
Assessment of Feeding Behavior of the Zoo-Housed Lesser Anteater (Tamandua tetradactyla) and Nutritional Values of Natural Prey

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Subjects, Enclosures, and Management
2.2. Procedures to Include Lesser Anteaters in Study 1
2.3. Study 1: Objective 1
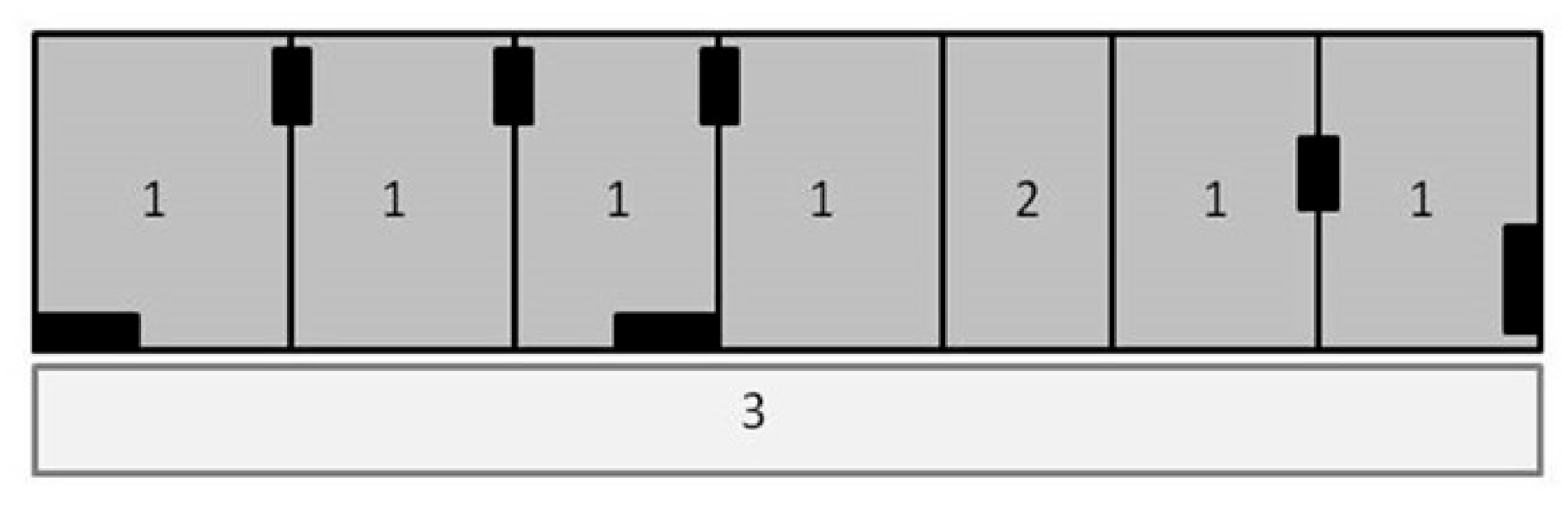
2.3.1. Evaluation Room
2.3.2. Behavioral Evaluations: Anteater Response to Food Stimuli
2.3.3. Methodology and Variables for Behavioral Evaluations
- -
- Ambulatory latency (s): time elapsed between placing the lesser anteater in the evaluation room with its four legs contacting the ground (see, for example, the cross in Figure 2a) and the complete head leaving the first square;
- -
- Visits to the feeders (counting): number of times the lesser anteater visited each feeder, considering a visit to be when the animal entered the square and contacted the feeder. On each session, we also registered which feeder lesser anteaters visited first;
- -
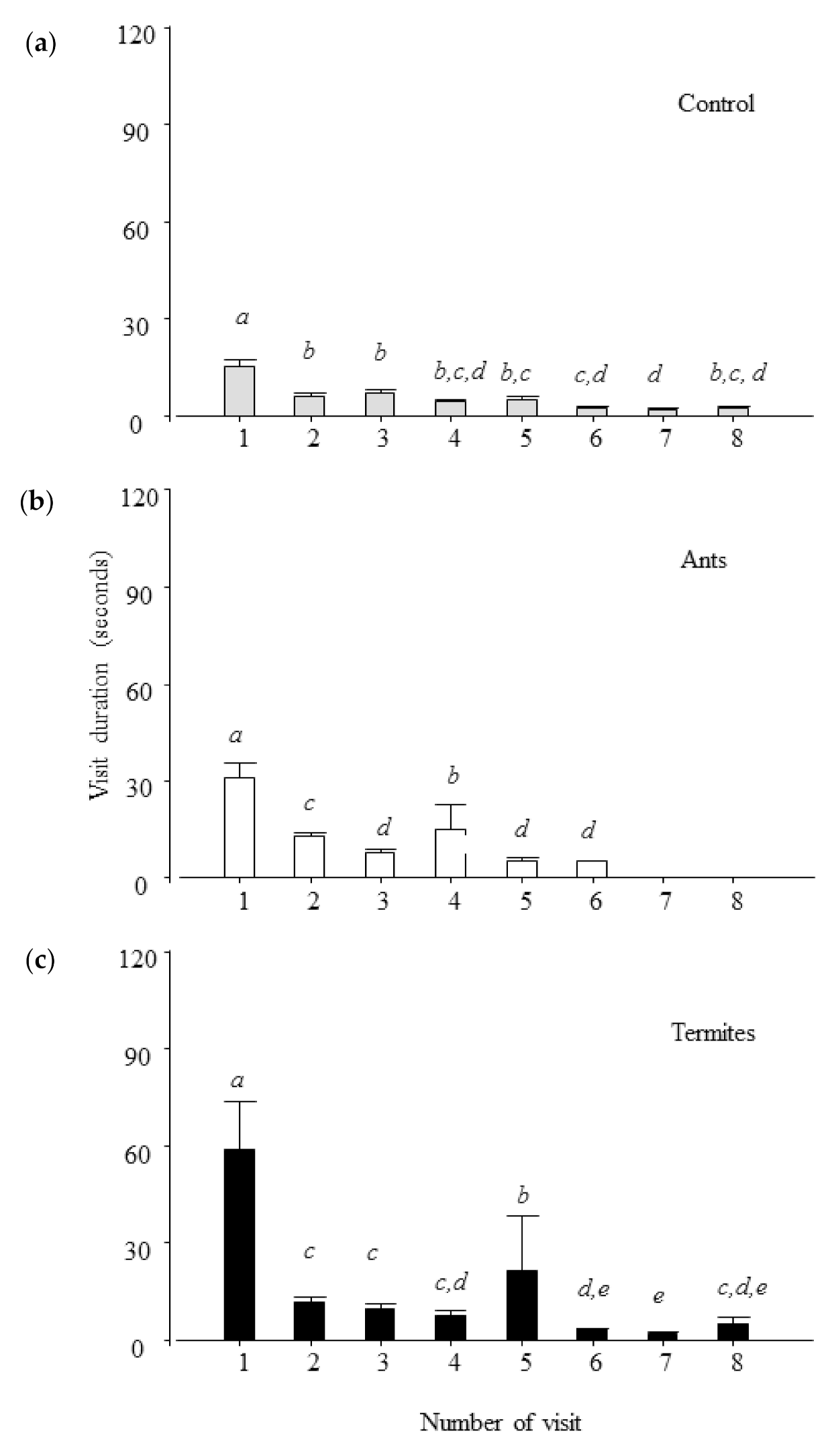
- Visit duration at feeders (s): time spent by each lesser anteater inside the square where the feeders were located, considering a visit to be each time the lesser anteater entered the square and contacted the feeder. We calculated the total visit duration per feeder. Considering the maximal number of visits in each feeder, we also calculated the visit duration per visit in order to characterize visits change over the 10 min of this trial;
- -
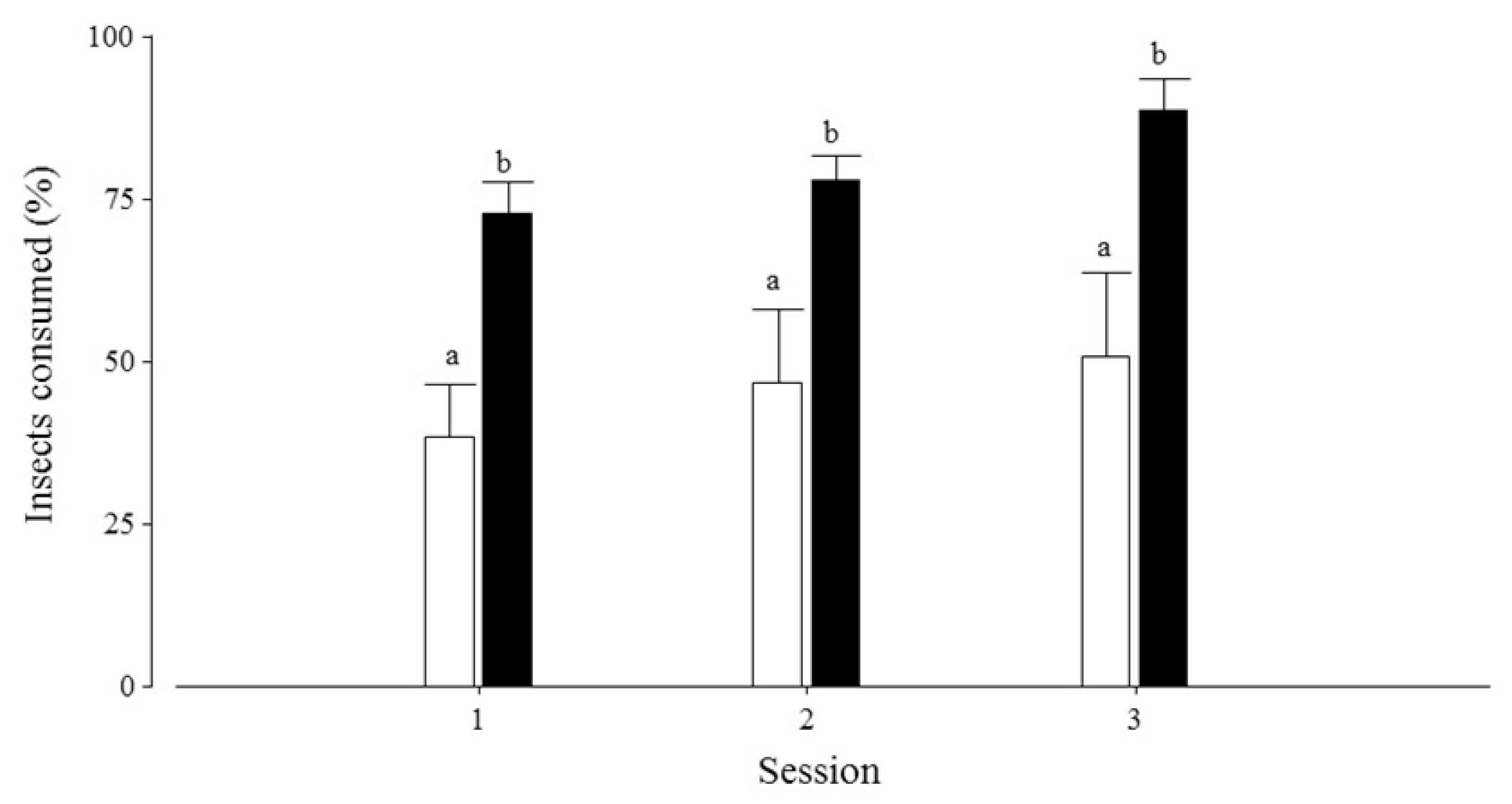
- Consumption of insects (g): quantity of insects consumed by each lesser anteater in each session. We calculated this variable based on the difference between the weight of the insect meal before and after the evaluation.
2.4. Study 2: Objective 2
2.4.1. Preparation of Insects and Nutritional Assessment
2.4.2. In Vitro Protein and Carbohydrate Digestibility
2.5. Statistical Analyses
3. Results
3.1. Study 1: Objective 1
3.2. Study 2: Objective 2
4. Discussion
5. Conclusions
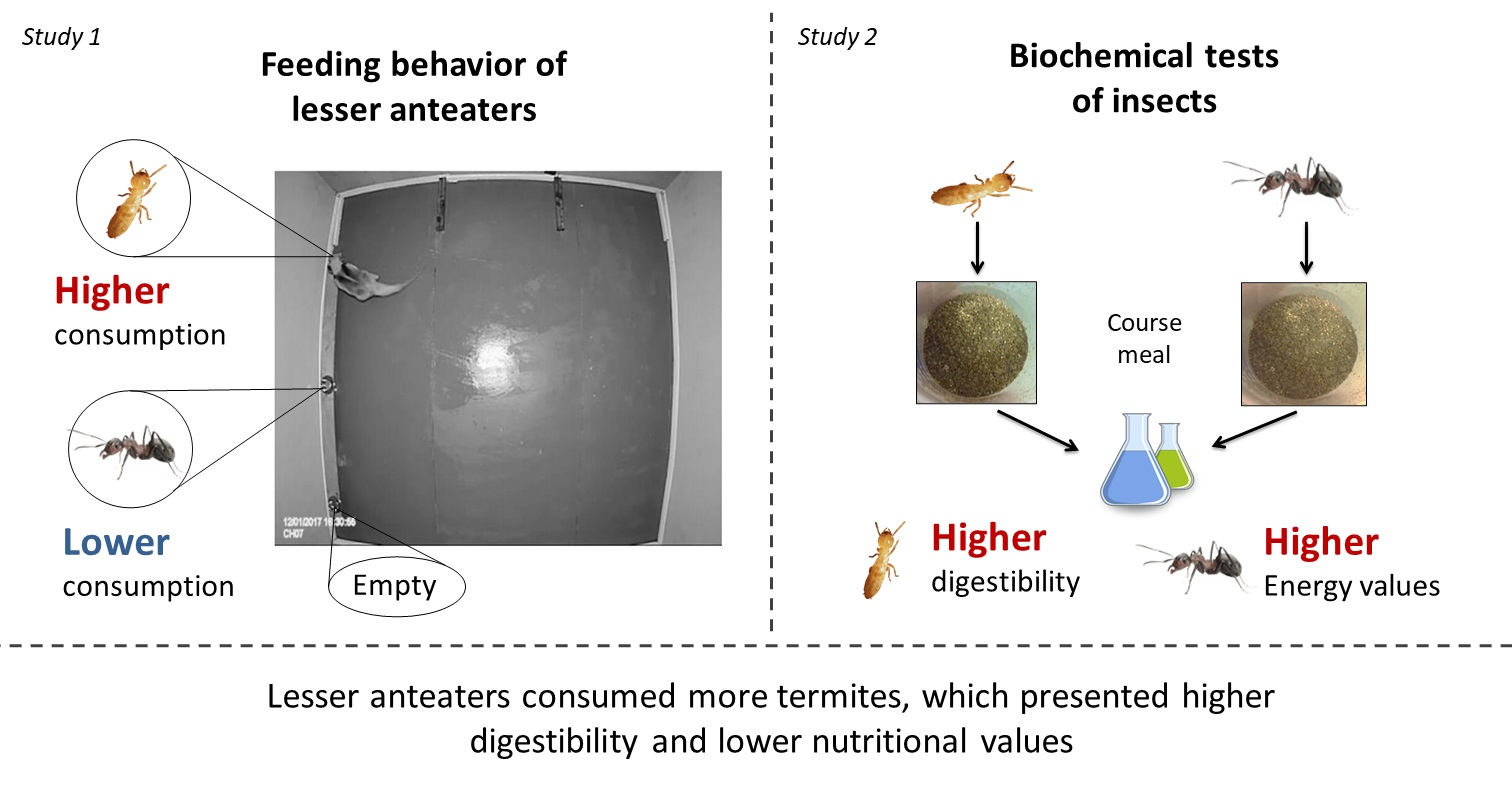
- Lesser anteaters spent more time at feeders with termites (C. fulviceps) than at feeders with ants (A. lundi) and consumed more termites than ants, showing a similar pattern of interaction during consumption;
- The ants and termites analyzed presented different values of humidity, protein, ash, lipids, and in vitro protein and carbohydrate digestibility. The calculation of caloric content of termites indicated lower energy values than ants but higher digestibility of proteins and carbohydrates.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hayssen, V. Tamandua tetradactyla (Pilosa: Myrmecophagidae). Mamm. Species 2011, 43, 64–74. [Google Scholar] [CrossRef]
- Montgomery, G.G. Movements, Foraging and Food Habits of the Four Extant Species of Neotropical Vermilinguas (Mammalia: Myrmecophagidae). In The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas; Montgomery, G.G., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1985; pp. 365–377, ISBN 10: 0874746493. [Google Scholar]
- Wetzel, R.M. The Identification and Distribution of Recent Xenarthra (= Edentata). In The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas; Mongomery, G.G., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1985; pp. 5–21, ISBN 10: 0874746493. [Google Scholar]
- Camilo-Alves, C.D.S.E.P.; Mourão, G.D.M. Responses of a Specialized Insectivorous Mammal (Myrmecophaga tridactyla) to Variation in Ambient Temperature. Biotropica 2006, 38, 52–56. [Google Scholar] [CrossRef]
- Lubin, Y.D.; Montgomery, G.G. Defenses of Nasutitermes termites (Isoptera, Termitidae) against Tamandua anteaters (Edentata, Myrmecophagidae). Biotropica 1981, 13, 66–76. [Google Scholar] [CrossRef]
- Sandoval-Gómez, V.E.; Ramérez-Chaves, H.E.; Marín, D. Registros de hormigas y termitas presentes en la dieta de osos hormigueros (Mammalia: Myrmecophagidae) en tres localidades de Colombia. Edentata 2012, 13, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Oyarzun, S.E.; Crawshaw, G.J.; Valdes, E.V. Nutrition of the tamandua: I. Nutrient composition of termites (Nasutitermes spp.) and stomach contents from wild tamanduas (Tamandua tetradactyla). Zoo Biol. 1996, 15, 509–524. [Google Scholar] [CrossRef]
- Torres, R.; Monguillot, J.; Bruno, G.; Michelutti, P.; Ponce, A. Ampliación del límite austral de la distribución del oso melero (Tamandua tetradactyla) en la Argentina. Not. Faun. 2009, 39, 1–5. [Google Scholar]
- Tamburini, D. Orden Cingulata. In Mamíferos de Córdoba y su Estado de Conservación; Torres, R., Tamburini, D., Eds.; Editorial de la Universidad Nacional de Córdoba: Cordoba, Argentina, 2018; pp. 87–111. ISBN 978-987-707-084-2. [Google Scholar]
- Superina, M.; Abba, A.M.; Vizcaíno, S.F. Orden Pilosa. In Libro Rojo de los Mamíferos de Argentina; Ojeda, R.A., Chillo, V., Díaz Isenrath, G., Eds.; Sociedad Argentina para el Estudio de los Mamíferos: Buenos Aires, Argentina, 2012; pp. 59–60. ISBN 987-987-98497-2-9. [Google Scholar]
- Miranda, F.; Fallabrino, A.; Arteaga, M.; Tirira, D.G.; Meritt, D.A.; Superina, M. Tamandua tetradactyla. The IUCN Red List of Threatened Species 2014: E.T21350A47442916. Available online: https://www.iucnredlist.org/ (accessed on 20 October 2020).
- Moore, B.D.; Marsh, K.J.; Wallis, I.R.; Foley, W.J. Taught by animals: How understanding diet selection leads to better zoo diets. Int. Zoo. Yearb. 2005, 39, 43–61. [Google Scholar] [CrossRef]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Aguilar, R.F.; Superina, M. Xenarthra. In Fowler’s Zoo and Wild Animal Medicine; Miller, R.E., Fowler, M.E., Eds.; Elsevier Saunders: St. Louis, MO, USA, 2015; pp. 355–369. ISBN 978-1-4557-7397-8. [Google Scholar] [CrossRef]
- Ward, A.M.; Crisse, S.D.; Cassaro, K.; Frank, E. Formulating Diets for Tamanduas (T. tetradactyla) in Brazilian Zoos. In Proceedings of the First Annual Conference of the Nutrition Advisory Group of the American Zoo and Aquarium Association, Toronto, ON, Canada, 1–2 May 1995; Dierenfeld, E., Atkinson, J., Valdes, E.V., Eds.; Metro Toronto Zoo and the University of Guelph: Toronto, ON, Canada, 1995. [Google Scholar]
- Morford, S.; Meyers, M.A. Giant anteater (Myrmecophaga tridactyla) diet survey. Edentata 2003, 5, 20–24. [Google Scholar]
- Ruiz, D.; Tello, M. Nutrición y Alimentación en la Rehabilitación de Vermilinguas y Estrategias Nutricionales Pre-Liberación. In Manual de Rehabilitación de Hormigueros de Colombia; Rojano, C., Miranda, L., Ávila, R., Eds.; Fundación Cunaguaro, Geopark Colombia S.A.S, Corporinoquía: El Yopal, Columbia, 2014; pp. 69–83. [Google Scholar]
- Eguizábal, G.V.; Palme, R.; Villarreal, D.; Dal Borgo, C.; Di Rienzo, J.A.; Busso, J.M. Assessment of adrenocortical activity and behavior of the collared anteater (Tamandua tetradactyla) in response to food-based environmental enrichment. Zoo Biol. 2013, 32, 632–640. [Google Scholar] [CrossRef]
- Neto, G.D.S.F.; Barros, A.C.; Sobroza, T.V.; Neves, P.U.C.; Phillips, M.J.; Guimarães, E.F. Period of the day and food-based enrichment affect behaviour activity of Tamandua tetradactyla in captivity? Pesqui. Ensino Cienc. Exatas Nat. 2020, 4, e1498. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard Press: Cambridge, MA, USA, 1990; ISBN 978-3-540-52092-4. [Google Scholar]
- Torales, G.J.; Laffont, E.R.; Godoy, M.C.; Coronel, J.M.; Arbino, M.O. Update on taxonomy and distribution of Isoptera from Argentina. Sociobiology 2005, 45, 853–886. [Google Scholar]
- Borror, D.J.; Triplehorn, C.A.; Johnson, N. An Introduction to the Study of Insects, 6th ed.; Holt Rinehart and Winston Saunders College Publishing: Philadelphia, PA, USA, 1989; ISBN 0-03-025397-7. [Google Scholar]
- Ali, M.F.; Morgan, E.D. Chemical communication in insect communities: A guide to insect pheromones with special emphasis on social insects. Biol. Rev. 1990, 65, 227–247. [Google Scholar] [CrossRef]
- Redford, K.H. Mammalian Myrmecophagy: Feeding, Foraging and Food Preference. Ph.D. Thesis, Harvard University, Cambridge, MA, USA, 1983. [Google Scholar]
- Redford, K.H.; Dorea, J.G. The nutritional value of invertebrates with emphasis on ants and termites as food for mammals. J. Zool. 1984, 203, 385–395. [Google Scholar] [CrossRef]
- Stephens, D.W.; Krebs, J.R. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar] [CrossRef]
- Watters, J.V.; Krebs, B.L.; Eschmann, C.L. Assessing Animal Welfare with Behavior: Onward with Caution. J. Zool. Bot. Gard. 2021, 2, 75–87. [Google Scholar] [CrossRef]
- Eguizábal, G.V.; Palme, R.; Superina, M.; Asencio, C.J.; García Capocasa, M.C.; Busso, J.M. Characterization and correlations of behavioral and adrenocortical activities of zoo-housed lesser anteaters (Tamandua tetradactyla). Zoo Biol. 2019, 38, 334–342. [Google Scholar] [CrossRef]
- Chiapero, F.; Ferrari, H.R.; Prieto, M.V.; Garcia Capocasa, M.C.; Busso, J.M. Multivariate analyses of the activity pattern and behavior of the lesser anteater on open and closed days at Córdoba Zoo, Argentina. J. Appl. Anim. Welf. Sci. 2021, 24, 83–97. [Google Scholar] [CrossRef]
- Mellor, D.J.; Beausoleil, N.J. Extending the ‘Five Domains’ model for animal welfare assessment to incorporate positive welfare states. Anim. Welf. 2015, 24, 241–253. [Google Scholar] [CrossRef]
- Superina, M.; Miranda, F.; Plese, T. Maintenance of Xenarthra in Captivity. In The Biology of the Xenarthra; Vizcaıno, S.F., Loughry, W.J., Eds.; University Press of Florida: Gainesville, FL, USA, 2008; pp. 232–243. ISBN 9780813031651. [Google Scholar]
- Dierenfeld, E.S.; Graffam, W.S. Manual de Nutrición y Dietas para Animales Silvestres en Cautiverio (Ejemplos para América Latina); ZCOG Zoo Conservation Outreach Group: New Orleans, LA, USA, 1996; pp. 42–43. [Google Scholar]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry, 5th ed.; W.H. Freeman and Company: New York, NY, USA, 2002; ISBN-10: 0-7167-3051-0. [Google Scholar]
- Boisen, S.; Fernández, J.A. Prediction of the total tract digestibility of energy in feedstuffs and pig diets by in vitro analyses. Anim. Feed Sci. Technol. 1997, 68, 277–286. [Google Scholar] [CrossRef]
- Gull, J.M.; Stahl, M.; Osmann, C.; Ortmann, S.; Kreuzer, M.; Hatt, J.M.; Clauss, M. Digestive physiology of captive giant anteaters (Myrmecophaga tridactyla): Determinants of faecal dry matter content. J. Anim. Physiol. Anim. Nutr. 2015, 99, 565–576. [Google Scholar] [CrossRef]
- Lin, M.F.; Chang, C.Y.; Yang, C.W.; Dierenfeld, E.S. Aspects of digestive anatomy, feed intake and digestion in the Chinese pangolin (Manis pentadactyla) at Taipei Zoo. Zoo Biol. 2015, 34, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Dierick, N.; Vervaeke, I.; Decuypere, J.; Henderickx, H. Protein Digestion in Pig Measured In Vivo and In Vitro. In Proceedings of the Third International Seminar on the Digestive Physiology in the Pig, Copenhagen, Denmark, 16–18 May 1985; Just, A., Jorgensen, H., Fernández, J.A., Eds.; National Institute of Animal Science: Copenhagen, Denmark, 1985; pp. 329–332. Available online: https://agris.fao.org/agris-search/search.do?recordID=DK8620175 (accessed on 24 December 2021).
- Loewus, F.A. Improvement in anthrone method for determination of carbohydrates. Anal. Chem. 1952, 24, 219. [Google Scholar] [CrossRef]
- Mangeaud, A.; Videla, M. Looking for the lost independence: Using Mixed Generalized Linear Models in choice tests. Ecol. Austral. 2005, 15, 199–206. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. Infostat Versión 2019; Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2019. [Google Scholar]
- Bateson, M. Mechanisms of decision-making and the interpretation of choice tests. Anim. Welf. 2004, 13, S115–S120. [Google Scholar]
- Warburton, H.; Mason, G. Is out of sight out of mind? The effects of resource cues on motivation in mink, Mustela vison. Anim. Behav. 2003, 65, 755–762. [Google Scholar] [CrossRef] [Green Version]
- Wells, D.L. Sensory stimulation as environmental enrichment for captive animals: A review. Appl. Anim. Behav. Sci. 2009, 118, 1–11. [Google Scholar] [CrossRef]
- Lubin, Y.D.; Montgomery, G.G.; Young, O.P. Food resources of anteaters (Edentata: Myrmecophagidae) I. A year’s census of arboreal nests of ants and termites on Barro Colorado Island, Panama Canal Zone. Biotropica 1977, 9, 26–34. [Google Scholar] [CrossRef]
- Redford, K.H. Feeding and food preference in captive and wild giant anteater (Myrmecophaga tridactyla). J. Zool. 1985, 205, 559–572. [Google Scholar] [CrossRef]
- Bell, G.P. Birds and mammals on an insect diet: A primer on diet composition analysis in relation to ecological energetics. Stud. Avian Biol. 1990, 13, 416–422. [Google Scholar]
- Allen, M.E. Nutritional Aspects of Insectivory. Ph.D. Dissertation, Michigan State University, East Lansing, MI, USA, 1989. Available online: https://d.lib.msu.edu/etd/16494 (accessed on 20 October 2020).
- Emerling, C.A.; Delsuc, F.; Nachman, M.W. Chitinase genes (CHIAs) provide genomic footprints of a post-Cretaceous dietary radiation in placental mammals. Sci. Adv. 2018, 4, eaar6478. [Google Scholar] [CrossRef] [Green Version]
- Diniz, L.S.M.; Costa, E.O.; Oliveira, P.M.A. Clinical disorders observed in anteaters (Myrmecophagidae, Edentata) in captivity. Vet. Res. Commun. 1995, 19, 409–415. [Google Scholar] [CrossRef]
- Arenales, A.; Gardiner, C.H.; Miranda, F.R.; Dutra, K.S.; Oliveira, A.R.; Mol, J.P.; Texeira Da Costa, E.L.M.; Tinoco, H.P.; Coelho, C.M.; Silva, R.O.S.; et al. Pathology of Free-Ranging and Captive Brazilian Anteaters. J. Comp. Pathol. 2020, 180, 55–68. [Google Scholar] [CrossRef]
- Kaspari, M.; Alonso, L.; O’Donnell, S. Three energy variables predict ant abundance at a geographical scale. Proc. R. Soc. Lond. B Biol. Sci. 2000, 267, 485–489. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable (%) | Insects | |
|---|---|---|
| Ants | Termites | |
| Humidity | 47.37 ± 0.52 | 42.19 ± 0.73 |
| Lipids | 10.88 ± 0.24 | 4.08 ± 0.43 |
| Proteins | 29.06 ± 0.74 | 12.96 ± 0.67 |
| Ash | 7.61 ± 0.07 | 12.45 ± 0.77 |
| Carbohydrates | 14.71 | 32.20 |
| In vitro protein digestibility | 41.66 ± 0.43 | 97.84 ± 1.63 |
| In vitro carbohydrate digestibility | 50.65 ± 0.87 | 75.34 ± 0.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zárate, V.; Mufari, J.R.; Abalos Luna, L.G.; Villarreal, D.P.; Busso, J.M. Assessment of Feeding Behavior of the Zoo-Housed Lesser Anteater (Tamandua tetradactyla) and Nutritional Values of Natural Prey. J. Zool. Bot. Gard. 2022, 3, 19-31. https://doi.org/10.3390/jzbg3010002
Zárate V, Mufari JR, Abalos Luna LG, Villarreal DP, Busso JM. Assessment of Feeding Behavior of the Zoo-Housed Lesser Anteater (Tamandua tetradactyla) and Nutritional Values of Natural Prey. Journal of Zoological and Botanical Gardens. 2022; 3(1):19-31. https://doi.org/10.3390/jzbg3010002
Chicago/Turabian StyleZárate, Valentín, Jesica R. Mufari, Lucía G. Abalos Luna, Daniel P. Villarreal, and Juan M. Busso. 2022. "Assessment of Feeding Behavior of the Zoo-Housed Lesser Anteater (Tamandua tetradactyla) and Nutritional Values of Natural Prey" Journal of Zoological and Botanical Gardens 3, no. 1: 19-31. https://doi.org/10.3390/jzbg3010002