
Increased COVID-19 Mortality and Deficient SARS-CoV-2 Immune Response Are Not Associated with Higher Levels of Endemic Coronavirus Antibodies
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
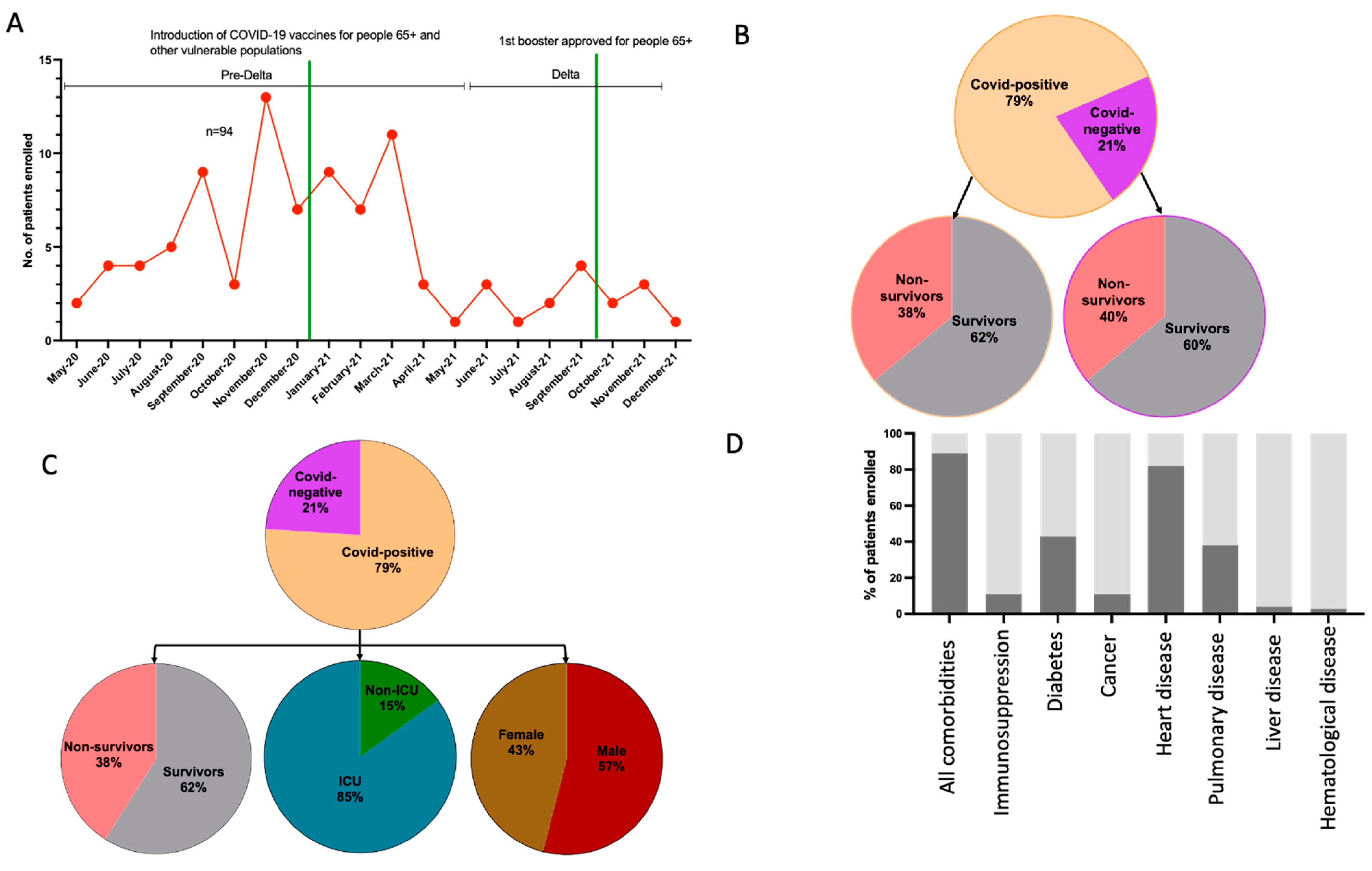
2.1. Study Population
2.2. CCCoV- and SARS-CoV-2 Ab Peptides
2.3. Reference Sera Used for Peptide Characterization Sensitivity and Specificity and ELISA Validation
2.4. ELISA
2.5. Statistical Analysis
3. Results
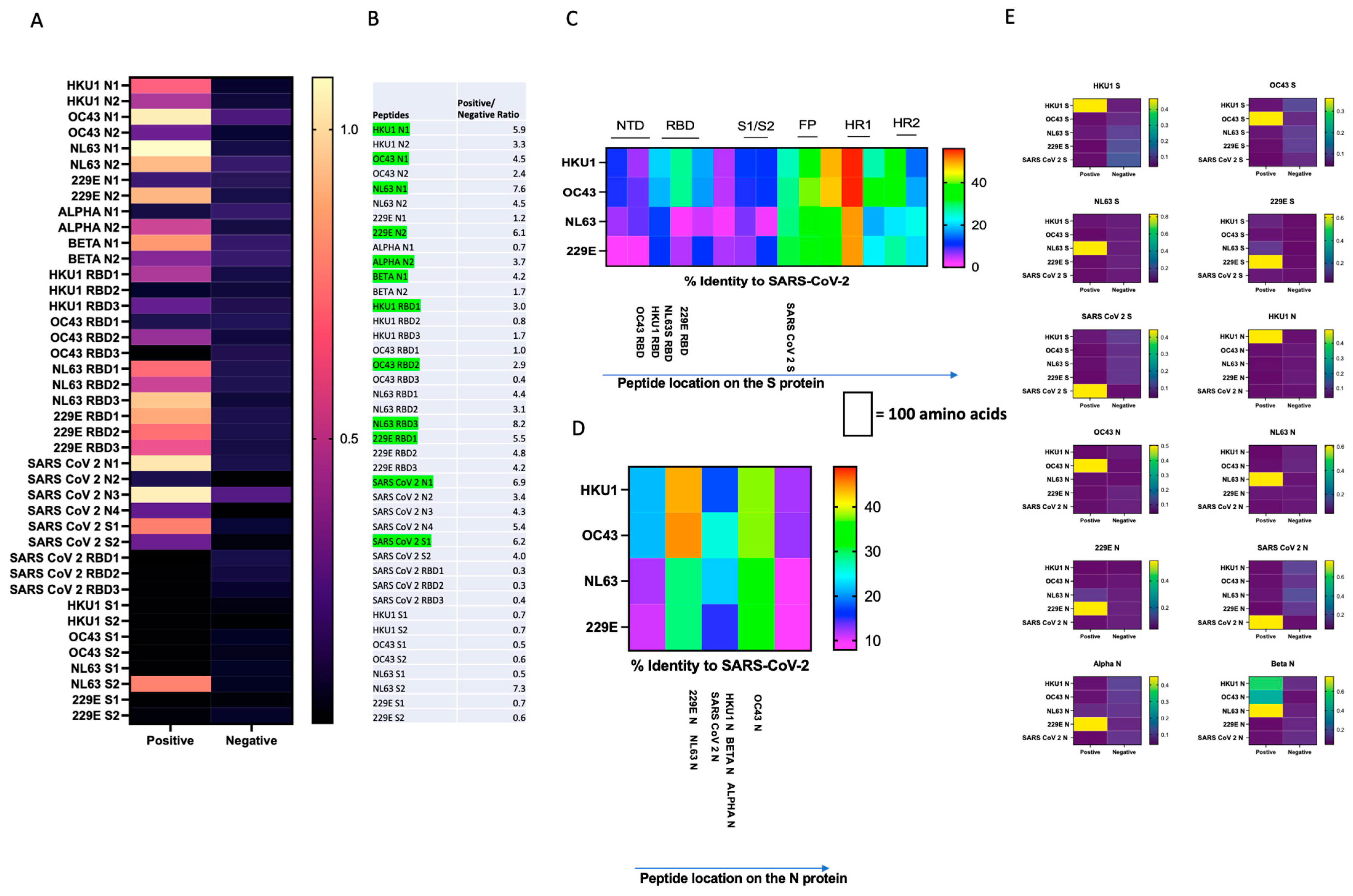
3.1. Design and Characterization of SARS-CoV-2 and CCCoV N and S Protein Peptides and ELISA Development Subsection
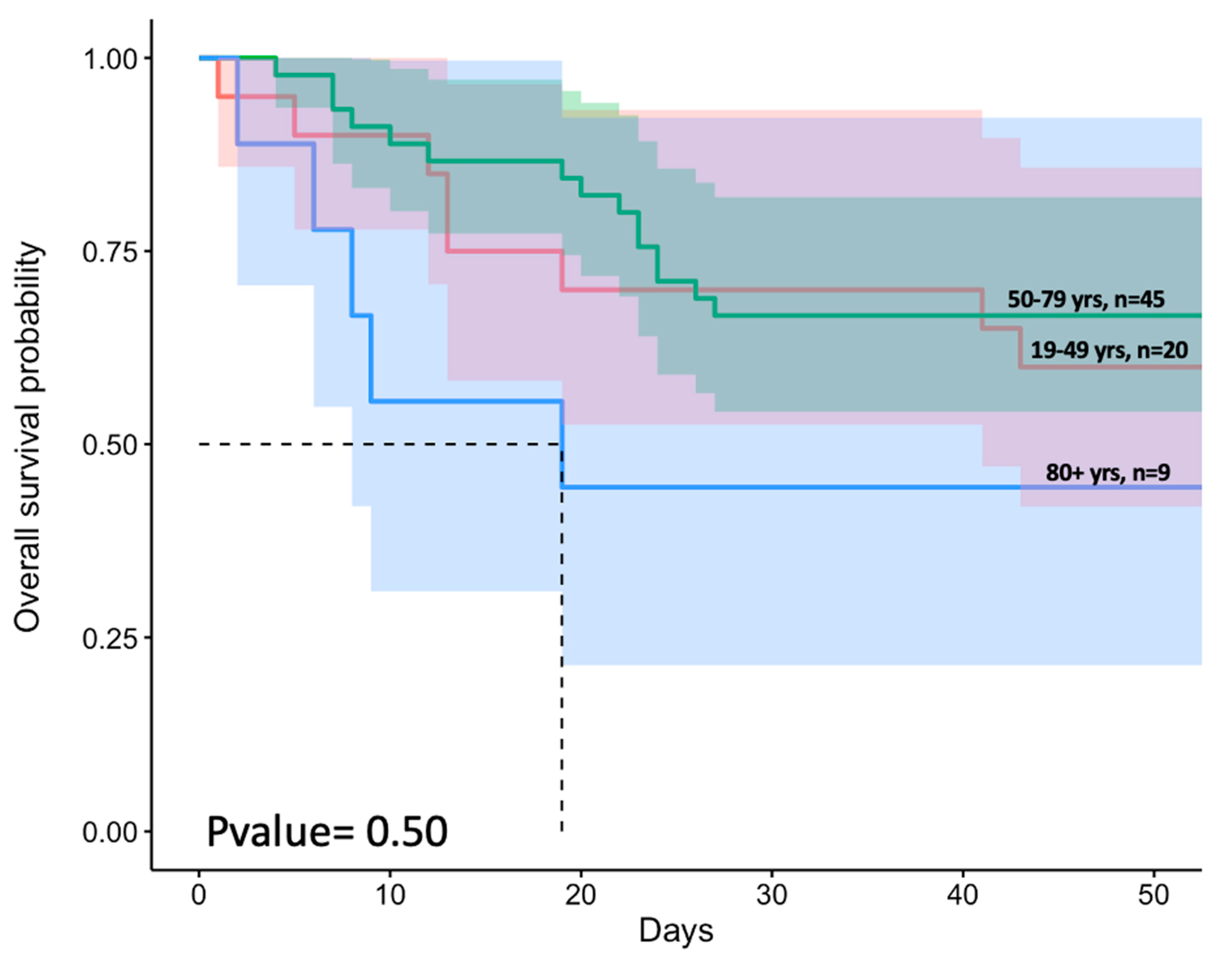
3.2. Advanced Age and Higher Prevalence of Comorbidities Were Associated with Increased Patient Mortality
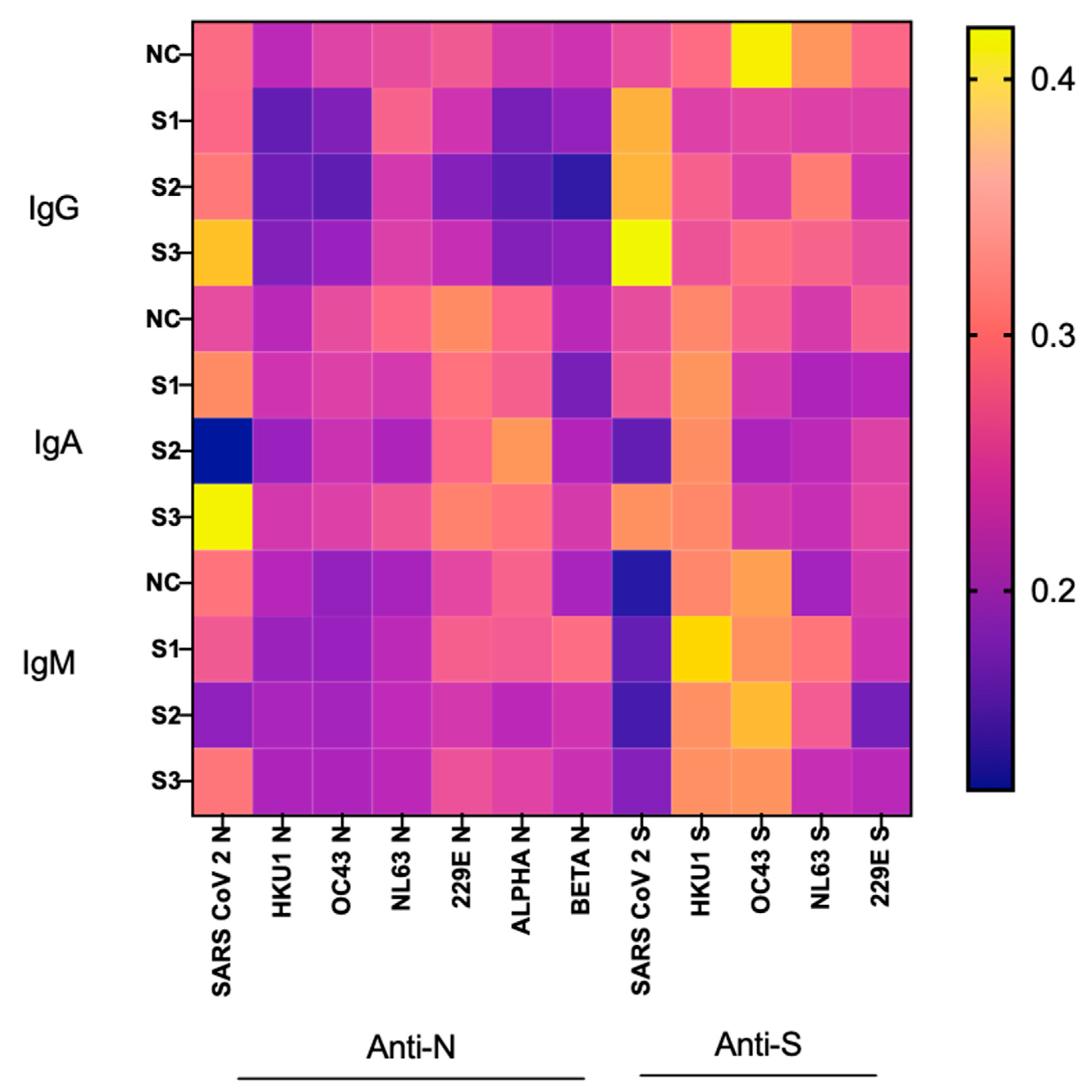
3.3. Correlation between CCCoV- and SARS-CoV-2-Specific Ab Levels and COVID-19 Severity
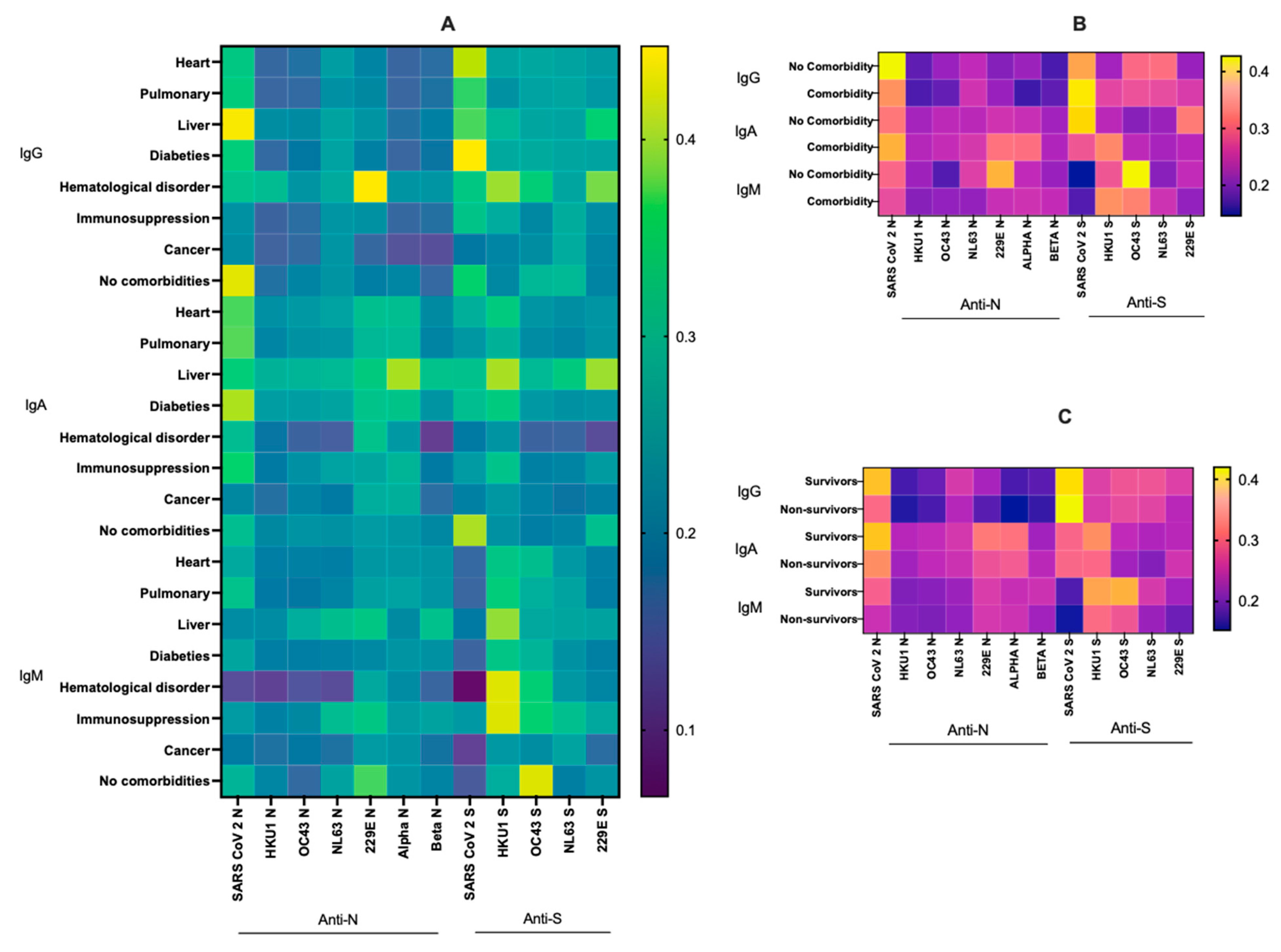
3.4. Relationship between Virus-Specific Ab Levels, Survival and Various Comorbidities
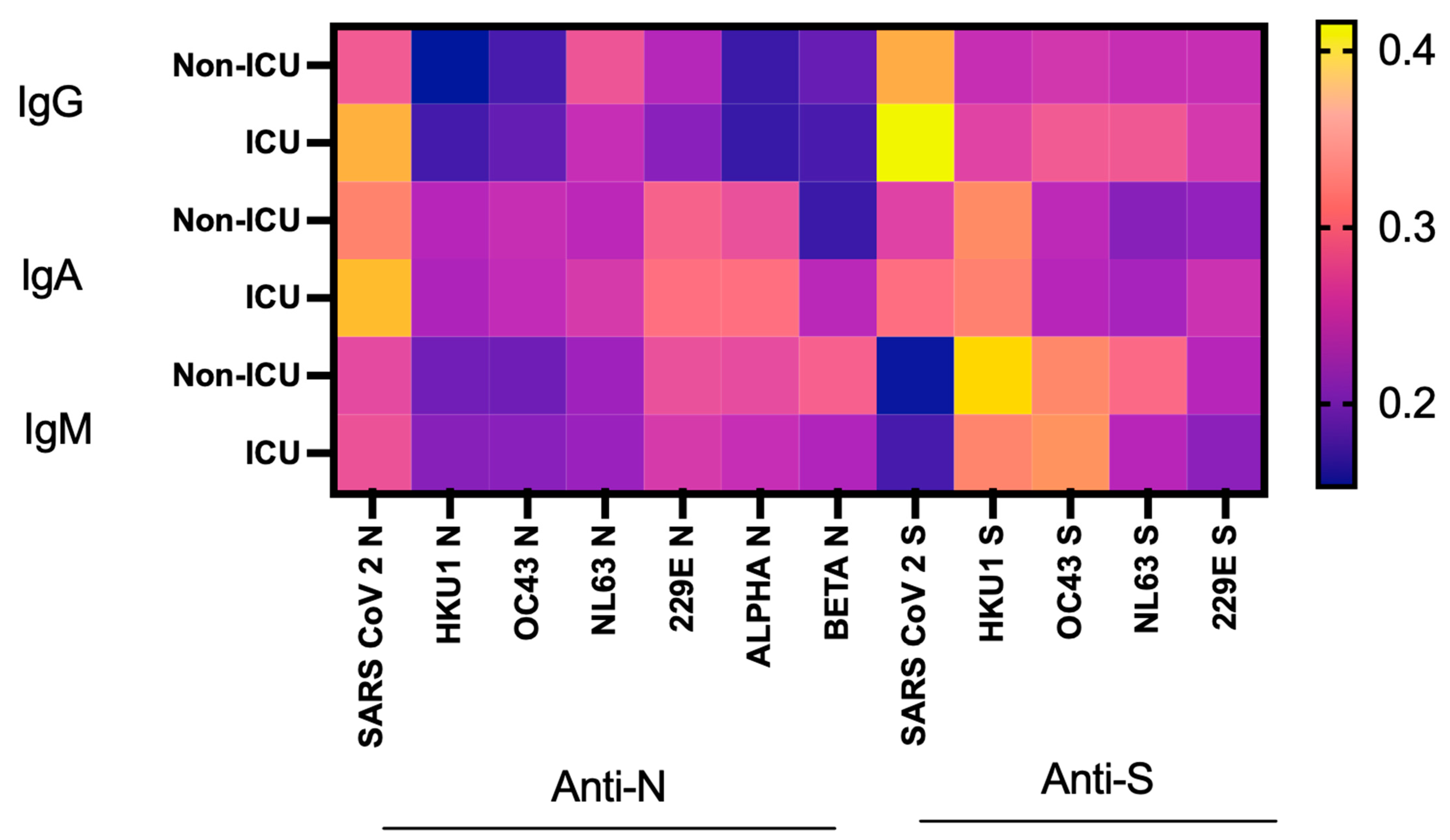
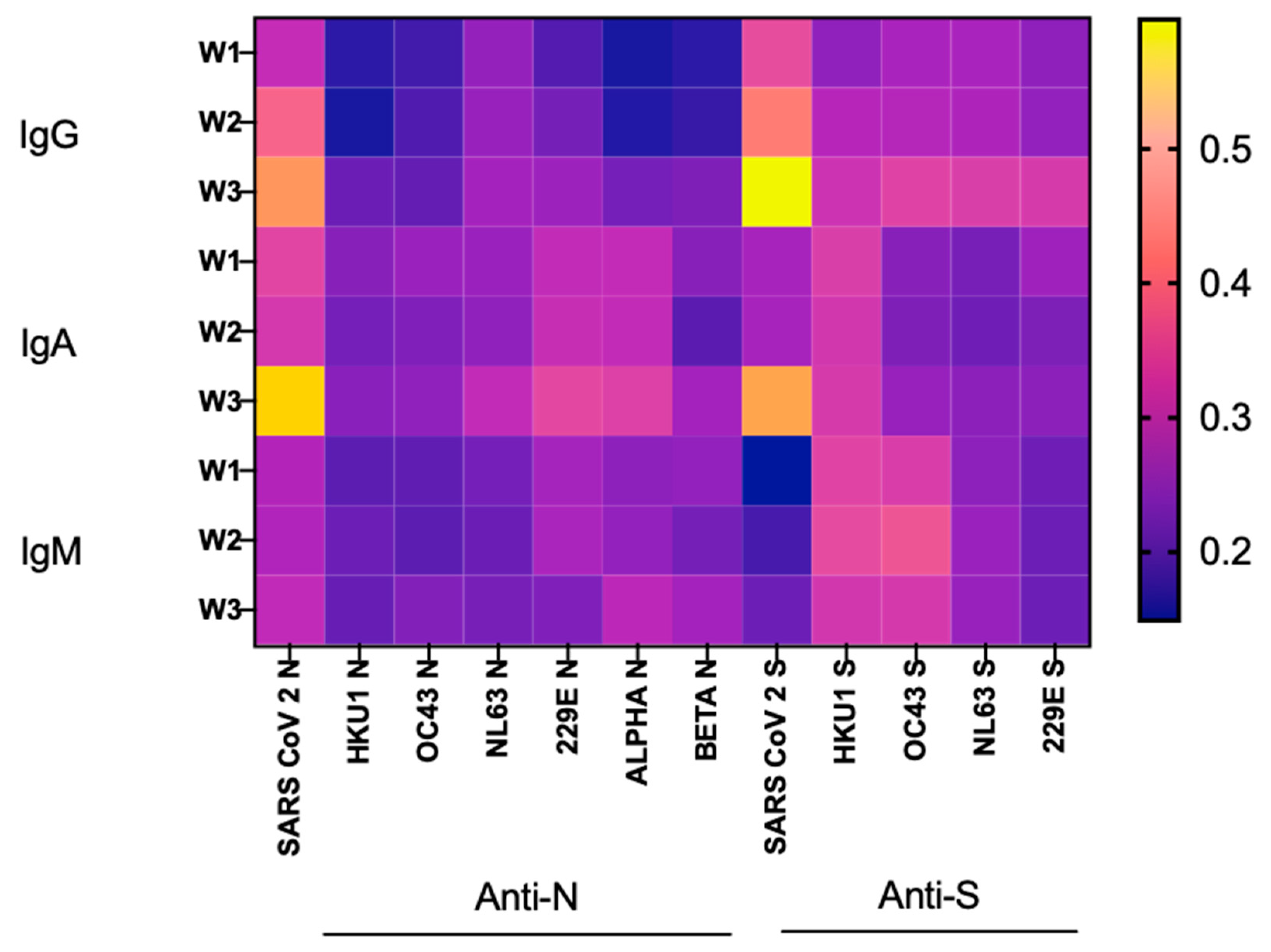
3.5. SARS-CoV-2 Ab Levels, Dynamics and the Risk of ICU Admission
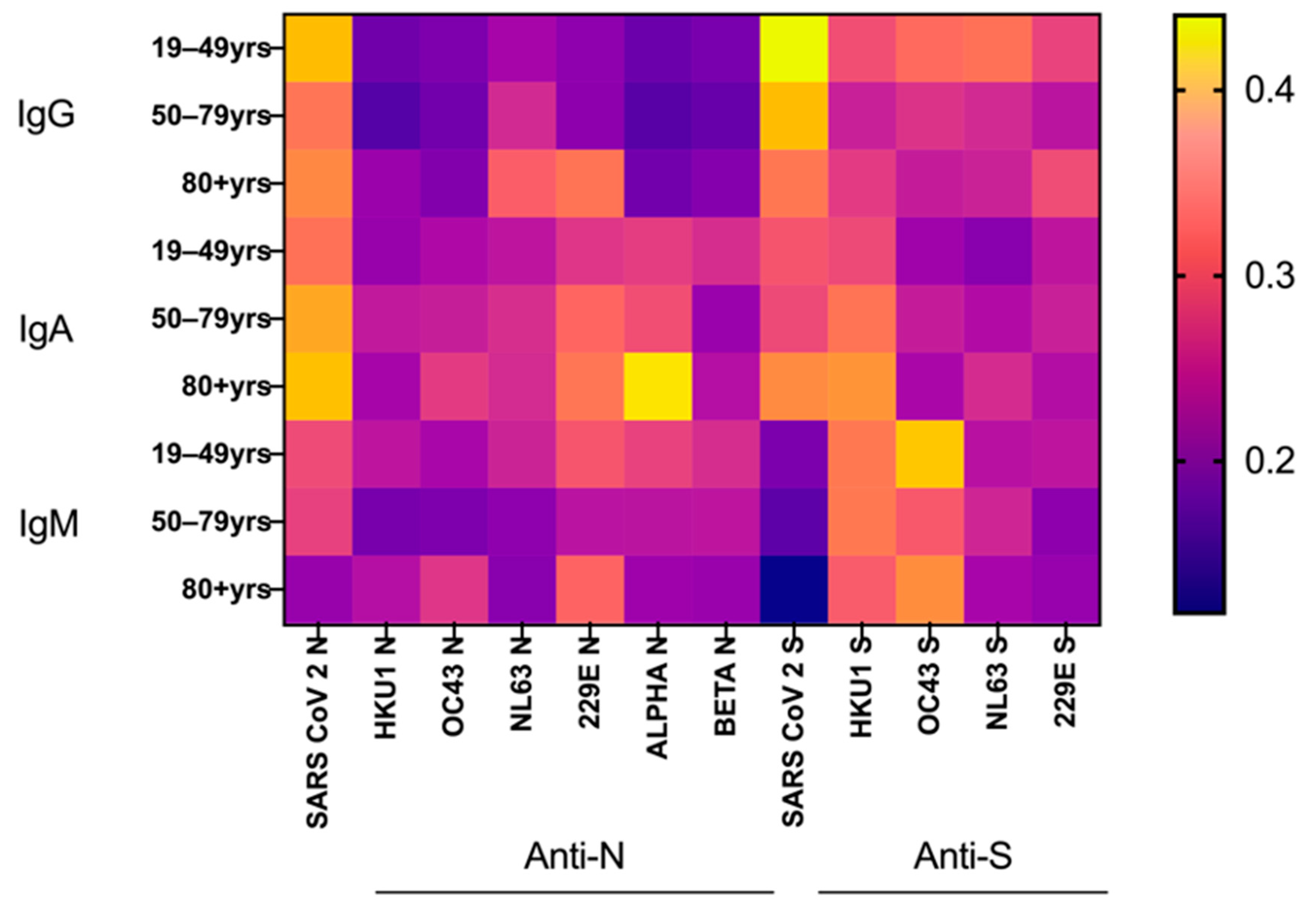
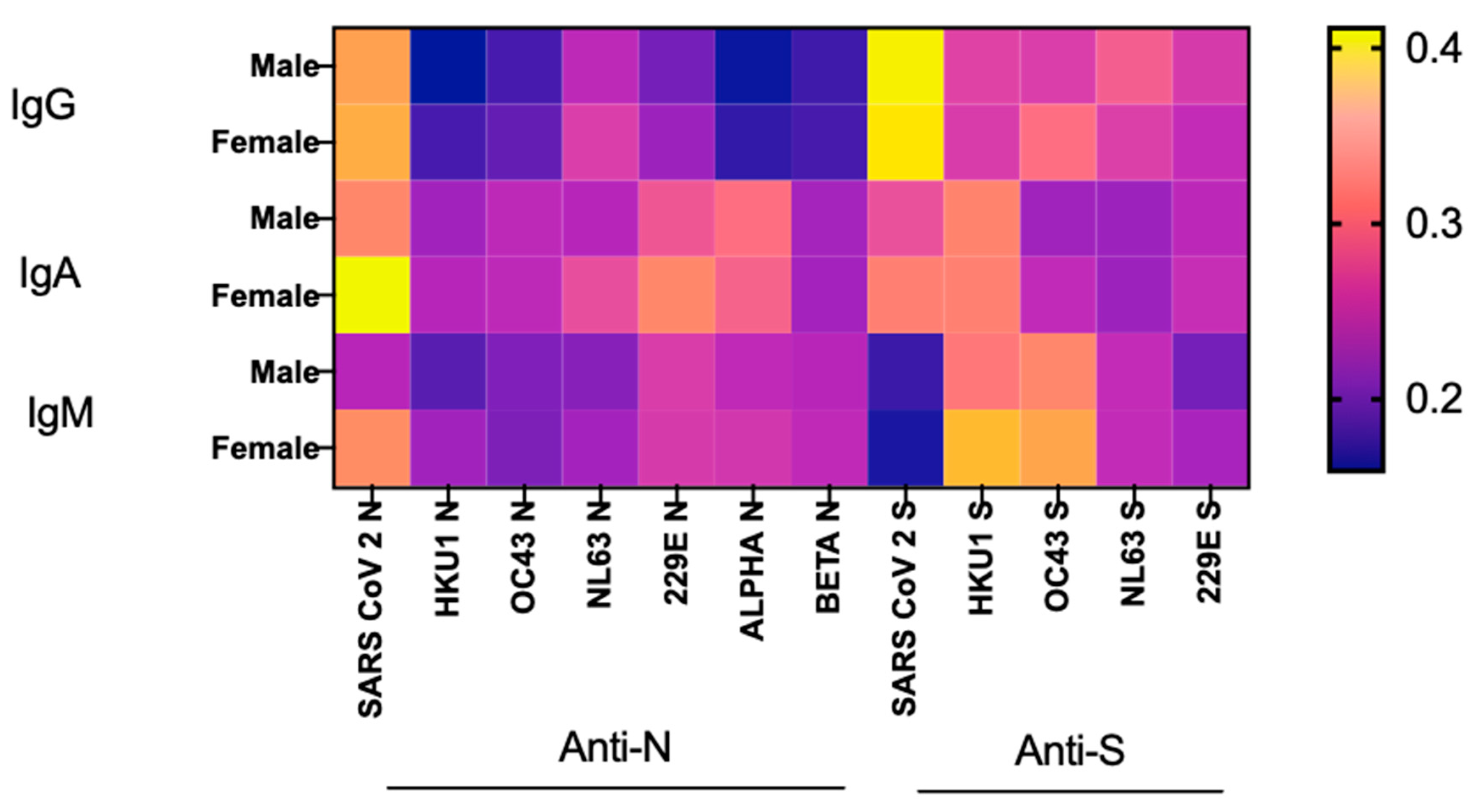
3.6. Age and Sex Effects on SARS-CoV-2 Ab Responses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; Junglen, S.; et al. Changes to virus taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2019). Arch. Virol. 2019, 164, 2417–2429. [Google Scholar] [CrossRef]
- Chan, J.F.; To, K.K.; Tse, H.; Jin, D.Y.; Yuen, K.Y. Interspecies transmission and emergence of novel viruses: Lessons from bats and birds. Trends Microbiol. 2013, 21, 544–555. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Wolf, J.; Brice, D.C.; Sun, Y.; Locke, M.; Cherry, S.; Castellaw, A.H.; Wehenkel, M.; Crawford, J.C.; Zarnitsyna, V.I.; et al. Pre-existing humoral immunity to human common cold coronaviruses negatively impacts the protective SARS-CoV-2 antibody response. Cell Host Microbe 2022, 30, 83–96.e84. [Google Scholar] [CrossRef]
- Miyara, M.; Saichi, M.; Sterlin, D.; Anna, F.; Marot, S.; Mathian, A.; Atif, M.; Quentric, P.; Mohr, A.; Claër, L.; et al. Pre-COVID-19 Immunity to Common Cold Human Coronaviruses Induces a Recall-Type IgG Response to SARS-CoV-2 Antigens Without Cross-Neutralisation. Front. Immunol. 2022, 13, 790334. [Google Scholar] [CrossRef] [PubMed]
- Monroe, I.; Dale, M.; Schwabe, M.; Schenkel, R.; Schenarts, P.J. The COVID-19 Patient in the Surgical Intensive Care Unit. Surg. Clin. North Am. 2022, 102, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, D.; Cameron, A.; Zhou, Q.; Wiltse, A.; Nayak, J.; Pecora, N.D.; Zand, M.S. IgG Against Human Betacoronavirus Spike Proteins Correlates With SARS-CoV-2 Anti-Spike IgG Responses and COVID-19 Disease Severity. J. Infect. Dis. 2022, 226, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Wells, D.A.; Cantoni, D.; Mayora-Neto, M.; Genova, C.D.; Sampson, A.; Ferrari, M.; Carnell, G.; Nadesalingam, A.; Smith, P.; Chan, A.; et al. Human seasonal coronavirus neutralization and COVID-19 severity. J. Med. Virol. 2022, 94, 4820–4829. [Google Scholar] [CrossRef]
- Sagar, M.; Reifler, K.; Rossi, M.; Miller, N.S.; Sinha, P.; White, L.F.; Mizgerd, J.P. Recent endemic coronavirus infection is associated with less-severe COVID-19. J. Clin. Investig. 2021, 131, e143380. [Google Scholar] [CrossRef]
- Wratil, P.R.; Schmacke, N.A.; Karakoc, B.; Dulovic, A.; Junker, D.; Becker, M.; Rothbauer, U.; Osterman, A.; Spaeth, P.M.; Ruhle, A.; et al. Evidence for increased SARS-CoV-2 susceptibility and COVID-19 severity related to pre-existing immunity to seasonal coronaviruses. Cell Rep. 2021, 37, 110169. [Google Scholar] [CrossRef]
- Waterlow, N.R.; Leeuwen, E.V.; Davies, N.G. CMMID COVID-19 Working Group; Flasche, S.; Eggo, R.M. How immunity from and interaction with seasonal coronaviruses can shape SARS-CoV-2 epidemiology. medRxiv 2021. [Google Scholar] [CrossRef]
- Lista, F.; Peragallo, M.S.; Biselli, R.; De Santis, R.; Mariotti, S.; Nisini, R.; D’Amelio, R. Have Diagnostics, Therapies, and Vaccines Made the Difference in the Pandemic Evolution of COVID-19 in Comparison with “Spanish Flu”? Pathogens 2023, 12, 868. [Google Scholar] [PubMed]
- Vlasova, A.N.; Zhang, X.; Hasoksuz, M.; Nagesha, H.S.; Haynes, L.M.; Fang, Y.; Lu, S.; Saif, L.J. Two-way antigenic cross-reactivity between severe acute respiratory syndrome coronavirus (SARS-CoV) and group 1 animal CoVs is mediated through an antigenic site in the N-terminal region of the SARS-CoV nucleoprotein. J. Virol. 2007, 81, 13365–13377. [Google Scholar] [CrossRef] [PubMed]
- Hohdatsu, T.; Yamada, M.; Tominaga, R.; Makino, K.; Kida, K.; Koyama, H. Antibody-dependent enhancement of feline infectious peritonitis virus infection in feline alveolar macrophages and human monocyte cell line U937 by serum of cats experimentally or naturally infected with feline coronavirus. J. Vet. Med. Sci. 1998, 60, 49–55. [Google Scholar] [CrossRef]
- Biswas, M.; Rahaman, S.; Biswas, T.K.; Haque, Z.; Ibrahim, B. Association of Sex, Age, and Comorbidities with Mortality in COVID-19 Patients: A Systematic Review and Meta-Analysis. Intervirology 2021, 64, 36–47. [Google Scholar] [CrossRef]
- Djaharuddin, I.; Munawwarah, S.; Nurulita, A.; Ilyas, M.; Tabri, N.A.; Lihawa, N. Comorbidities and mortality in COVID-19 patients. Gac. Sanit. 2021, 35 (Suppl. 2), S530–S532. [Google Scholar] [CrossRef]
- Henkens, M.; Raafs, A.G.; Verdonschot, J.A.J.; Linschoten, M.; van Smeden, M.; Wang, P.; van der Hooft, B.H.M.; Tieleman, R.; Janssen, M.L.F.; Ter Bekke, R.M.A.; et al. Age is the main determinant of COVID-19 related in-hospital mortality with minimal impact of pre-existing comorbidities, a retrospective cohort study. BMC Geriatr. 2022, 22, 184. [Google Scholar] [CrossRef]
- Mahmoud, M.; Carmisciano, L.; Tagliafico, L.; Muzyka, M.; Rosa, G.; Signori, A.; Bassetti, M.; Nencioni, A.; Monacelli, F. Patterns of Comorbidity and In-Hospital Mortality in Older Patients With COVID-19 Infection. Front. Med. 2021, 8, 726837. [Google Scholar] [CrossRef]
- Arnold, C.G.; Libby, A.; Vest, A.; Hopkinson, A.; Monte, A.A. Immune mechanisms associated with sex-based differences in severe COVID-19 clinical outcomes. Biol. Sex Differ. 2022, 13, 7. [Google Scholar] [CrossRef]
- Kwon, H.; Schafer, J.M.; Song, N.J.; Kaneko, S.; Li, A.; Xiao, T.; Ma, A.; Allen, C.; Das, K.; Zhou, L.; et al. Androgen conspires with the CD8(+) T cell exhaustion program and contributes to sex bias in cancer. Sci. Immunol. 2022, 7, eabq2630. [Google Scholar] [CrossRef]
- Qi, S.; Ngwa, C.; Morales Scheihing, D.A.; Al Mamun, A.; Ahnstedt, H.W.; Finger, C.E.; Colpo, G.D.; Sharmeen, R.; Kim, Y.; Choi, H.A.; et al. Sex differences in the immune response to acute COVID-19 respiratory tract infection. Biol. Sex Differ. 2021, 12, 66. [Google Scholar] [CrossRef] [PubMed]
- Schurz, H.; Salie, M.; Tromp, G.; Hoal, E.G.; Kinnear, C.J.; Möller, M. The X chromosome and sex-specific effects in infectious disease susceptibility. Hum. Genom. 2019, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Hejblum, B.P.; Simon, N.; Jojic, V.; Dekker, C.L.; Thiébaut, R.; Tibshirani, R.J.; Davis, M.M. Systems analysis of sex differences reveals an immunosuppressive role for testosterone in the response to influenza vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 869–874. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coronavirus Species, Peptide Location | Sequence | Antigenicity Score | Peptide Position | Hydrophobicity (%) | Source |
|---|---|---|---|---|---|
| HKU1 N | GSKLELVKRESEADSPVKDV | 21.2 | 324–343 | 35 | Biomatik |
| OC43 N | AEDISLLKKMDEPYTEDTSE | 26 | 428–447 | 30 | Biomatik |
| NL63 N | PRADKPSQLKKPRWKRVPTR | 21.6 | 223–242 | 40 | Biomatik |
| 229E N | SSETKEQKHEMQKPRWKRQP | 22.2 | 234–253 | 20 | Biomatik |
| SARS-CoV-2 N | HIDAYKTFPPTEPKKDKKKK | 21.8 | 356–375 | 30 | Biomatik |
| ALPHA N | VANGVKAKGYPQFAELVPST | NA | 286–322 | 50 | Biomatik |
| BETA N | MLKLGTSDPQFPILAELAPT | NA | 303–322 | 60 | Biomatik |
| HKU1 S | SSRNESWHFDKSEPLCLFKK | 12.4 | 168–187 | 30 | Biomatik |
| OC43 S | LNCPLDPRLKGSFNDRDTGP | 15.8 | 19–38 | 35 | Biomatik |
| NL63 S | IYNRVKSGSPGDSSWHIYLK | 9.4 | 527–546 | 30 | Biomatik |
| 229E S | SWSDGDVITGVPKPVEGVSS | 10 | 415–434 | 40 | Biomatik |
| SARS-CoV-2 S | YDPLQPELDSFKEELDKYFK | 19.6 | 1120–1139 | 35 | Biomatik |
| SARS-CoV-2 seronegative serum samples | Negative serum samples (n = 7) from healthy individuals prior to 2019 (provided by Shan-Lu Liu)—SARS-CoV-2 N and S protein ELISA development and validation. Commercial pre-pandemic normal human serum (SARS-CoV-2 Neutralizing Antibody-Negative Pre-pandemic Human Serum, Cayman chemicals, Item No. 31569)—ELISA validation and as a negative control sample to determine cutoff values for each SARS-CoV-2 Ab test. Seventy-eight SARS-CoV-2 seronegative samples—SeroNet Blinded Panel. |
| SARS-CoV-2 seropositive serum samples | Thirty samples from 10 COVID-19 patients from BUCK-ICU biorepository (3 longitudinal samples per patient) with variable disease severity—SARS-CoV-2 N and S protein ELISA development and validation. Thirty-one SARS-CoV-2 seropositive samples—SeroNet Blinded Panel. |
| Virus/antigen-specific and negative rabbit serum samples | Commercial CCCoV-specific and negative rabbit antisera (Table S1) were used for CCCoV N and S protein ELISA development and validation. |
| Test | SARS-CoV-2 (N) | SARS-CoV-2 (S) |
|---|---|---|
| Sensitivity | 95.08% | 96.77% |
| Specificity | 97.65% | 100% |
| Accuracy | 96.58% | 98.63% |
| Clinical and Demographic Variables | Patient Median Age | p Value |
|---|---|---|
| Survivors (n = 46) | 61 | 0.32 |
| Non-survivors (n = 28) | 62 | |
| Male (n = 42) | 61 | 0.44 |
| Female (n = 32) | 63 | |
| No comorbidity (n = 8) | 36 | 0.001 |
| Comorbidity (n = 66) | 63 |
| Comorbidity | Patients | |
|---|---|---|
| Non-survivors, % | Survivors, % | |
| Heart disease (n = 76) | 39 | 61 |
| Pulmonary disease (n = 37) | 41 | 59 |
| Liver disease (n = 5) | 80 | 20 |
| Diabetes (n = 39) | 41 | 59 |
| Hematological disorder (n = 3) | 0 | 100 |
| Immunosuppression (n = 12) | 42 | 58 |
| Cancer (n = 11) | 55 | 45 |
| No comorbidities (n = 8) | 25 | 75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, B.; Oltz, E.M.; Bednash, J.S.; Horowitz, J.C.; Amimo, J.O.; Raev, S.A.; Fernández, S.; Anghelina, M.; Liu, S.-L.; Rubinstein, M.P.; et al. Increased COVID-19 Mortality and Deficient SARS-CoV-2 Immune Response Are Not Associated with Higher Levels of Endemic Coronavirus Antibodies. Immuno 2023, 3, 330-345. https://doi.org/10.3390/immuno3030020
Adhikari B, Oltz EM, Bednash JS, Horowitz JC, Amimo JO, Raev SA, Fernández S, Anghelina M, Liu S-L, Rubinstein MP, et al. Increased COVID-19 Mortality and Deficient SARS-CoV-2 Immune Response Are Not Associated with Higher Levels of Endemic Coronavirus Antibodies. Immuno. 2023; 3(3):330-345. https://doi.org/10.3390/immuno3030020
Chicago/Turabian StyleAdhikari, Bindu, Eugene M. Oltz, Joseph S. Bednash, Jeffrey C. Horowitz, Joshua O. Amimo, Sergei A. Raev, Soledad Fernández, Mirela Anghelina, Shan-Lu Liu, Mark P. Rubinstein, and et al. 2023. "Increased COVID-19 Mortality and Deficient SARS-CoV-2 Immune Response Are Not Associated with Higher Levels of Endemic Coronavirus Antibodies" Immuno 3, no. 3: 330-345. https://doi.org/10.3390/immuno3030020