concern | Flapping | 15 | 310,000 | 350,000 | 2 | 0 | 1 | 1 | 0 | 2.1 |
| Sylvia melanocephala | Passerine | Sedentary | Least concern | Flapping | 14 | 1,400,000 | 4,000,000 | 1 | 0 | 0 | 0 | 1 | 2.2 |
| Sylvia undata | Passerine | Sedentary | Near threatened | Flapping | 14 | 2,300,000 | 2,500,000 | 3 | 2 | 1 | 0 | 0 | 2.3 |
| Turdus iliacus | Passerine | Partial migration | Near threatened | Flapping | 21 | 0 | 6,000,000 | 1 | 0 | 0 | 0 | 1 | 0.9 |
| Turdus merula | Passerine | Partial migration | Least concern | Flapping | 26 | 4,100,000 | 52,000,000 | 4 | 1 | 0 | 2 | 1 | 4.8 |
| Turdus philomelos | Passerine | Partial migration | Least concern | Flapping | 21 | 300,000 | 18,000,000 | 4 | 1 | 1 | 1 | 1 | 3.6 |
| Turdus viscivorus | Passerine | Partial migration | Least concern | Flapping | 28 | 550,000 | 2,800,000 | 2 | 1 | 0 | 1 | 0 | 3.8 |
| Upupa epops | Upupiformes | Aestival | Least concern | Flapping | 27 | 600,000 | 1,100,000 | 1 | 0 | 0 | 0 | 1 | 2.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balmori-de la Puente, A.; Balmori, A. Flight Type and Seasonal Movements Are Important Predictors for Avian Collisions in Wind Farms. Birds 2023, 4, 85-100. https://doi.org/10.3390/birds4010007
Balmori-de la Puente A, Balmori A. Flight Type and Seasonal Movements Are Important Predictors for Avian Collisions in Wind Farms. Birds. 2023; 4(1):85-100. https://doi.org/10.3390/birds4010007
Chicago/Turabian StyleBalmori-de la Puente, Alfonso, and Alfonso Balmori. 2023. "Flight Type and Seasonal Movements Are Important Predictors for Avian Collisions in Wind Farms" Birds 4, no. 1: 85-100. https://doi.org/10.3390/birds4010007