
Investigating the Effect of Disturbance on Prey Consumption in Captive Congo Caecilians Herpele squalostoma


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Models, Experimental Design, and Data-Collection
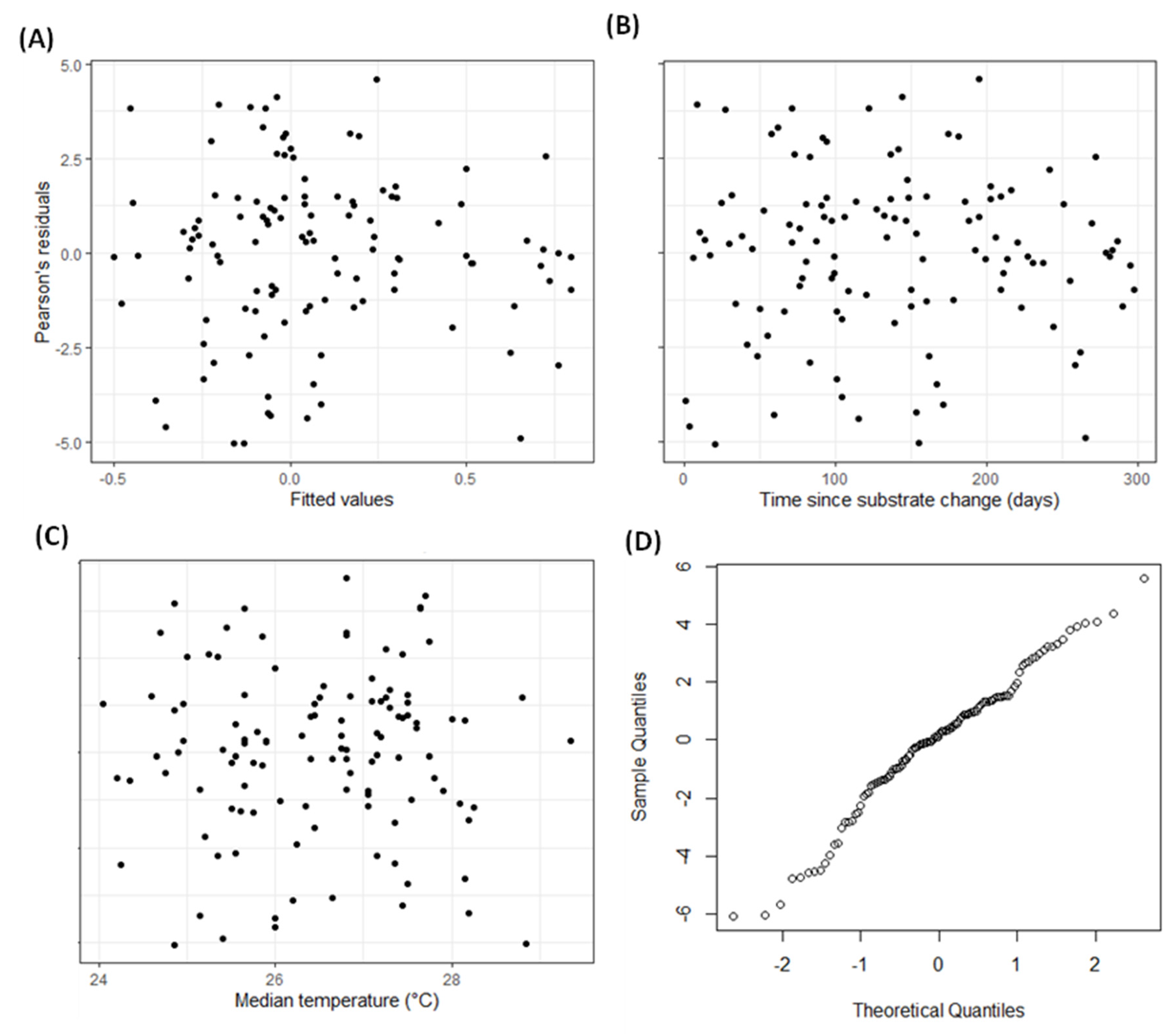
2.2. Statistical Analysis
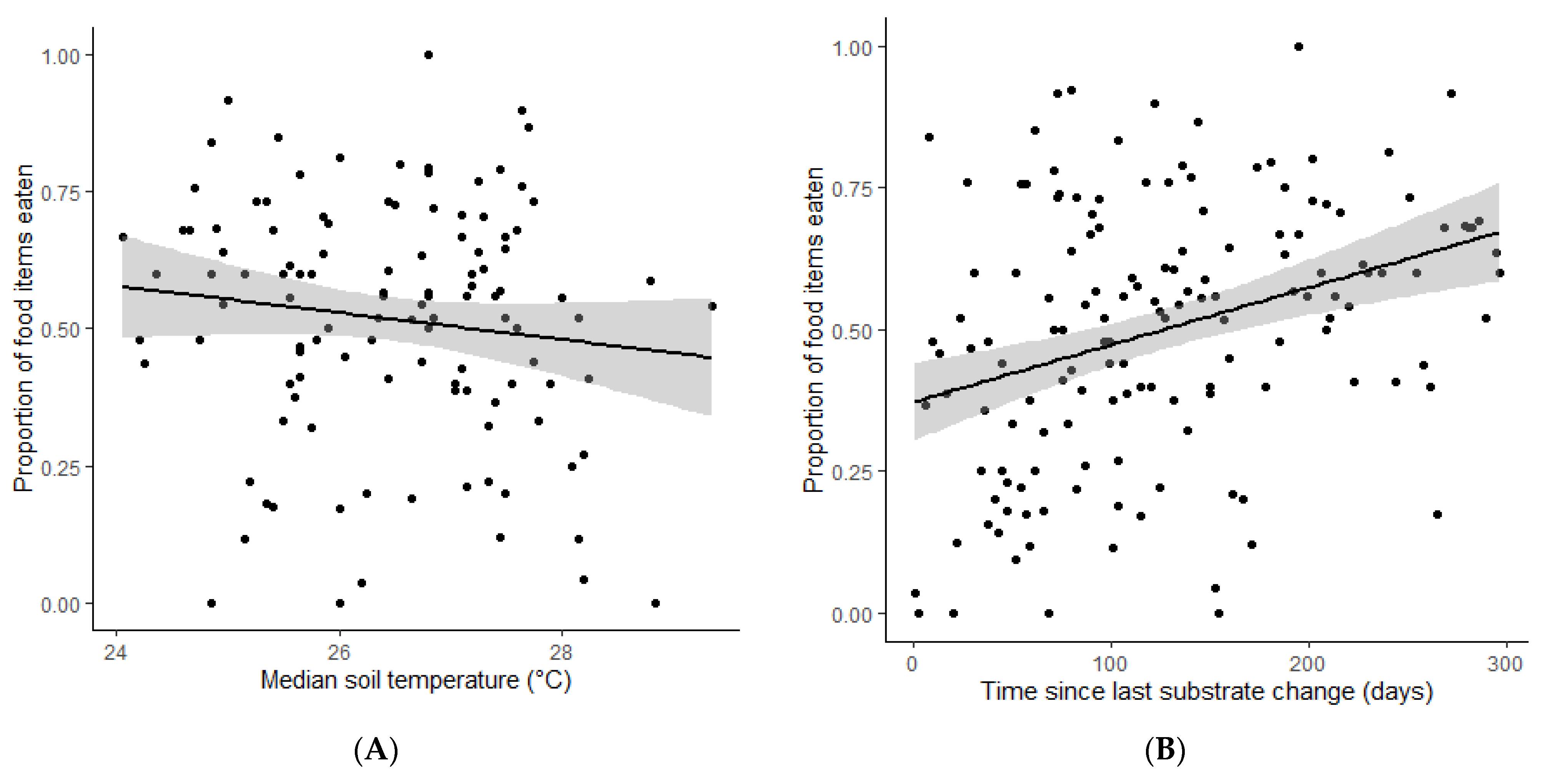
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Whitham, J.C.; Wielebnowski, N. New directions for zoo animal welfare science. Appl. Anim. Behav. Sci. 2013, 147, 247–260. [Google Scholar] [CrossRef]
- Department of the Environment, Transport and the Regions. Secretary of State’s Standards of Modern Zoo Practice; Department of the Environment, Transport and the Regions: London, UK, 2012. [Google Scholar]
- Van Waeyenberge, J.; Aerts, J.; Hellebuyck, T.; Pasmans, F.; Martel, A. Stress in wild and captive snakes: Quantification, effects and the importance of management. Vlaams Diergeneeskd. Tijdschr. 2018, 87, 59–65. [Google Scholar] [CrossRef]
- Narayan, E.J.; Forsburg, Z.R.; Davis, D.R.; Gabor, C.R. Non-invasive methods for measuring and monitoring stress physiology in imperiled amphibians. Front. Ecol. Evol. 2019, 7, 431. [Google Scholar] [CrossRef] [Green Version]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Warwick, C.; Arena, P.; Lindley, S.; Jessop, M.; Steedman, C. Assessing reptile welfare using behavioural criteria. Practice 2013, 35, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Hossie, T.J.; Ferland-Raymond, B.; Burness, G.; Murray, D.L. Morphological and behavioural responses of frog tadpoles to perceived predation risk: A possible role for corticosterone mediation? Ecoscience 2010, 17, 100–108. [Google Scholar] [CrossRef]
- Woody, S.M.; Santymire, R.M.; Cronin, K.A. Posture as a Non-Invasive Indicator of Arousal in American Toads (Anaxyrus americanus). J. Zool. Bot. Gard. 2021, 2, 1–9. [Google Scholar] [CrossRef]
- Benn, A.L.; McLelland, D.J.; Whittaker, A.L. A review of welfare assessment methods in reptiles, and preliminary application of the welfare quality® protocol to the pygmy blue-tongue skink, Tiliqua adelaidensis, using animal-based measures. Animals 2019, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Frost, D.R. Amphibian Species of the World: An Online Reference; Version 6.1; American Museum of Natural History: New York, NY, USA, 2021; Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 17 August 2021). [CrossRef]
- Species360. 2021. Available online: www.Species360.org (accessed on 15 August 2021).
- Gower, D.J.; Wilkinson, M. The conservation biology of caecilians. Conserv. Biol. 2005, 19, 45–55. [Google Scholar] [CrossRef]
- Tapley, B.; Michaels, C.J.; Gower, D.J.; Wilkinson, M. The use of visible implant elastomer to permanently identify caecilians (Amphibia: Gymnophiona). Herpetol. Bull. 2019, 150. [Google Scholar] [CrossRef]
- Lavelle, P.; Bignell, D.; Lepage, M.; Wolters, V.; Roger, P.A.; Ineson, P.O.W.H.; Heal, O.W.; Dhillion, S. Soil function in a changing world: The role of invertebrate ecosystem engineers. Eur. J. Soil Biol. 1997, 33, 159–193. [Google Scholar]
- Measey, G.J.; Gower, D.J.; Oommen, O.V.; Wilkinson, M. A mark–recapture study of the caecilian amphibian Gegeneophis ramaswamii (Amphibia: Gymnophiona: Caeciliidae) in southern India. J. Zool. 2003, 261, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Loader, S.P.; Gower, D.J. Trophic ecology of East African caecilians (Amphibia: Gymnophiona), and their impact on forest soil invertebrates. J. Zool. 2006, 269, 117–126. [Google Scholar] [CrossRef]
- Jared, C.; Antoniazzi, M.M.; Wilkinson, M.; Delabie, J.H. Conservation of the caecilian Siphonops annulatus (Amphibia, Gymnophiona) in Brazilian cacao plantations: A successful relationship between a fossorial animal and an agrosystem. Agrotrópica 2015, 27, 233–238. [Google Scholar] [CrossRef]
- Bonnet, X.; Shine, R.; Lourdais, O. Taxonomic chauvinism. Trends Ecol. Evol. 2002, 17, 1–3. [Google Scholar] [CrossRef]
- O’Reilly, J.C. Feeding in caecilians. In Feeding: Form, Function and Evolution in Tetrapod Vertebrates; Elsevier: Amsterdam, The Netherlands, 2000; pp. 149–166. [Google Scholar]
- Wilkinson, M.; Müller, H.; Gower, D.J. On Herpele multiplicata (Amphibia: Gymnophiona: Caeciliidae). Afr. J. Hepatol. 2003, 52, 119–122. [Google Scholar]
- Maddock, S.T.; Lewis, C.J.; Wilkinson, M.; Day, J.J.; Morel, C.; Kouete, M.; Gower, D.T. Non-lethal DNA sampling for caecilian amphibians. Herpetol. J. 2014, 24, 255–260. [Google Scholar]
- Tapley, B.; Bryant, Z.; Grant, S.; Kother, G.; Feltrer, Y.; Masters, N.; Strike, T.; Gill, I.; Wilkinson, M.; Gower, D.J. Towards evidence-based husbandry for caecilian amphibians: Substrate preference in Geotrypetes seraphini (Amphibia: Gymnophiona: Dermophiidae). Herpetol. Bull. 2014, 129, 15–18. [Google Scholar]
- Rendle, M.E.; Tapley, B.; Perkins, M.; Bittencourt-Silva, G.; Gower, D.J.; Wilkinson, M. Itraconazole treatment of Batrachochytrium dendrobatidis (Bd) infection in captive caecilians (Amphibia: Gymnophiona) and the first case of Bd in a wild neotropical caecilian. J. Zoo Aquar. Res. 2015, 3, 137–140. [Google Scholar]
- Flach, E.J.; Feltrer, Y.; Gower, D.J.; Jayson, S.; Michaels, C.J.; Pocknell, A.; Rivers, S.; Perkins, M.; Rendle, M.E.; Stidworthy, M.F.; et al. Postmortem findings in eight species of captive caecilian (Amphibia: Gymnophiona) over a ten-year period. J. Zoo Wildl. Med. 2020, 50, 79–890. [Google Scholar] [CrossRef]
- Whatley, C.; Tapley, B.; Michaels, C.J. Substrate preference in the fossorial caecilian Microcaecila unicolor (Amphibia: Gymnophiona, Siphonopidae). Herpetol. Bull. 2020, 152, 18–20. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Herpele squalostoma. The IUCN Red List of Threatened Species 2018: E.T59565A16958011. Available online: https://doi.org/10.2305/IUCN.UK.2018-1.RLTS.T59565A16958011.en (accessed on 1 August 2021).
- Wilkinson, M.; Sherratt, E.; Starace, F.; Gower, D.J. A new species of skin-feeding caecilian and the first report of reproductive mode in Microcaecilia (Amphibia: Gymnophiona: Siphonopidae). PLoS ONE 2013, 8, e57756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouete, M.T.; Ndeme, E.S.; Gower, D.J. Further observations of reproduction and confirmation of oviparity in Herpele squalostoma (Stutchbury, 1836) (Amphibia: Gymnophiona: Herpelidae). Herpetol. Notes 2013, 6, 583–586. [Google Scholar]
- Michaels, C.J.; Gini, B.F.; Preziosi, R.F. The importance of natural history and species-specific approaches in amphibian ex-situ conservation. Herpetol. J. 2014, 24, 135–145. [Google Scholar]
- Channing, A.; Rödel, M.O. Field Guide to the Frogs & Other Amphibians of Africa; Penguin Random House: Cape Town, South Africa, 2019. [Google Scholar]
- Kouete, M.T.; Wilkinson, M.; Gower, D.J. First reproductive observations for Herpele Peters, 1880 (Amphibia: Gymnophiona: Herpelidae): Evidence of extended parental care and maternal dermatophagy in H. squalostoma (Stutchbury, 1836). Int. Sch. Res. Notices 2012, 2012, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kupfer, A.; Maxwell, E.; Reinhard, S.; Kuehnel, S. The evolution of parental investment in caecilian amphibians: A comparative approach. Biol. J. Linn. Soc. 2016, 119, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Measey, G.J.; Gower, D.J.; Oommen, O.V.; Wilkinson, M. A subterranean generalist predator: Diet of the soil-dwelling caecilian Gegeneophis ramaswamii (Amphibia; Gymnophiona; Caeciliidae) in southern India. C. R. Biol. 2004, 327, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Kouete, M.T.; Blackburn, D.C. Dietary partitioning in two co-occurring caecilian species (Geotrypetes seraphini and Herpele squalostoma) in Central Africa. Integr. Org. Biol. 2020, 2, obz035. [Google Scholar] [CrossRef] [Green Version]
- ImageJ. Image Processing and Analysis in Java. Available online: https://imagej.nih.gov/ij/ (accessed on 16 August 2021).
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Millar, R.B.; Anderson, M.J. Remedies for pseudoreplication. Fish. Res. 2004, 70, 397–407. [Google Scholar] [CrossRef]
- Johnson, P.C. Extension of Nakagawa & Schielzeth’s R2GLMM to random slopes models. Methods Ecol. Evol. 2014, 5, 944–946. [Google Scholar] [PubMed] [Green Version]
- Raudenbush, S.W.; Yang, M.L.; Yosef, M. Maximum likelihood for generalized linear models with nested random effects via high-order, multivariate Laplace approximation. J. Comput. Graph. Stat. 2000, 9, 141–157. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. Mu-MIn: Multi-model inference. R Package Version 1.43.17. 2020. Available online: http://R-Forge.R-project.org/projects/mumin/ (accessed on 22 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org/ (accessed on 22 September 2021).
- Rouse, J.D.; Bishop, C.A.; Struger, J. Nitrogen pollution: An assessment of its threat to amphibian survival. Environ. Health Perspect. 1999, 107, 799–803. [Google Scholar] [CrossRef]
- Lentini, A.M. 34. Husbandry and care of amphibians. In Zookeeping; University of Chicago Press: Chicago, IL, USA, 2013; pp. 335–346. [Google Scholar]
- Densmore, C.L.; Green, D.E. Diseases of amphibians. ILAR J. 2007, 48, 235–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odum, R.A.; Zippel, K.C. Amphibian water quality: Approaches to an essential environmental parameter. Int. Zoo Yearb. 2008, 42, 40–52. [Google Scholar] [CrossRef]
- Voorhies, M.R. Vertebrate burrows. In The Study of Trace Fossils; Springer: Berlin/Heidelberg, Germany, 1975; pp. 325–350. [Google Scholar]
- Gower, D.J.; Loader, S.P.; Moncrieff, C.B.; Wilkinson, M. Niche separation and comparative abundance of Boulengerula boulengeri and Scolecomorphus vittatus (Amphibia: Gymnophiona) in an East Usambara forest, Tanzania. Afr. J. Herpetol. 2004, 53, 183–190. [Google Scholar] [CrossRef]
- Measey, G.J. Are caecilians rare? An east African perspective. J. East Afr. Nat. Hist. 2004, 93, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Kupfer, A.; Wilkinson, M.; Gower, D.J.; Müller, H.; Jehle, R. Care and parentage in a skin-feeding caecilian amphibian. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2008, 309, 460–467. [Google Scholar] [CrossRef]
- Kupfer, A.; Nabhitabhata, J.; Himstedt, W. Life history of amphibians in the seasonal tropics: Habitat, community and population ecology of a caecilian (genus Ichthyophis). J. Zool. 2005, 266, 237–247. [Google Scholar] [CrossRef]
- Michaels, C.J.; Preziosi, R.F. Clinical and naturalistic substrates differ in bacterial communities and in their effects on skin microbiota in captive fire salamanders (Salamandra salamandra). Herpetol. Bull. 2020, 151, 10–16. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Final Model | Estimate | Std. Error | z-Value | OR | 2.5% CI | 97.5% CI |
|---|---|---|---|---|---|---|
| Parameter estimates | ||||||
| Intercept | 2.580 | 0.848 | 3.041 | 2.506 | 69.789 | |
| Days since substrate change | 0.004 | 0.0005 | 7.678 | 1.004 | 1.003 | 1.005 |
| Average substrate temperature | −0.11 | 0.032 | −3.524 | 0.896 | 0.838 | 0.951 |
| Model fit | ||||||
| R2 marginal | 0.453 | |||||
| R2 conditional | 0.453 | |||||
| Dispersion estimate (Chi2) | 5.49 |
| Tank Substrate | Fresh Substrate | |||||
|---|---|---|---|---|---|---|
| Ammonia mg/L | Nitrite mg/L | Nitrate mg/L | Ammonia mg/L | Nitrite mg/L | Nitrate mg/L | |
| Mean | 0.03 | 0.03 | 0 | 0.04 | 0.06 | 2.00 |
| Median | 0.02 | 0.03 | 0 | 0 | 0.05 | 2.00 |
| Range | 0–0.07 | 0.02–0.04 | 0 | 0–0.12 | 0.05–0.07 | 2.00 |
| N | 3 | 3 | 3 | 3 | 3 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carter, K.C.; Fieschi-Méric, L.; Servini, F.; Wilkinson, M.; Gower, D.J.; Tapley, B.; Michaels, C.J. Investigating the Effect of Disturbance on Prey Consumption in Captive Congo Caecilians Herpele squalostoma. J. Zool. Bot. Gard. 2021, 2, 705-715. https://doi.org/10.3390/jzbg2040050
Carter KC, Fieschi-Méric L, Servini F, Wilkinson M, Gower DJ, Tapley B, Michaels CJ. Investigating the Effect of Disturbance on Prey Consumption in Captive Congo Caecilians Herpele squalostoma. Journal of Zoological and Botanical Gardens. 2021; 2(4):705-715. https://doi.org/10.3390/jzbg2040050
Chicago/Turabian StyleCarter, Kimberley C., Léa Fieschi-Méric, Francesca Servini, Mark Wilkinson, David J. Gower, Benjamin Tapley, and Christopher J. Michaels. 2021. "Investigating the Effect of Disturbance on Prey Consumption in Captive Congo Caecilians Herpele squalostoma" Journal of Zoological and Botanical Gardens 2, no. 4: 705-715. https://doi.org/10.3390/jzbg2040050