
Respiratory Infection by Cyathostoma (Hovorkonema) americana in a Population of Burrowing Owls (Athene cunicularia)—A Potential Case of Zoo–Wildlife Cross-Transmission

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
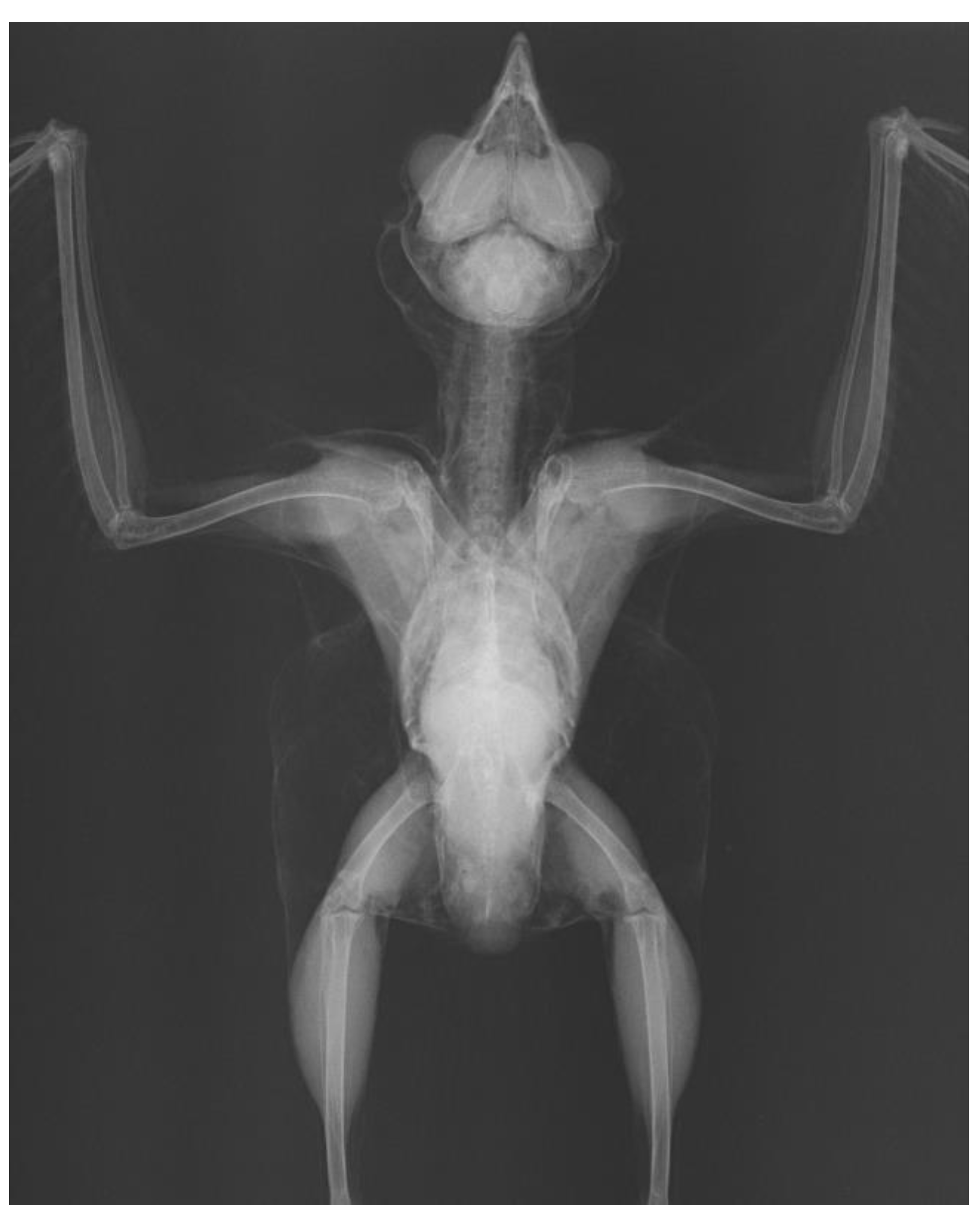
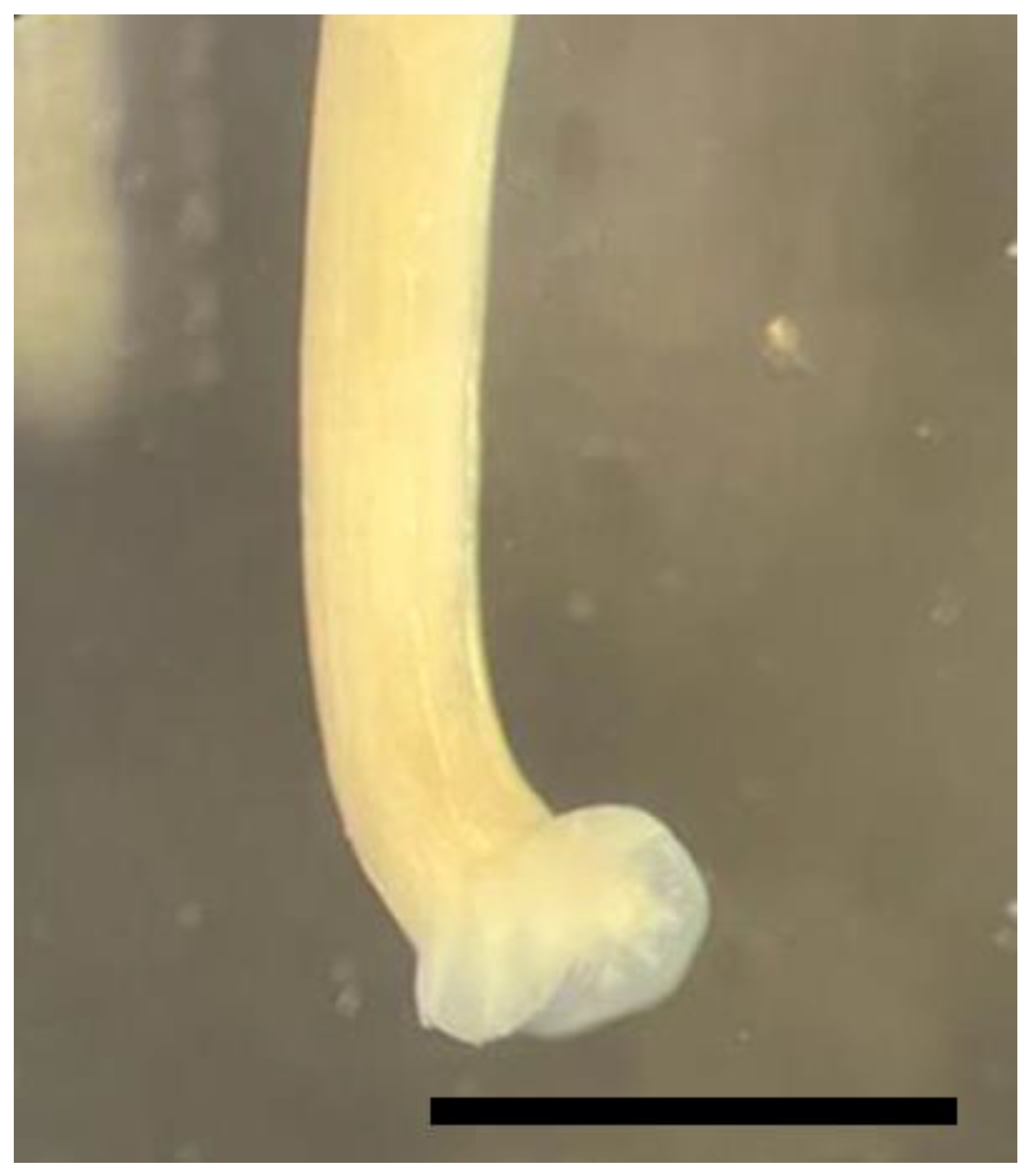
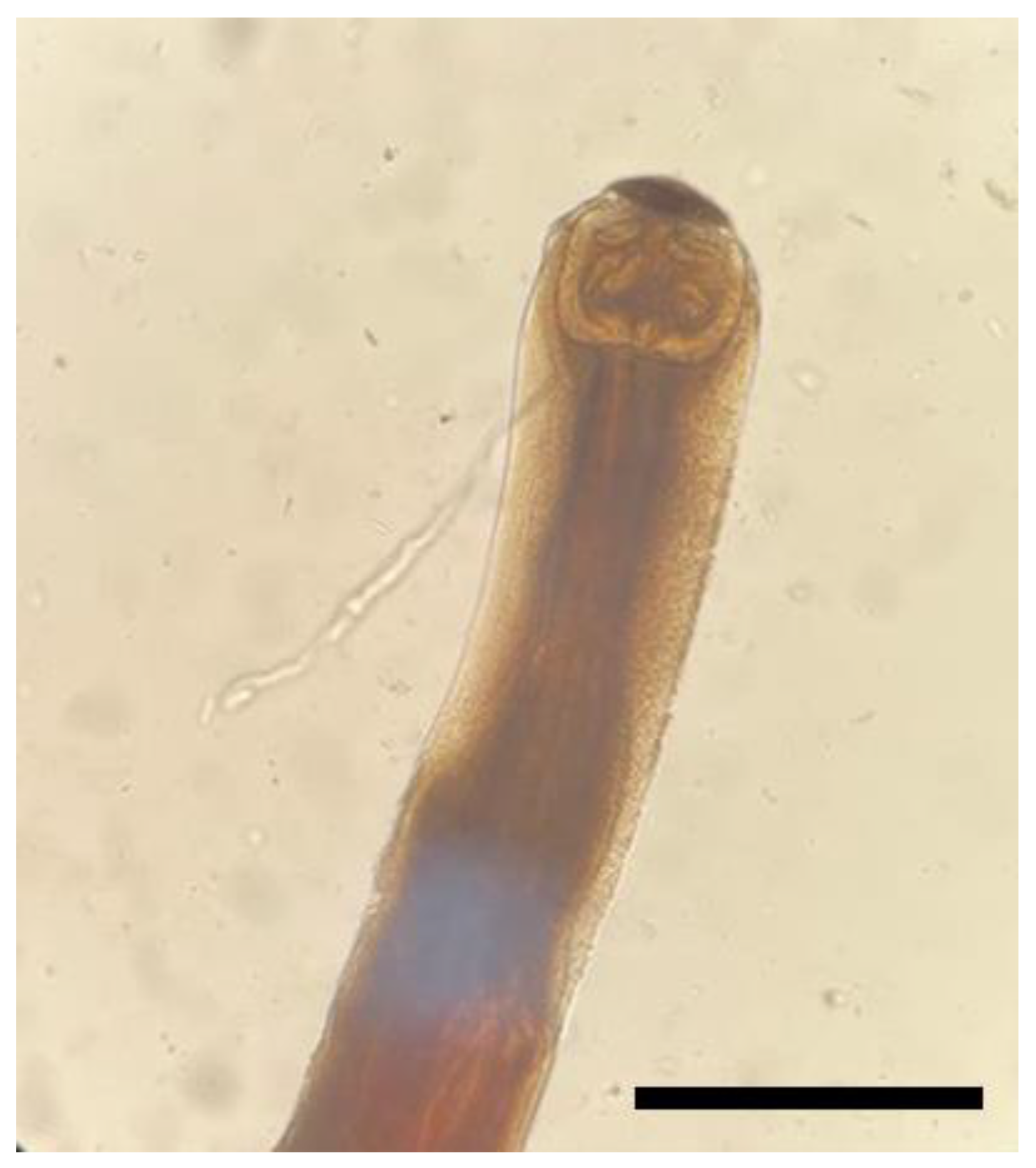
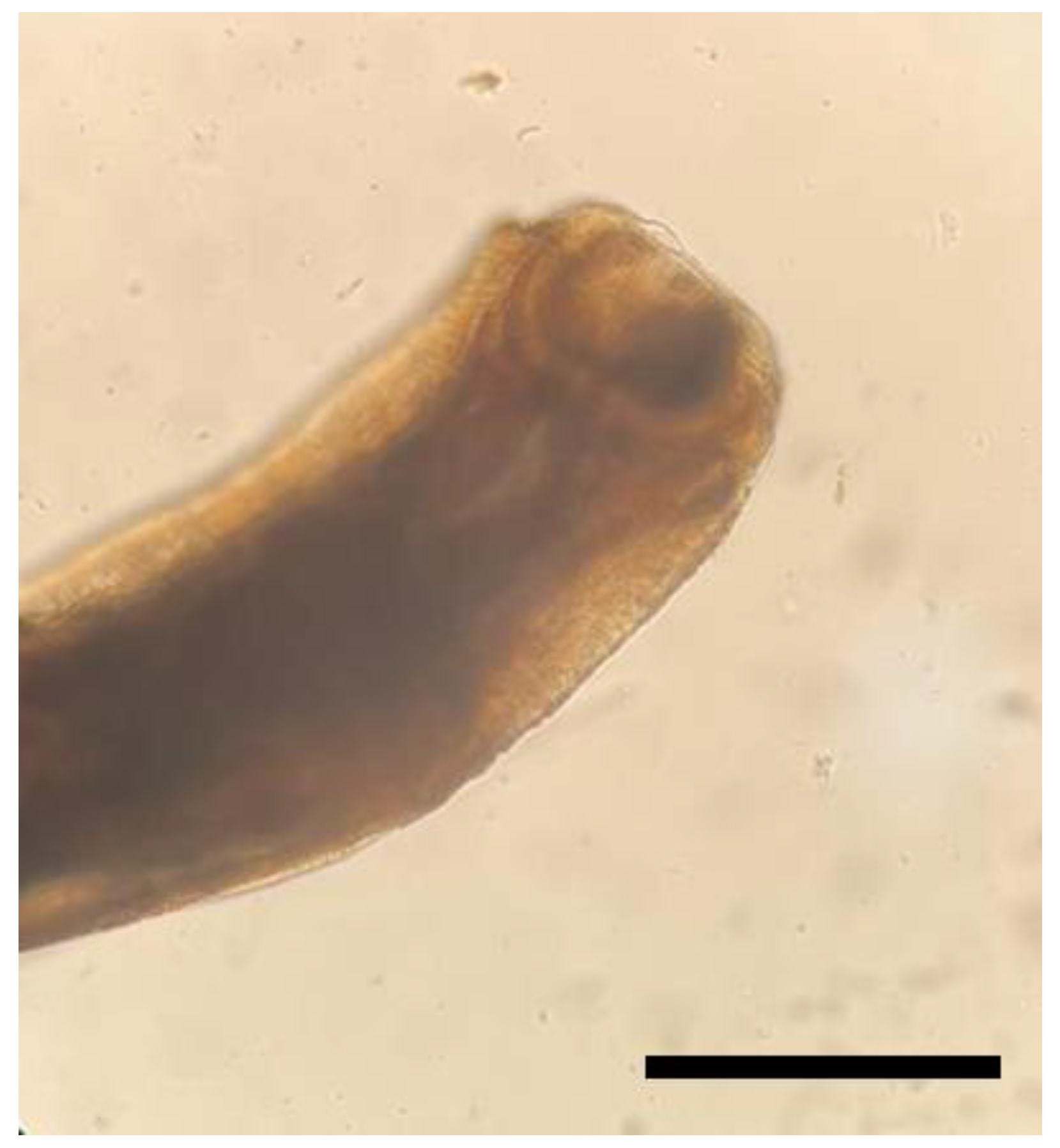
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The IUCN Red List of Threatened Species Home Page. BirdLife International 2016, Athene cunicularia. Available online: https://www.iucnredlist.org/species/pdf/93227732 (accessed on 6 July 2023).
- Rosenberg, K.V.; Kennedy, J.A.; Dettmers, R.; Ford, R.P.; Reynolds, D.; Alexander, J.D.; Beardmore, C.J.; Blancher, P.J.; Bogart, R.E.; Butcher, G.S.; et al. Partners in Flight Landbird Conservation Plan: 2016 Revision for Canada and Continental United States; Bogart, R.E., Ed.; Partners in Flight. 2016, pp. 1–105. Available online: https://partnersinflight.org/resources/the-plan/ (accessed on 27 November 2023).
- Brandes, S. Athene cunicularia, Animal Diversity Web. Available online: http://animaldiversity.org/accounts/Athene_cunicularia/ (accessed on 12 July 2023).
- Haug, E.; Oliphant, L. Movements, activity patterns, and habitat use of Burrowing Owls in Saskatchewan. J. Wildl. Manag. 1990, 54, 27–35. [Google Scholar] [CrossRef]
- Sarno, R.J.; Nixon, P.A.; Mealey, B.K.; Concoby, R.E.; Mrykalo, R.J.; Grigione, M.M. Suitability of translocation sites for Florida burrowing owls: Prey availability and diet. Southeast. Nat. 2012, 11, 755–764. [Google Scholar] [CrossRef]
- Trulio, L.; Higgins, P. The diet of western burrowing owls in an urban landscape. West. N. Am. Nat. 2012, 73, 348–356. [Google Scholar] [CrossRef]
- BBL—Longevity Records of North American Birds. Available online: http://genomics.senescence.info/species/biblio.php?id=427 (accessed on 23 October 2014).
- Kanarek, G.; Zaleśny, G.; Sitko, J.; Blanco, A.I. Taxonomic status of Cyathostoma nematodes (Nematoda: Syngaminae) parasitizing respiratory tracts of birds of prey and owls in Europe and North America: How many species are there? Helminthologia 2016, 53, 47–54. [Google Scholar] [CrossRef]
- Fernando, M.A.; Barta, J.R. Tracheal Worms. In Parasitic Diseases of Wild Birds, 1st ed.; Atkinson, C.T., Thomas, N.J., Hunter, D.B., Eds.; John Wiley & Sons, Ltd.: Ames, IA, USA, 2008; pp. 343–354. ISBN 978-0-813-82081-1. [Google Scholar]
- Hunter, D.B.; McKeever, K.; Bartlett, C. Cyathostoma Infections in Screech Owls, Saw-whet Owls, and Burrowing Owls in Southern Ontario. In Raptor Biomedicine, 1st ed.; Redig, P.T., Cooper, J.E., Remple, J.D., Hunder, D.B., Eds.; University of Minnesota Press: Minneapolis, MN, USA, 1993; pp. 54–56. ISBN 978-0816622191. [Google Scholar]
- Krone, O.; Friedrich, D.; Honisch, M. Specific status and pathogenicity of syngamid nematodes in bird species (Ciconiformes, Falconiformes, Gruiformes) from Germany. J. Helminthol. 2007, 81, 67–73. [Google Scholar] [CrossRef]
- Kanarek, G.; Horne, E.C.; Zaleśny, G. Cyathostoma (Cyathostoma) phenisci Baudet, 1937 (Nematoda: Syngamidae), a parasite of respiratory tract of African penguin Spheniscus demersus: Morphological and molecular characterization with some ecological and veterinary notes. Parasitol. Int. 2013, 62, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, M.; Mikaelian, I.; Sterner, M.; Villeneuve, A.; Fitzgerald, G.; McLaughlin, J.; Lair, S.; Martineau, D. Respiratory Nematodiases in Raptors in Quebec. J. Wildl. Dis. 1999, 35, 375–380. [Google Scholar] [CrossRef]
- Vaughan-Higgins, R.; Murphy, S.; Carter, I.; Pocknell, A.; Harris, E.; Sainsbury, A.W. Fatal epicarditis in a hen harrier (Circus cyaneus) a red-listed bird of high conservation concern in Britain associated with Cyathostoma species and Escherichia coli infection. Vet. Rec. 2013, 173, 477. [Google Scholar] [CrossRef]
- Lichenfels, J.R. Keys to the genera of the superfamily Strongyloidea. In CIH Keys to the Nematode Parasites of Vertebrates; Anderson, R.C., Chabaud, A.G., Willmott, S., Eds.; Commonwealth Institute of Helminthology: Farnham Royal, UK, 1980; Volume 7, 41p. [Google Scholar]
- Chapin, E.A. Review of the nematode genera Syngamus Sieb. and Cyathostoma E. Blanch. J. Agric. Res. 1925, 30, 557–570. [Google Scholar]
- Borgsteede, F.H.; Okulewicz, A.; Zoun, P.M.; Okulewicz, J. The helminth fauna of birds of prey (Accipitriformes, Falconiformes and Strigiformes) in the Netherlands. Acta Parasitol. 2003, 48, 200–207. [Google Scholar]
- Stiko, J.; Okulewicz, A. Checklist of the Nematodes in Birds in the Czech Republic and the Slovak Republic; Comenius Museum: Přerov, Czech Republic, 2010; 104p, ISBN 9788090371392. [Google Scholar]
- Kells, N.J. Review: The Five Domains model and promoting positive welfare in pigs. Animals 2022, 16, 100378. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human–Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef] [PubMed]
- Tallo-Parra, O.; Salas, M.; Manteca, X. Zoo Animal Welfare Assessment: Where Do We Stand? Animals 2023, 13, 1966. [Google Scholar] [CrossRef] [PubMed]
- Panayotova-Pencheva, M.S. Parasites in Captive Animals: A Review of Studies in Some European Zoos. Zool. Gart. 2013, 82, 60–71. [Google Scholar] [CrossRef]
- Papini, R.; Girivetto, M.; Marangi, M.; Mancianti, F.; Giangaspero, A. Endoparasite Infections in Pet and Zoo Birds in Italy. Sci. World J. 2012, 2012, 253127. [Google Scholar] [CrossRef] [PubMed]
- Mellen, J.; MacPhee, M.S. Philosophy of Environmental Enrichment: Past, Present, and Future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Carrera-Játiva, P.D.; Morgan, E.R.; Barrows, M.; Wronski, T. Gastrointestinal parasites in captive and free-ranging birds and potential cross-transmission in a zoo environment. J. Zoo Wildl. Med. 2018, 49, 116–128. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Barnabe, A.S.; Federsoni, I.P.; Ferraz, R.R.; Marco, R.M.; Garcia, I.P. Efficacy of parasitological diagnosis methods in wild animals kept in captivity. Arq. Do Inst. Biol. 2017, 84, e0502016. [Google Scholar] [CrossRef]
- Crosta, L. Respiratory Diseases of Parrots: Anatomy, Physiology, Diagnosis and Treatment. Vet. Clin. Exot. Anim. Pract. 2021, 24, 397–418. [Google Scholar] [CrossRef]
- Forbes, N.A. Raptors: Parasitic disease. In BSAVA Manual of Raptors, Pigeons and Passerine Birds, 1st ed.; Chitty, J., Lierz, M., Eds.; British Small Animal Veterinary Association: Gloucester, UK, 2008; Chapter 19; pp. 205–206. ISBN 978-1-905319-04-6. [Google Scholar]
- Lierz, M.; Schuster, R.; Ehrlein, J.; Gobel, T. Findings of Hovorkonema variegatum in northern goshawk (Accipiter gentilis). Kleintierpraxis 1998, 43, 43–46. [Google Scholar]
- Mitchell, E.; Johns, J. Avian Hematology and Related Disorders. Vet. Clin. Exot. Anim. Pract. 2008, 11, 501–522. [Google Scholar] [CrossRef] [PubMed]
- Bowles, H.L.; Odberg, E.; Harrison, G.J.; Kottwitz, J.J. Surgical resolution of soft tissue disorders. In Clinical Avian Medicine, 2nd ed.; Harrison, G.J., Lightfoot, T.L., Eds.; Spix Publishing, Inc.: Palm Beach, FL, USA, 2006; Volume 2, p. 794. ISBN 00-9754994-0-8. [Google Scholar]
- Doneley, B.; Harrison, G.J.; Lightfoot, T.L. Maximizing Information from the Physical Examination. In Clinical Avian Medicine, 2nd ed.; Harrison, G.J., Lightfoot, T.L., Eds.; Spix Publishing, Inc.: Palm Beach, FL, USA, 2006; Volume 1, p. 169. ISBN 00-9754994-0-8. [Google Scholar]
- Lee, S.-Y.; Kim, H.-J.; Kim, J.-W. Deflation Treatment for Subcutaneous Emphysema in a Goffin Cockatoo (Cacatua goffini). J. Vet. Clin. 2011, 28, 519–521. [Google Scholar]
- Vogel, H. About biology and morphology of Cyathostoma variegatum (Creplin, 1849) [Syngamus variegatum]. Z. Infekt.-Skrankh 1928, 34, 97–117. [Google Scholar]
- Ali, M.M. A review and revision of the subfamily Cyathostominae Nicoll, 1927 (Nematoda, Syngamidae). Acta Parasitol. Pol. 1970, 17, 237–246. [Google Scholar]
- Ryzhikov, K.M. The biology of a nematode Cyathostoma bronchialis (Syngamidae: Strongylata). Helminthologia 1980, 17, 241–244. [Google Scholar]
- Hartwich, G.; Nematoda, I.I. Strongylida: Strongyloidea und Ancylostomatoidea. In The Wildlife of Germany; Gustav Fisher Verlag: Jena, Germany; Frankfurt, Germany, 1994; p. 157. [Google Scholar]
- Kanarek, G. The occurrence of Cyathostoma (Cyathostoma) microspiculum (Skrjabin, 1913) (Nematoda: Syngamidae) in the great cormorant [Phalacrocorax carbo (L., 1758)] in north-eastern Poland. J. Helminthol. 2009, 83, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Okulewicz, A. Cyathostoma variegatum (Creplin, 1849) Chapin, 1925 and Cyathostoma lari Blanchard, 1849 (Nematoda, Syngamidae) in Poland. Wiad. Parazytol. 1984, 30, 53–56. [Google Scholar]
- Borgsteede, F.H.; Okulewicz, A. Justification of the species Cyathostoma (Hovorkonema) americana. Helminthologia 2001, 38, 115–121. [Google Scholar]
- Griffiths, H.J.; Leary, R.M.; Fenstermacher, R. A new record for gapeworm (Cyathostoma bronchialis) infection of domestic geese in North America. Am. J. Vet. Res. 1954, 15, 298–299. [Google Scholar]
- Ryžikov, K.M.; Zavadil, R. On the identity of species of the genus Cyathostoma collected from ostrich. Acta Univ. Agric. Silvic. Brno 1958, 2, 125–132. [Google Scholar]
- Hernandez-Rodriguez, S.; Calerto-Carretero, R.; Becerra-Martel, C.; Dominguez de Tena, M.; Moreno-Montañez, T.; Martinez-Gomez, F.; Barasona-Mata, J. Cyathostoma bronchialis (Mühling, 1884) Chapin, 1925 (Nematoda: Syngamidae) in Anser anser, first record in Spain. Rev. Iber. Parasitol. 1975, 35, 367–371. [Google Scholar]
- Carrera-Játiva, P.D.; Morgan, E.R.; Barrows, M.; Jiménez-Uzcátegui, G.; Roosevelt, J.; Tituaña, A. Free-ranging avifauna as a source of generalist parasites for captive birds in zoological settings: An overview of parasite records and potential for cross-transmission. J. Adv. Vet. Anim. Res. 2020, 7, 482–500. [Google Scholar] [CrossRef] [PubMed]
- The IUCN Red List of Threatened Species Home Page. BirdLife International 2016, Bubulcus ibis. Available online: https://www.iucnredlist.org/fr/species/22697109/155477521 (accessed on 27 November 2023).
- Omonona, A.O.; Ademola, I.O.; Ogunrotimi, M.O. Parasites of cattle egrets (Bubulcus ibis) and the associated haematological and biochemical changes. J. Environ. Ext 2014, 12, 31–40. [Google Scholar]
- Adediran, O.A.; Jubril, A.J.; Uwalaka, E.C.; Jarikre, T.A. Parasites, haematology and morphological changes in tissues of a migratory Bubulcus ibis. J. Zool. 2015, 4, 26–31. [Google Scholar] [CrossRef]
- Pires, H.; Matos, A.; Brida, T.; Lopes, F.; Alho, A.M.; Madeira de Carvalho, L.M. Parasitology survey in wild birds kept for rehabilitation in central eastern Portugal. In Proceedings of the10th International Symposium on Wild Fauna, Vila Real, Portugal, 21–23 September 2017; p. 92. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, G.N.; Cruz, J.T.; Pinto, M.; Leal, M.; Flanagan, C.; Urbani, N.; de Carvalho, L.M. Respiratory Infection by Cyathostoma (Hovorkonema) americana in a Population of Burrowing Owls (Athene cunicularia)—A Potential Case of Zoo–Wildlife Cross-Transmission. J. Zool. Bot. Gard. 2023, 4, 788-799. https://doi.org/10.3390/jzbg4040056
Marques GN, Cruz JT, Pinto M, Leal M, Flanagan C, Urbani N, de Carvalho LM. Respiratory Infection by Cyathostoma (Hovorkonema) americana in a Population of Burrowing Owls (Athene cunicularia)—A Potential Case of Zoo–Wildlife Cross-Transmission. Journal of Zoological and Botanical Gardens. 2023; 4(4):788-799. https://doi.org/10.3390/jzbg4040056
Chicago/Turabian StyleMarques, Gonçalo N., João T. Cruz, Matilde Pinto, Miriam Leal, Carla Flanagan, Nuno Urbani, and Luís Madeira de Carvalho. 2023. "Respiratory Infection by Cyathostoma (Hovorkonema) americana in a Population of Burrowing Owls (Athene cunicularia)—A Potential Case of Zoo–Wildlife Cross-Transmission" Journal of Zoological and Botanical Gardens 4, no. 4: 788-799. https://doi.org/10.3390/jzbg4040056