
Overlooked and Under-Studied: A Review of Evidence-Based Enrichment in Varanidae



{kind=link}
Abstract
:1. Introduction
2. Reptile Enrichment: What Do We Know?
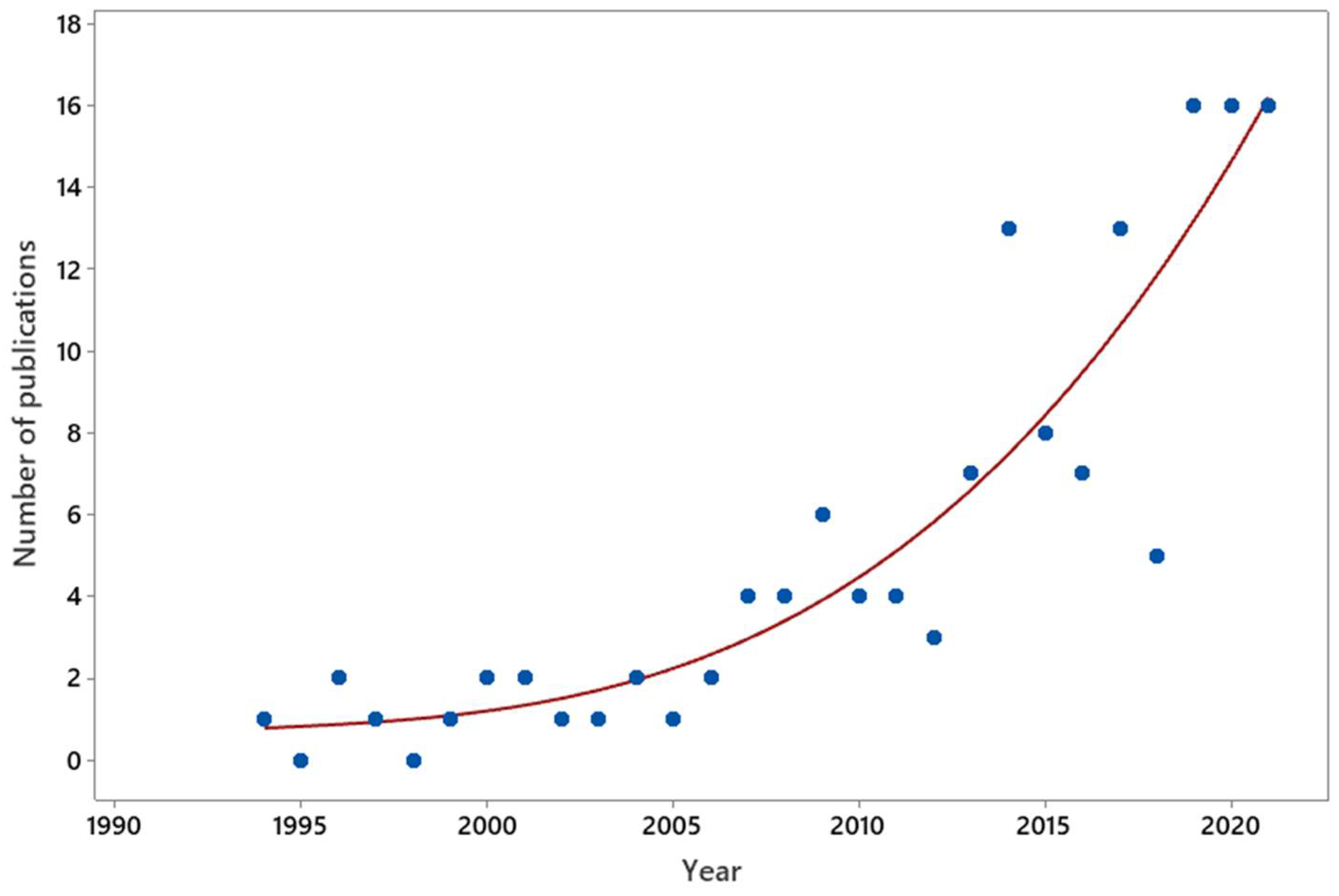
2.1. Left in the Cold
2.2. Do Reptiles Benefit?
2.3. The Necessity of Evaluation
3. Is Enrichment Vital for Varanids?
3.1. A Brief Background of Varanidae
3.2. Cognitive Abilities
3.3. Play Behaviour
4. Future Directions for Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rees, P.A. An Introduction to Zoo Biology and Management; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Markowitz, H. Engineering Environments for Behavioral Opportunities in the Zoo. Behav. Anal. 1978, 1, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.J.; Martin, A.L. Animal Training, Environmental Enrichment, and Animal Welfare: A History of Behavior Analysis in Zoos. J. Zool. Bot. Gard. 2021, 2, 531. [Google Scholar] [CrossRef]
- Clark, F.E. Cognitive Enrichment and Welfare: Current Approaches and Future Directions. Anim. Behav. Cognit. 2017, 4, 52–71. [Google Scholar] [CrossRef] [Green Version]
- Riley, L.M.; Rose, P.E. Concepts, Applications, Uses and Evaluation of Environmental Enrichment: Perceptions of Zoo Professionals. J. Zoo Aquar. Res. 2020, 8, 18–28. [Google Scholar]
- Banton-Jones, K. Why Enrichment Isn’t an Extra. Available online: www.wildenrichment.com (accessed on 11 July 2021).
- Shepherdson, D.J. Tracing the Path of Environmental Enrichment in Zoos. In Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, D., Mellen, J.D., Hutchins, M., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 1–12. [Google Scholar]
- Buchanan-Smith, H.M. Environmental Enrichment for Primates in Laboratories. Adv. Sci. Res. 2011, 5, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Bloomsmith, M.A.; Brent, L.Y.; Schapiro, S.J. Guidelines for Developing and Managing an Environmental Enrichment Program for Nonhuman Primates. Lab. Anim. Sci. 1991, 41, 372–377. [Google Scholar] [PubMed]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty Years Later: Enrichment Practices for Captive Mammals. Zoo Biol. 2010, 29, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Skibiel, A.L.; Trevino, H.S.; Naugher, K. Comparison of Several Types of Enrichment for Captive Felids. Zoo Biol. 2007, 26, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Mellen, J.; Sevenich MacPhee, M. Philosophy of Environmental Enrichment: Past, Present, and Future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Mason, G.; Clubb, R.; Latham, N.; Vickery, S. Why and How Should We Use Environmental Enrichment to Tackle Stereotypic Behaviour? Appl. Anim. Behav. Sci. 2007, 102, 163–188. [Google Scholar] [CrossRef] [Green Version]
- Melfi, V.A. There Are Big Gaps in Our Knowledge, and Thus Approach, to Zoo Animal Welfare: A Case for Evidence-Based Zoo Animal Management. Zoo Biol. 2009, 28, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Brereton, J.E.; Rowden, L.J.; de Figueiredo, R.L.; Riley, L.M. What’s New from the Zoo? An Analysis of Ten Years of Zoo-Themed Research Output. Palgrave Commun 2019, 5, 1–10. [Google Scholar]
- Swaisgood, R.R. Current Status and Future Directions of Applied Behavioral Research for Animal Welfare and Conservation. Appl. Anim. Behav. Sci. 2007, 102, 139–162. [Google Scholar] [CrossRef]
- Eagan, T. Evaluation of Enrichment for Reptiles in Zoos. J. Appl. Anim. Welf. Sci. 2019, 22, 69–77. [Google Scholar] [CrossRef]
- De Azevedo, C.S.; Cipreste, C.F.; Young, R.J. Environmental Enrichment: A GAP Analysis. Appl. Anim. Behav. Sci. 2007, 102, 329–343. [Google Scholar] [CrossRef]
- Maple, T.L.; Perdue, B.M. Environmental Enrichment. In Zoo Animal Welfare; Springer: Berlin/Heidelberg, Germany, 2013; pp. 95–117. [Google Scholar]
- Alligood, C.; Leighty, K. Putting the “E” in SPIDER: Evolving Trends in the Evaluation of Environmental Enrichment Efficacy in Zoological Settings. Anim. Behav. Cogn. 2015, 2, 200–217. [Google Scholar] [CrossRef]
- Case, B.C.; Lewbart, G.A.; Doerr, P.D. The Physiological and Behavioural Impacts of and Preference for an Enriched Environment in the Eastern Box Turtle (Terrapene carolina carolina). Appl. Anim. Behav. Sci. 2005, 92, 353–365. [Google Scholar] [CrossRef]
- Doody, J.S.; Burghardt, G.M.; Dinets, V. Breaking the Social–Non-Social Dichotomy: A Role for Reptiles in Vertebrate Social Behavior Research? Ethology 2013, 119, 95–103. [Google Scholar] [CrossRef]
- Learmonth, M.J. The Matter of Non-Avian Reptile Sentience, and Why It “Matters” to Them: A Conceptual, Ethical and Scientific Review. Animals 2020, 10, 901. [Google Scholar] [CrossRef]
- Warwick, C.; Arena, P.; Lindley, S.; Jessop, M.; Steedman, C. Assessing Reptile Welfare Using Behavioural Criteria. InPractice 2013, 35, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Roth, T.C.; Krochmal, A.R.; LaDage, L.D. Reptilian Cognition: A More Complex Picture via Integration of Neurological Mechanisms, Behavioral Constraints, and Evolutionary Context. BioEssays 2019, 41, 1900033. [Google Scholar] [CrossRef]
- Burghardt, G.M.; Ward, B.; Rosscoe, R. Problem of Reptile Play: Environmental Enrichment and Play Behavior in a Captive Nile Soft-Shelled Turtle, Trionyx Triunguis. Zoo Biol. 1996, 15, 223–238. [Google Scholar] [CrossRef]
- Londoño, C.; Bartolomé, A.; Carazo, P.; Font, E. Chemosensory Enrichment as a Simple and Effective Way to Improve the Welfare of Captive Lizards. Ethology 2018, 124, 674–683. [Google Scholar] [CrossRef]
- Almli, L.M.; Burghardt, G.M. Environmental Enrichment Alters the Behavioral Profile of Ratsnakes (Elaphe). J. Appl. Anim. Welf. Sci. 2006, 9, 85–109. [Google Scholar] [CrossRef]
- Therrien, C.L.; Gaster, L.; Cunningham-Smith, P.; Manire, C.A. Experimental Evaluation of Environmental Enrichment of Sea Turtles. Zoo Biol. 2007, 26, 407–416. [Google Scholar] [CrossRef]
- Passos, L.F.; Garcia, G.; Young, R. How Does Captivity Affect Skin Colour Reflectance of Golden Mantella Frogs? Herpetol. J. 2020, 30, 13–19. [Google Scholar] [CrossRef]
- Bashaw, M.J.; Gibson, M.D.; Schowe, D.M.; Kucher, A.S. Does Enrichment Improve Reptile Welfare? Leopard Geckos (Eublepharis macularius) Respond to Five Types of Environmental Enrichment. Appl. Anim. Behav. Sci. 2016, 184, 150–160. [Google Scholar]
- Burghardt, G.M. Environmental Enrichment and Cognitive Complexity in Reptiles and Amphibians: Concepts, Review, and Implications for Captive Populations. Appl. Anim. Behav. Sci. 2013, 147, 286–298. [Google Scholar] [CrossRef]
- Rose, P.; Evans, C.; Coffin, R.; Miller, R.; Nash, S. Using Student-Centred Research to Evidence-Base Exhibition of Reptiles and Amphibians: Three Species-Specific Case Studies. J. Zoo Aquar. Res. 2014, 2, 25–32. [Google Scholar]
- Hoehfurtner, T.; Wilkinson, A.; Nagabaskaran, G.; Burman, O.H.P. Does the Provision of Environmental Enrichment Affect the Behaviour and Welfare of Captive Snakes? Appl. Anim. Behav. Sci. 2021, 239, 105324. [Google Scholar] [CrossRef]
- Bannister, C.C.; Thomson, A.J.C.; Cuculescu-Santana, M. Can Colored Object Enrichment Reduce the Escape Behavior of Captive Freshwater Turtles? Zoo Biol. 2021, 40, 160–168. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific Approaches to Enrichment and Stereotypies in Zoo Animals: What’s Been Done and Where Should We Go Next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Saudargas, R.A.; Drummer, L.C. Single Subject (Small N) Research Designs and Zoo Research; Wiley: Hoboken, NJ, USA, 1996. [Google Scholar]
- Burghardt, G.M. A Brief Glimpse at the Long Evolutionary History of Play. Anim. Behav. Cogn. 2014, 1, 90–98. [Google Scholar] [CrossRef]
- Januszczak, I.S.; Bryant, Z.; Tapley, B.; Gill, I.; Harding, L.; Michaels, C.J. s Behavioural Enrichment Always a Success? Comparing Food Presentation Strategies in an Insectivorous Lizard (Plica plica). Appl. Anim. Behav. Sci. 2016, 183, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Kanaan, V.T.; Hötzel, M.J. Does Environmental Enrichment Really Matter? A Case Study Using the Eastern Fence Lizard, Sceloporus Undulatus. Appl. Anim. Behav. Sci. 2011, 135, 169–170. [Google Scholar] [CrossRef]
- Izzo, G.N.; Bashaw, M.J.; Campbell, J.B. Enrichment and Individual Differences Affect Welfare Indicators in Squirrel Monkeys (Saimiri sciureus). J. Comp. Psychol. 2011, 125, 347. [Google Scholar] [CrossRef]
- Alligood, C.A.; Dorey, N.R.; Mehrkam, L.R.; Leighty, K.A. Applying Behavior-Analytic Methodology to the Science and Practice of Environmental Enrichment in Zoos and Aquariums. Zoo Biol. 2017, 36, 175–185. [Google Scholar] [CrossRef]
- Learmonth, M.J.; Sherwen, S.; Hemsworth, P.H. Assessing Preferences of Two Zoo-Housed Aldabran Giant Tortoises (Aldabrachelys gigantea) for Three Stimuli Using a Novel Preference Test. Zoo Biol. 2021, 40, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, A.; Huber, L. Cold-Blooded Cognition: Reptilian Cognitive Abilities. In The Oxford Handbook of Comparative Evolutionary Psychology; Oxford University Press: Oxford, UK, 2012; pp. 129–141. [Google Scholar]
- Reading, R.P.; Miller, B.; Shepherdson, D. The Value of Enrichment to Reintroduction Success. Zoo Biol. 2013, 32, 332–341. [Google Scholar] [CrossRef]
- Westlund, K. Training Is Enrichment—And Beyond. Appl. Anim. Behav. Sci. 2014, 152, 1–6. [Google Scholar] [CrossRef]
- Degregorio, B.; Sperry, J.; Tuberville, T.; Weatherhead, P. Translocating Ratsnakes: Does Enrichment Offset Negative Effects of Time in Captivity? Wildl. Res. 2017, 44. [Google Scholar] [CrossRef]
- Roe, J.H.; Frank, M.R.; Kingsbury, B.A. Experimental Evaluation of Captive-Rearing Practices to Improve Success of Snake Reintroductions. Herpetol. Conserv. Biol. 2015, 10, 711–722. [Google Scholar]
- Roth, T.C.; Krochmal, A.R. The Role of Age-Specific Learning and Experience for Turtles Navigating a Changing Landscape. Curr. Biol. 2015, 25, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Meester, G.D.; Baeckens, S. Reinstating Reptiles: From Clueless Creatures to Esteemed Models of Cognitive Biology. Behaviour 2021, 158, 1057–1076. [Google Scholar] [CrossRef]
- Mehrkam, L.R.; Dorey, N.R. Preference Assessments in the Zoo: Keeper and Staff Predictions of Enrichment Preferences across Species. Zoo Biol. 2015, 34, 418–430. [Google Scholar] [CrossRef]
- Rosier, R.L.; Langkilde, T. Does Environmental Enrichment Really Matter? A Case Study Using the Eastern Fence Lizard, Sceloporus Undulatus. Appl. Anim. Behav. Sci. 2011, 131, 71–76. [Google Scholar] [CrossRef]
- Cronin, K.; Ross, S. Technical Contribution: A Cautionary Note on the Use of Behavioural Diversity (H-Index) in Animal Welfare Science. Anim. Welf. 2019, 28, 157–164. [Google Scholar] [CrossRef]
- Rosier, R.L.; Langkilde, T.L. In Response to the Letter to the Editor Regarding the Article: “Does Environmental Enrichment Really Matter? A Case Study Using the Eastern Fence Lizard, Sceloporus Undulatus”. Appl. Anim. Behav. Sci. 2011, 135, 171–172. [Google Scholar] [CrossRef]
- Tarou, L.R.; Bashaw, M.J. Maximizing the Effectiveness of Environmental Enrichment: Suggestions from the Experimental Analysis of Behavior. Appl. Anim. Behav. Sci. 2007, 102, 189–204. [Google Scholar] [CrossRef]
- Bennett, D. A Little Book of Monitor Lizards: A Guide to the Monitor Lizards of the World and Their Care in Captivity; Viper Press: Aberdeen, UK, 1995. [Google Scholar]
- Uetz, P.; Freed, P.; Hošek, J. The Reptile Database. Available online: https://reptile-database.reptarium.cz/ (accessed on 11 July 2021).
- Ziegler, T.; Rauhaus, A.; Gill, I. A Preliminary Review of Monitor Lizards in Zoological Gardens. Biawak 2016, 10, 26–35. [Google Scholar]
- Valdez, J.W. Using Google Trends to Determine Current, Past, and Future Trends in the Reptile Pet Trade. Animals 2021, 11, 676. [Google Scholar] [CrossRef]
- CITES. CITES Trade Database; UNEP World Conservation Monitoring Centre: Cambridge, UK, 2021. [Google Scholar]
- Pianka, E.; King, D. Varanoid Lizards of the World; Indiana University Press: Bloomington, IN, USA, 2004. [Google Scholar]
- Welton, L.J.; Siler, C.D.; Bennett, D.; Diesmos, A.; Duya, M.R.; Dugay, R.; Rico, E.L.B.; Van Weerd, M.; Brown, R.M. A Spectacular New Philippine Monitor Lizard Reveals a Hidden Biogeographic Boundary and a Novel Flagship Species for Conservation. Biol. Lett. 2010, 6, 654–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, S.J.; de Kort, S.R.; Bennett, D.; van Weerd, M. Diet and Habitat Requirements of the Philippine Endemic Frugivorous Monitor Lizard Varanus Bitatawa. Biawak 2018, 12, 12–22. [Google Scholar]
- Dryden, G.; Green, B.; King, D.; Losos, J. Water and Energy Turnover in a Small Monitor Lizard, Varanus-Acanthurus. Wildl. Res. 1990, 17, 641–646. [Google Scholar] [CrossRef]
- Frappell, P.B.; Schultz, T.J.; Christian, K.A. The Respiratory System in Varanid Lizards: Determinants of O2 Transfer. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 133, 239–258. [Google Scholar] [CrossRef]
- Cross, S.L.; Craig, M.D.; Tomlinson, S.; Bateman, P.W. I Don’t like Crickets, I Love Them: Invertebrates Are an Important Prey Source for Varanid Lizards. J. Zool. 2020, 310, 323–333. [Google Scholar] [CrossRef]
- Cieri, R.L.; Farmer, C.G. Computational Fluid Dynamics Reveals a Unique Net Unidirectional Pattern of Pulmonary Airflow in the Savannah Monitor Lizard (Varanus exanthematicus). Anat. Rec. 2020, 303, 1768–1791. [Google Scholar] [CrossRef] [PubMed]
- Schachner, E.R.; Cieri, R.L.; Butler, J.P.; Farmer, C.G. Unidirectional Pulmonary Airflow Patterns in the Savannah Monitor Lizard. Nature 2014, 506, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Clemente, C.J.; Withers, P.C.; Thompson, G.G. Metabolic Rate and Endurance Capacity in Australian Varanid Lizards (Squamata: Varanidae: Varanus). Biol. J. Linn. Soc. 2009, 97, 664–676. [Google Scholar] [CrossRef] [Green Version]
- Northcutt, R.G. Variation in Reptilian Brains and Cognition. Brain Behav. Evol. 2013, 82, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O.; Stacho, M.; Ströckens, F. The Brains of Reptiles and Birds. Evol. Neurosci. 2020, 159–212. [Google Scholar]
- Arena, P.; Warwick, C. Spatial and Thermal Considerations. In Health and Welfare of Captive Reptiles; Animal Welfare Series; Springer: Cham, Switzerland, 2022; Volume 22. [Google Scholar]
- Shettleworth, S.J. Animal Cognition and Animal Behaviour. Anim. Behav. 2001, 61, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Mendyk, R.W.; Horn, H.G. Skilled Forelimb Movements and Extractive Foraging in the Arboreal Monitor Lizard Varanus Beccarii (Doria, 1874). Herpetol. Rev. 2011, 42, 343–349. [Google Scholar]
- Cooper, T.; Liew, A.; Andrle, G.; Cafritz, E.; Dallas, H.; Niesen, T.; Slater, E.; Stockert, J.; Vold, T.; Young, M. Latency in Problem Solving as Evidence for Learning in Varanid and Helodermatid Lizards, with Comments on Foraging Techniques. Copeia 2019, 107, 78–84. [Google Scholar] [CrossRef]
- Cooper, T.L.; Zabinski, C.L.; Adams, E.J.; Berry, S.M.; Pardo-Sanchez, J.; Reinhardt, E.M.; Roberts, K.M.; Watzek, J.; Brosnan, S.F.; Hill, R.L. Long-Term Memory of a Complex Foraging Task in Monitor Lizards (Reptilia: Squamata: Varanidae). J. Herpetol. 2020, 54, 378–383. [Google Scholar] [CrossRef]
- Kane, D.; Davis, A.C.; Michaels, C.J. Play Behaviour by Captive Tree Monitors, Varanus Macraei and Varanus Prasinus. Herpetol. Bull. 2019, 149, 28–31. [Google Scholar] [CrossRef]
- King, D.; Green, B. Goannas: The Biology of Varanid Lizards; UNSW Press: Kensington, Australia, 1999. [Google Scholar]
- Korte, S.M.; Olivier, B.; Koolhaas, J.M. A New Animal Welfare Concept Based on Allostasis. Physiol. Behav. 2007, 92, 422–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meehan, C.L.; Mench, J.A. The Challenge of Challenge: Can Problem Solving Opportunities Enhance Animal Welfare? Appl. Anim. Behav. Sci. 2007, 102, 246–261. [Google Scholar] [CrossRef]
- Myers, D.G.; Diener, E. Who Is Happy? Psychol. Sci. 1995, 6, 10–19. [Google Scholar] [CrossRef]
- Nakamura, J.; Csikszentmihalyi, M. The Concept of Flow. In Flow and the Foundations of Positive Psychology; Springer: Berlin/Heidelberg, Germany, 2014; pp. 239–263. [Google Scholar]
- Loop, M.S. Auto-Shaping: A Simple Technique for Teaching a Lizard to Perform a Visual Discrimination Task. Copeia 1976, 1976, 574–576. [Google Scholar] [CrossRef]
- Firth, I.; Turner, M.; Robinson, M.; Meek, R. Response of Monitor Lizards (Varanus spp. ) to a Repeated Food Source: Evidence for Association Learning? Herpetol. Bull. 2003, 84, 1–4. [Google Scholar]
- Manrod, J.D.; Hartdegen, R.; Burghardt, G.M. Rapid Solving of a Problem Apparatus by Juvenile Black-Throated Monitor Lizards (Varanus albigularis albigularis). Anim. Cognit. 2008, 11, 267–273. [Google Scholar] [CrossRef]
- Gaalema, D.E. Visual Discrimination and Reversal Learning in Rough-Necked Monitor Lizards (Varanus rudicollis). J. Comp. Psychol. 2011, 125, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Ward-Fear, G.; Thomas, J.; Webb, J.K.; Pearson, D.J.; Shine, R. Eliciting Conditioned Taste Aversion in Lizards: Live Toxic Prey Are More Effective than Scent and Taste Cues Alone. Integr. Zool. 2017, 12, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Andrews, R.M.; Pezaro, N.; Doody, J.S.; Guarino, F.; Green, B. Oviposition to Hatching: Development of Varanus Rosenbergi. J. Herpetol. 2017, 51, 396–401. [Google Scholar] [CrossRef]
- Watterson, I.; Abbs, D.; Bhend, J.; Chiew, F.; Church, J.; Ekström, M.; Kirono, D.; Lenton, A.; Lucas, C.; McInnes, K. Climate Change in Australia Projections for Australia’s Natural Resource Management Regions; CSIRO: Canberra, Australia, 2015. [Google Scholar]
- Dayananda, B.; Webb, J.K. Incubation under Climate Warming Affects Learning Ability and Survival in Hatchling Lizards. Biol. Lett. 2017, 13, 20170002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abayarathna, T.; Webb, J.K. Effects of Incubation Temperatures on Learning Abilities of Hatchling Velvet Geckos. Anim. Cognit. 2020, 23, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Kis, A.; Huber, L.; Wilkinson, A. Social Learning by Imitation in a Reptile (Pogona vitticeps). Anim. Cognit. 2015, 18, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, D.W.; Byrne, R.W.; Whiting, M.J. Age-Dependent Social Learning in a Lizard. Biol. Lett. 2014, 10, 20140430. [Google Scholar] [CrossRef]
- Iwaniuk, A.N.; Nelson, J.E.; Pellis, S.M. Do Big-Brained Animals Play More? Comparative Analyses of Play and Relative Brain Size in Mammals. J. Comp. Psychol. 2001, 115, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Stanner, M. Mammal-like Feeding Behavior of Varanus Salvator and Its Conservational Implications. Biawak 2010, 4, 128–131. [Google Scholar]
- Burghardt, G.M. The Genesis of Animal Play: Testing the Limits; MIT Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Khandakar, N.; Jeny, K.N.; Islam, S.; Hakim, M.A.; Pony, I.A. Play Behavior by a Yellow Monitor, Varanus Flavescens (Hardwicke and Gray 1827). Reptiles Amphib. 2020, 27, 257–258. [Google Scholar] [CrossRef]
- Bartholomew, G.A.; Tucker, V.A. Size, Body Temperature, Thermal Conductance, Oxygen Consumption, and Heart Rate in Australian Varanid Lizards. Physiol. Zool. 1964, 37, 341–354. [Google Scholar] [CrossRef]
- Sweet, S.S. Spatial Ecology of Varanus Glauerti and V. Glebopalma in Northern Australia. Mertensiella 1999, 11, 317–366. [Google Scholar]
- Eidenmüller, B. Bisher Nicht Beschriebene Verhaltensweisen von Varanus (Varanus) Flavirufus Mertens 1958, Varanus (Odatria) Acanthurus Boulenger 1885 Und Varanus (Odatria) Storri Mertens 1966 Im Terrarium. Monitor 1993, 2, 11–21. [Google Scholar]
- D’Amore, D.C.; Clulow, S.; Doody, J.S.; Rhind, D.; McHenry, C.R. Claw Morphometrics in Monitor Lizards: Variable Substrate and Habitat Use Correlate to Shape Diversity within a Predator Guild. Ecol. Evol. 2018, 8, 6766–6778. [Google Scholar] [CrossRef]
- Krebs, U.; Wünstel, U. Observations and Experiments on “Spinning Behavior” in Varanus Albigularis. Biawak 2019, 13, 54–61. [Google Scholar]
- Mason, G.J.; Latham, N. Can’t Stop, Won’t Stop: Is Stereotypy a Reliable Animal Welfare Indicator? Anim. Welf. 2004, 13, S57–S69. [Google Scholar]
- Tilly, S.-L.C.; Dallaire, J.; Mason, G.J. Middle-Aged Mice with Enrichment-Resistant Stereotypic Behaviour Show Reduced Motivation for Enrichment. Anim. Behav. 2010, 80, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.G.; Lightman, S.L.; Gilmour, G.; Marston, H.; Robinson, E.S. Evidence for Deficits in Behavioural and Physiological Responses in Aged Mice Relevant to the Psychiatric Symptom of Apathy. Brain Neurosci. Adv. 2021, 5, 23982128211015110. [Google Scholar] [CrossRef] [PubMed]
- Meagher, R. Is Boredom an Animal Welfare Concern? Anim. Welf. 2018, 28, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Clark, F.E. Great Ape Cognition and Captive Care: Can Cognitive Challenges Enhance Well-Being? Appl. Anim. Behav. Sci. 2011, 135, 1–12. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The Use of the Elevated plus Maze as an Assay of Anxiety-Related Behavior in Rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howard, D.; Freeman, M.S. Overlooked and Under-Studied: A Review of Evidence-Based Enrichment in Varanidae. J. Zool. Bot. Gard. 2022, 3, 32-43. https://doi.org/10.3390/jzbg3010003
Howard D, Freeman MS. Overlooked and Under-Studied: A Review of Evidence-Based Enrichment in Varanidae. Journal of Zoological and Botanical Gardens. 2022; 3(1):32-43. https://doi.org/10.3390/jzbg3010003
Chicago/Turabian StyleHoward, Darcy, and Marianne Sarah Freeman. 2022. "Overlooked and Under-Studied: A Review of Evidence-Based Enrichment in Varanidae" Journal of Zoological and Botanical Gardens 3, no. 1: 32-43. https://doi.org/10.3390/jzbg3010003