
Identifying Offspring Sex Ratio Skews in Zoological Facilities Using Large Historical Datasets

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
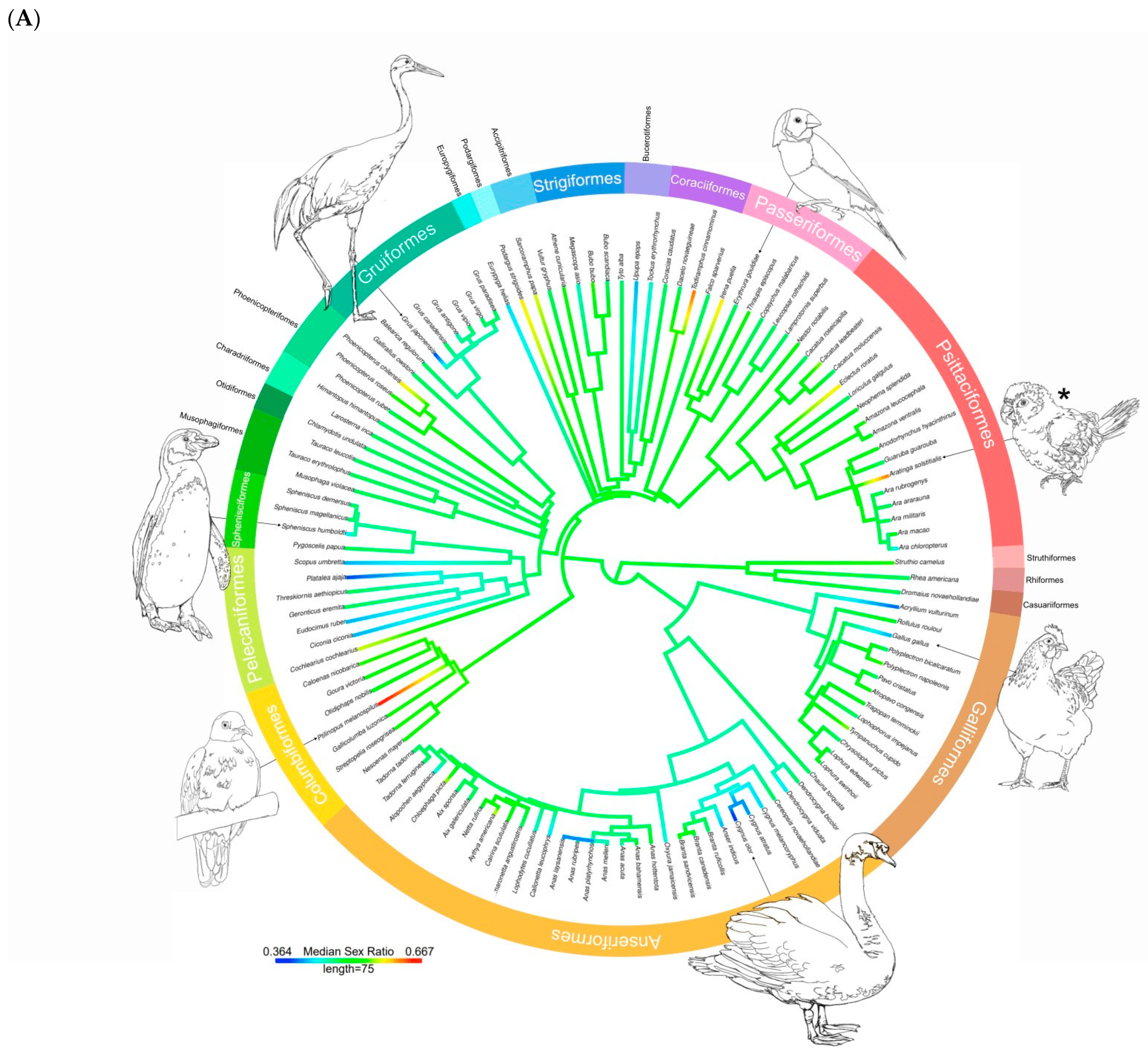
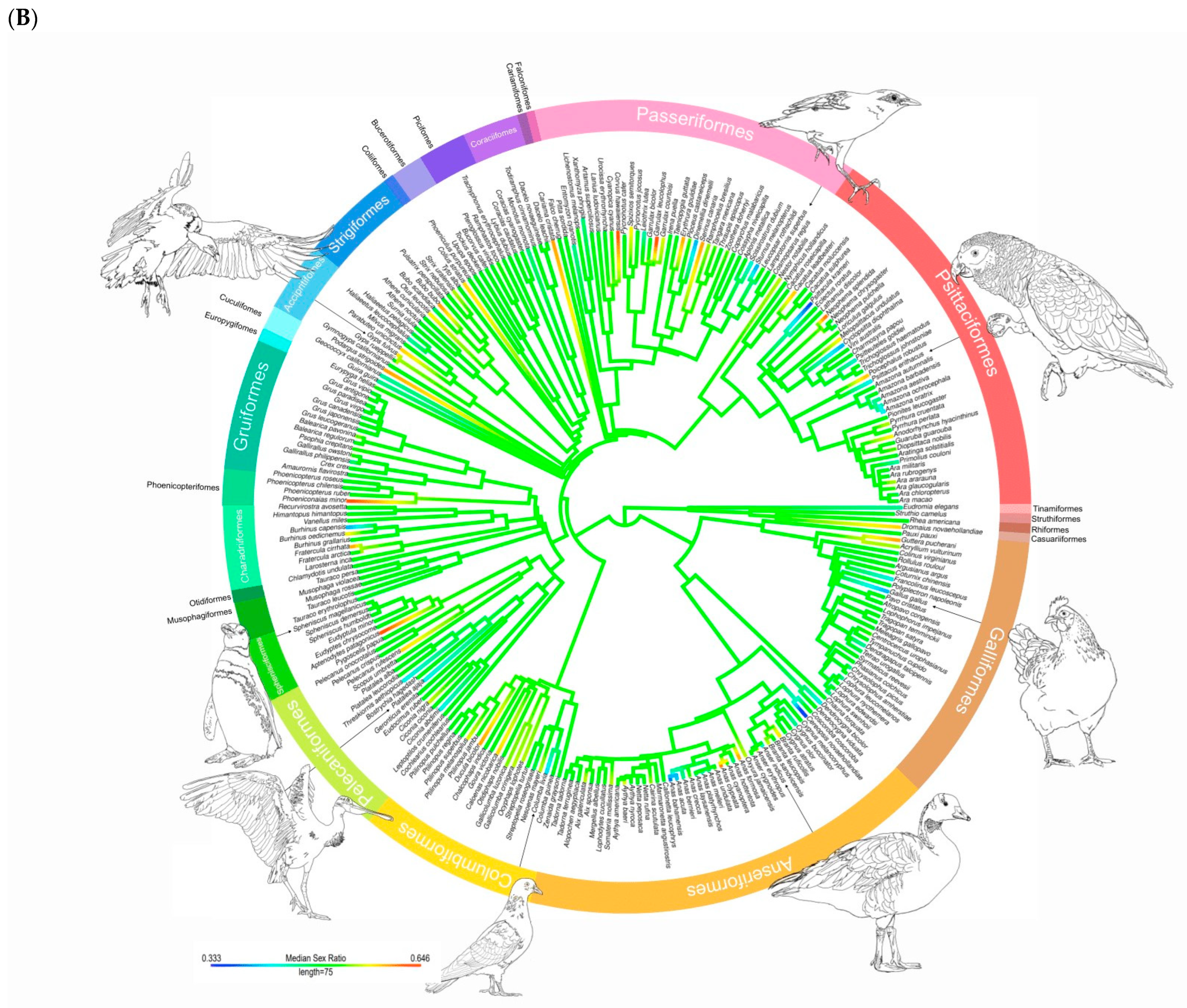
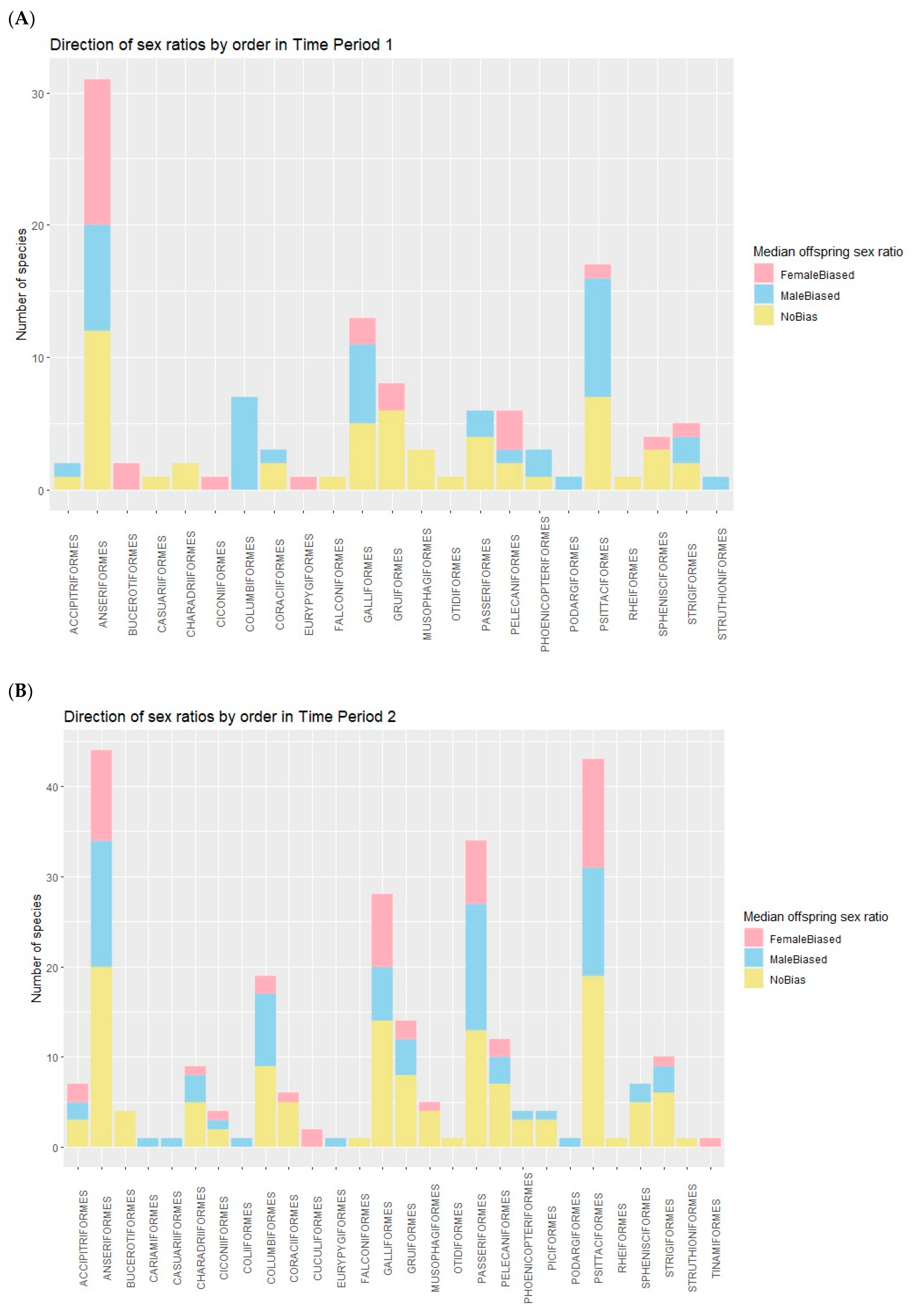
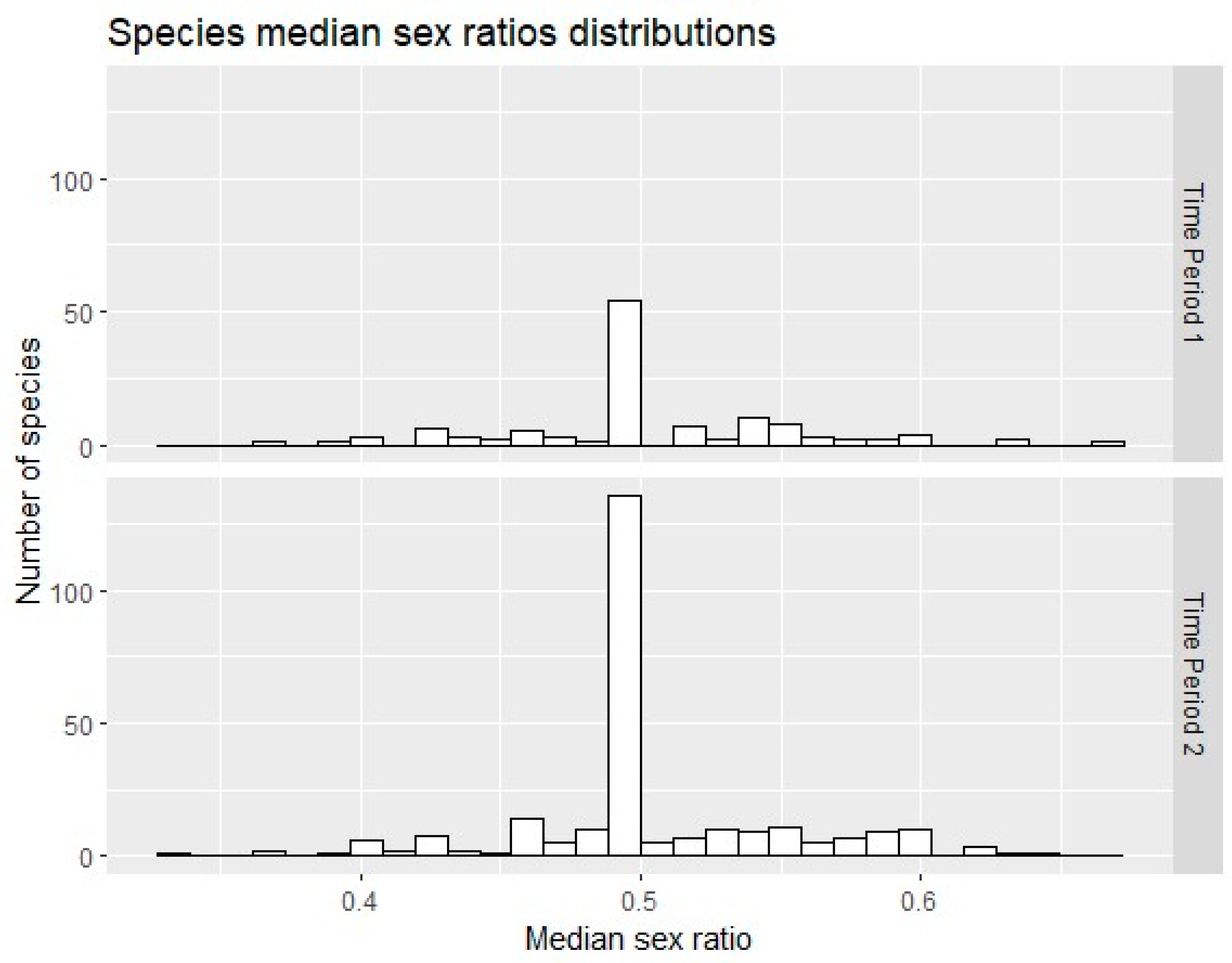
3. Results
3.1. Descriptive Statistics
3.2. Significance Testing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, R. The Genetical Theory of Natural Selection; Clarendon Press: Oxford, UK, 1930. [Google Scholar]
- Edwards, A.F.W.; Katlav, A.; Nguyen, D.T.; Cook, J.M.; Riegler, M.; Heine, K.B.; Shech, E.; Takacs, P.; Otto, P.A.; Stearns, S.C.; et al. Natural Selection and the Sex Ratio: Fisher’s Sources. Am. Nat. 1998, 151, 564. [Google Scholar] [CrossRef]
- Brand, J.N.; Harmon, L.J.; Schärer, L. Mating behavior and reproductive morphology predict macroevolution of sex allocation in hermaphroditic flatworms. BMC Biol. 2022, 20, 35. [Google Scholar] [CrossRef]
- Navara, K.J. Hormone-Mediated Adjustment of Sex Ratio in Vertebrates. Integr. Comp. Biol. 2013, 53, 877–887. [Google Scholar] [CrossRef]
- Sinclair, J.P.; Emlen, J.; Freeman, D.C. Biased Sex Ratios in Plants: Theory and Trends. Bot. Rev. 2012, 78, 63–86. [Google Scholar] [CrossRef]
- Kalmbach, E.; Nager, R.G.; Griffiths, R.; Furness, R.W. Increased reproductive effort results in male-biased offspring sex ratio: An experimental study in a species with reversed sexual size dimorphism. Proc. R. Soc. B Biol. Sci. 2001, 268, 2175–2179. [Google Scholar] [CrossRef]
- Clark, A.B. Sex Ratio and Local Resource Competition in a Prosimian Primate. Science 1978, 201, 163–165. [Google Scholar] [CrossRef]
- Silk, J.B. Local Resource Competition and Facultative Adjustment of Sex Ratios in Relation to Competitive Abilities. Am. Nat. 1983, 121, 56–66. [Google Scholar] [CrossRef]
- Komdeur, J.; Daan, S.; Tinbergen, J.; Mateman, C. Extreme adaptive modification in sex ratio of the Seychelles warbler’s eggs. Nature 1997, 385, 522–525. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H.; Iason, G.R. Sex Ratio Variation in Mammals. Q. Rev. Biol. 1986, 61, 339–374. [Google Scholar] [CrossRef]
- Emlen, S.T.; Merritt Emlen, J.; Levin, S.A. Sex-Ratio Selection in Species with Helpers-At-The-Nest. Am. Nat. 1986, 127, 1–8. Available online: https://about.jstor.org/terms (accessed on 10 December 2021). [CrossRef]
- Weladji, R.B.; Holand, Ø.; Yoccoz, N.G.; Lenvik, D. Maternal age and offspring sex ratio variation in reindeer (Rangifer tarandus). Ann. Zool. Fennici. 2003, 40, 357–363. [Google Scholar]
- Blank, J.L.; Nolan, V. Offspring sex ratio in red-winged blackbirds is dependent on maternal age. Proc. Natl. Acad. Sci. USA 1983, 80, 6141–6145. [Google Scholar] [CrossRef] [PubMed]
- Côté, S.D.; Festa-Bianchet, M. Offspring Sex Ratio in Relation to Maternal Age and Social Rank in Mountain Goats (Oreamnos americanus). Behav. Ecol. Sociobiol. 2001, 49, 260–265. Available online: https://www.jstor.org/stable/4601885 (accessed on 10 December 2021). [CrossRef]
- Aslam, M.A.; Groothuis, T.G.; Smits, M.A.; Woelders, H. Effect of Corticosterone and Hen Body Mass on Primary Sex Ratio in Laying Hen (Gallus gallus), Using Unincubated Eggs. Biol. Reprod. 2014, 90, 76. [Google Scholar] [CrossRef]
- Goerlich, V.C.; Dijkstra, C.; Boonekamp, J.J.; Groothuis, T.G.G. Change in Body Mass Can Overrule the Effects of Maternal Testosterone on Primary Offspring Sex Ratio of First Eggs in Homing Pigeons. Physiol. Biochem. Zool. 2010, 83, 490–500. [Google Scholar] [CrossRef]
- Parker, T.H. Maternal Condition, Reproductive Investment, and Offspring Sex Ratio in Captive Red Junglefowl (Gallus gallus). Ornithology 2002, 119, 840. [Google Scholar] [CrossRef]
- Nager, R.G.; Monaghan, P.; Griffiths, R.; Houston, D.C.; Dawson, R. Experimental demonstration that offspring sex ratio varies with maternal condition. Proc. Natl. Acad. Sci. USA 1999, 96, 570–573. Available online: www.pnas.org (accessed on 10 December 2021). [CrossRef]
- Clutton-Brock, T.H.; Albon, S.D.; Guinness, F.E. Maternal dominance, breeding success and birth sex ratios in red deer. Nature 1984, 308, 358–360. [Google Scholar] [CrossRef]
- Daan, S.; Dijkstra, C.; Weissing, F.J. An evolutionary explanation for seasonal trends in avian sex ratios. Behav. Ecol. 1996, 7, 426–430. [Google Scholar] [CrossRef]
- Cordero, P.J.; Viñuela, J.; Aparicio, J.M.; Veiga, J.P. Seasonal variation in sex ratio and sexual egg dimorphism favouring daughters in first clutches of the spotless starling. J. Evol. Biol. 2001, 14, 829–834. [Google Scholar] [CrossRef]
- Saalfeld, S.T.; Conway, W.C.; Haukos, D.A.; Johnson, W.P. Seasonal Variation in Offspring Sex Ratio in the Snowy Plover. West. N. Am. Nat. 2013, 73, 60–71. [Google Scholar] [CrossRef]
- Smallwood, P.D.; Smallwood, J.A. Seasonal shifts in sex ratios of fledgling American kestrels (Falco sparverius paulus): The Early Bird Hypothesis. Evol. Ecol. 1998, 12, 839–853. [Google Scholar] [CrossRef]
- Sequeira, R.; MacKauer, M. Seasonal Variation in Body Size and Offspring Sex Ratio in Field Populations of the Parasitoid Wasp, Aphidius ervi (Hymenoptera: Aphidiidae). Oikos 1993, 68, 340. [Google Scholar] [CrossRef]
- Barclay, R.M.R. Variable Variation: Annual and Seasonal Changes in Offspring Sex Ratio in a Bat. PLoS ONE 2012, 7, e36344. [Google Scholar] [CrossRef] [PubMed]
- Navara, K.J. The Bees Do It, but What About the Birds? Evidence for Sex Ratio Adjustment in Birds. In Choosing Sexes; Springer: Berlin/Heidelberg, Germany, 2018; pp. 71–97. [Google Scholar]
- Navara, K.J. Potential Mechanisms of Sex Ratio Adjustment in Birds. In Choosing Sexes; Springer: Berlin/Heidelberg, Germany, 2018; pp. 99–121. [Google Scholar]
- Navara, K.J. Hormones Rule the Roost: Hormonal Influences on Sex Ratio Adjustment in Birds and Mammals. In Choosing Sexes; Springer: Berlin/Heidelberg, Germany, 2018; pp. 123–154. [Google Scholar]
- Pierron, F.; Lorioux, S.; Héroin, D.; Daffe, G.; Etcheverria, B.; Cachot, J.; Morin, B.; Dufour, S.; Gonzalez, P. Transgenerational epigenetic sex determination: Environment experienced by female fish affects offspring sex ratio. Environ. Pollut. 2021, 277, 116864. [Google Scholar] [CrossRef]
- Frankham, R.; Wilcken, J. Does inbreeding distort sex-ratios? Conserv. Genet. 2006, 7, 879–893. [Google Scholar] [CrossRef]
- Love, O.P.; Chin, E.H.; Wynne-Edwards, K.E.; Williams, T.D. Stress Hormones: A Link between Maternal Condition and Sex-Biased Reproductive Investment. Am. Nat. 2005, 166, 751–766. [Google Scholar] [CrossRef]
- Gam, A.E.; Mendonça, M.T.; Navara, K.J. Acute corticosterone treatment prior to ovulation biases offspring sex ratios towards males in zebra finches Taeniopygia guttata. J. Avian Biol. 2011, 42, 253–258. [Google Scholar] [CrossRef]
- Henriksen, R.; Rettenbacher, S.; Groothuis, T.G. Maternal corticosterone elevation during egg formation in chickens (Gallus gallus domesticus) influences offspring traits, partly via prenatal undernutrition. Gen. Comp. Endocrinol. 2013, 191, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Pike, T.W.; Petrie, M. Experimental evidence that corticosterone affects offspring sex ratios in quail. Proc. R. Soc. B Biol. Sci. 2006, 273, 1093–1098. [Google Scholar] [CrossRef]
- Pinson, S.E.; Parr, C.M.; Wilson, J.L.; Navara, K.J. Acute Corticosterone Administration during Meiotic Segregation Stimulates Females to Produce More Male Offspring. Physiol. Biochem. Zool. 2011, 84, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Rutkowska, J.; Cichoń, M. Maternal testosterone affects the primary sex ratio and offspring survival in zebra finches. Anim. Behav. 2006, 71, 1283–1288. [Google Scholar] [CrossRef]
- Rubolini, D.; Romano, M.; Martinelli, R.; Saino, N. Effects of elevated yolk testosterone levels on survival, growth and immunity of male and female yellow-legged gull chicks. Behav. Ecol. Sociobiol. 2005, 59, 344–352. [Google Scholar] [CrossRef]
- Navara, K.J. Programming of offspring sex ratios by maternal stress in humans: Assessment of physiological mechanisms using a comparative approach. J. Comp. Physiol. B 2010, 180, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Firman, R.C. Exposure to high male density causes maternal stress and female-biased sex ratios in a mammal. Proc. R. Soc. B Biol. Sci. 2020, 287, 20192909. [Google Scholar] [CrossRef]
- Martin, M.S.; Owen, M.; Wintle, N.J.P.; Zhang, G.; Zhang, H.; Swaisgood, R.R. Stereotypic behaviour predicts reproductive performance and litter sex ratio in giant pandas. Sci. Rep. 2020, 10, 7263. [Google Scholar] [CrossRef]
- Bradbury, R.B.; Blakey, J.K. Diet, maternal condition, and offspring sex ratio in the zebra finch, Poephila guttata. Proc. R. Soc. B Biol. Sci. 1998, 265, 895–899. [Google Scholar] [CrossRef]
- Rutstein, A.N.; Slater, P.J.B.; Graves, J.A.; Rutstein, A.N.; Slater, P.J.B.; Graves, J.A. Diet quality and resource allocation in the zebra finch. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2004, 271 (Suppl. S5), S286–S289. [Google Scholar] [CrossRef]
- Benito, M.M.; Schielzeth, H.; González-Solís, J.; Becker, P.H. Sex ratio adjustments in common terns: Influence of mate condition and maternal experience. J. Avian Biol. 2013, 44, 179–188. [Google Scholar] [CrossRef]
- Svensson, E.; Nilsson, J.A. Mate Quality Affects Offspring Sex Ratio in Blue Tits. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1996, 263, 357–361. Available online: https://about.jstor.org/term (accessed on 10 December 2021).
- Kilner, R. Primary and secondary sex ratio manipulation by zebra finches. Anim. Behav. 1998, 56, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Hosey, G.R. How does the zoo environment affect the behaviour of captive primates? Appl. Anim. Behav. Sci. 2005, 90, 107–129. [Google Scholar] [CrossRef]
- Lees, C.M.; Wilcken, J. Sustaining the Ark: The challenges faced by zoos in maintaining viable populations. Int. Zoo Yearb. 2009, 43, 6–18. [Google Scholar] [CrossRef]
- Redford, K.H.; Jensen, D.B.; Breheny, J.J. Integrating the Captive and the Wild. Science 2012, 338, 1157–1158. [Google Scholar] [CrossRef] [PubMed]
- Bowkett, A.E. Recent Captive-Breeding Proposals and the Return of the Ark Concept to Global Species Conservation. Conserv. Biol. 2009, 23, 773–776. [Google Scholar] [CrossRef]
- Firman, R.C.; Ellis, C.M.; Thorn, S.; Mawson, P.R. Parental effects on offspring sex ratio in the Numbat (Myrmecobius fasciatus): Does captivity influence paternal sex allocation? J. Mammal. 2023, 104, 1036–1046. [Google Scholar] [CrossRef]
- Ward, S.J.; Sherwen, S.; Clark, F.E. Advances in Applied Zoo Animal Welfare Science. J. Appl. Anim. Welf. Sci. 2018, 21 (Suppl. S1), 23–33. [Google Scholar] [CrossRef]
- Chicago Zoological Society. Version 1.7. 2014. Species360 Zoological Information Management System. Available online: https://species360.org/ (accessed on 15 September 2023).
- Abdi, H. Holm’s Sequential Bonferroni Procedure. In Encyclopedia of Research Design; Salkind, N., Ed.; Sage Publication, Inc.: Sage Oaks, CA, USA, 2010; Available online: http://www.utd.edu/~herve (accessed on 1 September 2023).
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Nesterenko, O.; Kashentseva, T. Female-Biased Sex Ratio of Nestlings of Captive Red-Crowned Cranes in Russia. In Proceedings of the VII European Crane Conference: Breeding, Resting, Migration and Biology, Stralsund, Germany, 14–17 October 2010; Nowald, G., Weber, A., Fanke, J., Weinhardt, E., Donner, N., Eds.; Deutsches Meeresmuseum Stralsund: Stralsund, Germany, 2010; pp. 123–125. Available online: https://www.researchgate.net/publication/272492001 (accessed on 18 March 2023).
- Taylor, T.D.; Parkin, D.T. Sex ratios observed in 80 species of parrots. J. Zool. 2008, 276, 89–94. [Google Scholar] [CrossRef]
- Lehikoinen, A.; Öst, M.; Hollmén, T.; Kilpi, M. Does sex-specific duckling mortality contribute to male bias in adult common eiders? Condor 2008, 110, 574–578. [Google Scholar] [CrossRef]
- Eberhart-Phillips, L.J.; Küpper, C.; Miller, T.E.X.; Cruz-López, M.; Maher, K.H.; dos Remedios, N.; Stoffel, M.A.; Hoffman, J.I.; Krüger, O.; Székely, T. Sex-specific early survival drives adult sex ratio bias in snowy plovers and impacts mating system and population growth. Proc. Natl. Acad. Sci. USA 2017, 114, E5474–E5481. [Google Scholar] [CrossRef] [PubMed]
- Benito, M.M.; González-Solís, J. Sex ratio, sex-specific chick mortality and sexual size dimorphism in birds. J. Evol. Biol. 2007, 20, 1522–1530. [Google Scholar] [CrossRef] [PubMed]
- Maness, T.J.; Anderson, D.J. Predictors of Juvenile Survival in Birds. Ornithol. Monogr. 2013, 78, 1–55. [Google Scholar] [CrossRef]
- Dutton, C.J.; Tieber, A. A modified protocol for sex identification of in ovo avian embryos and its application as a management tool for endangered species conservation programs. J. Zoo Wildl. Med. 2001, 32, 176–180. [Google Scholar] [CrossRef]
- Jensen, T.; Mace, M.; Durrant, B. Sexing of mid-incubation avian embryos as a management tool for zoological breeding programs. Zoo Biol. 2012, 31, 694–704. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado, J.M.; Miller, L.J. Identifying Offspring Sex Ratio Skews in Zoological Facilities Using Large Historical Datasets. J. Zool. Bot. Gard. 2023, 4, 680-691. https://doi.org/10.3390/jzbg4040048
Machado JM, Miller LJ. Identifying Offspring Sex Ratio Skews in Zoological Facilities Using Large Historical Datasets. Journal of Zoological and Botanical Gardens. 2023; 4(4):680-691. https://doi.org/10.3390/jzbg4040048
Chicago/Turabian StyleMachado, Julia M., and Lance J. Miller. 2023. "Identifying Offspring Sex Ratio Skews in Zoological Facilities Using Large Historical Datasets" Journal of Zoological and Botanical Gardens 4, no. 4: 680-691. https://doi.org/10.3390/jzbg4040048